Isopoda
Les Isopodes (Isopoda) forment un ordre extrêmement varié parmi les Crustacés, et ne comptent pas moins de 10 000 espèces. L'ordre des Isopodes a été déterminé par Pierre-André Latreille en 1817.

| Règne | Animalia |
|---|---|
| Embranchement | Arthropoda |
| Sous-embr. | Crustacea |
| Classe | Malacostraca |
| Sous-classe | Eumalacostraca |
| Super-ordre | Peracarida |
La taille des Isopodes varie de 0,5 mm à 50 cm (Bathynome géant). Si les espèces les plus familières, comme les cloportes, ont le corps aplati dorsoventralement, leur aspect extérieur et les détails de leur morphologie peuvent varier considérablement en fonction des adaptations à des conditions de vie très variées. Les espèces parasites, nombreuses, sont celles qui présentent les particularités les plus remarquables.
La plupart des Isopodes sont marins, mais il existe des espèces d'eau douce (notamment les Aselles), et aussi un groupe important d’espèces terrestres (les Cloportes, Oniscoidea (en)). Les Isopodes sont même les seuls crustacés à avoir réussi à se libérer du milieu aquatique tout au long de leur cycle vital.
Les Isopodes sont herbivores, détritivores, carnivores ou parasites. Certaines espèces, notamment du genre Limnoria, rongeuses de bois (xylophages) en milieu marin, sont susceptibles d'avoir un impact économique important.
Anatomie

Les isopodes constituent un groupe extrêmement diversifié et la variété des morphologies possibles ne permet d’envisager ici que le cas des formes libres dans leurs aspects les plus caractéristiques[2],[3].
Comme chez tous les eumalacostracés, le corps des isopodes est constitué de 19 somites (segments = métamères) : 5 pour le céphalon, 8 pour le péréion et 6 pour le pléon.
Au moins un métamère du péréion (parfois 2) se soude au céphalon pour constituer la tête ; il ne reste donc plus que 7 segments libres. D’autre part, à l’arrière du corps, le dernier somite du pléon est souvent soudé au telson, ce qui ne laisse que 5 somites libres. Le nombre des segments plus ou moins complètement fusionnés entre eux et au telson peut être plus important, l’ensemble constituant un pléotelson.
Tête
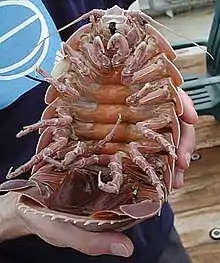
Elle porte les yeux, pairs, fixes et 6 paires d’appendices :
- Les antennules (A1), en général courtes, voire presque vestigiales dans les espèces terrestres, et uniramées.
- Les antennes (A2) de longueur très variable. Elles comportent un pédoncule de 5-6 articles. Chez les aselles, une écaille (= squama) sur le troisième d’entre eux représente un exopodite.
- Les mandibules (Md) possèdent une partie tranchante (processus incisif), une région broyeuse (processus molaire) et entre les deux, au moins du côté gauche, une lame mobile (lacinia mobilis). Elle est parfois munie d’un palpe de 3 articles.
- Les maxillules (Mx1) et
- les maxilles (Mx2) sont deux paires d’appendices de structure lamelleuse, appliquées en arrière des mandibules.
- Les maxillipèdes (Mxp), qui sont les appendices du premier métamère du péréion soudé à la tête. Incorporés aux pièces buccales ils n’ont plus de fonctions locomotrices et diffèrent nettement des pattes suivantes. Leur a coxa est munie d’une lamelle (épipodite) qui recouvre plus ou moins les parties latérales de pièces antérieures. D’autre part, chez les femelles incubantes, la base des Mxp peut porter une expansion lamelleuse, semblable à un oostégite, dont les battements amélioreraient la circulation de l’eau dans la cavité incubatrice et l’oxygénation des embryons.
Chez les espèces parasites les pièces buccales sont regroupées en un cône pointu permettant de percer le tégument de l’hôte et d’aspirer son liquide circulant (« sang »).
Il n’y a pas de carapace, au sens zoologique du terme, chez les isopodes.
Péréion
Il comporte 7 somites porteurs d’une paire d’appendices, les péréiopodes, ou pattes, dédiés généralement à la marche mais susceptibles de présenter des adaptations diverses (notamment à la préhension). Leur relative similitude en taille et forme justifie l’appellation d’isopodes (iso = semblable, égal). La validité de cette dénomination est cependant contestée [3].
Les pattes sont constituées d’une base de deux articles, la coxa puis le basis, sur lequel s’articule le reste de l’appendice composé de 5 articles, l’ischion (généralement long), le merus, le carpe, le propode et le dactyle (appelés également ischiopodite, méropodite etc.) qui représente la rame interne (endopodite) de l’appendice primitif. Il n’y a pas de rame externe (exopodite). Le dactyle peut être porteur de griffes.
Il est fréquent que la coxa s’étale en plaque coxale et se soude avec le corps de telle sorte qu’elle devient visible sur la face dorsale de l’animal en remplaçant totalement ou en partie l’épimère (jonction entre la plaque dorsale et ventrale du segment).
Chez les femelles en période de reproduction, des plaques lamelleuses, ou oostégites, se développent sur la face ventrale des segments thoraciques libres, en principe à partir de la coxa. Il peut en exister 7 paires, plus généralement 5 et parfois une seule (segment 4) chez les arcturidés. Les oostégites délimitent avec la paroi ventrale du corps une poche incubatrice ou marsupium dans laquelle se développent les œufs.
Les orifices génitaux femelles sont situés à la base des péréiopodes 5 et les orifices mâles à l’extrémité des deux papilles génitales (« pénis », fusionnés en un seul chez les cloportes) situés au milieu de la face ventrale du dernier (7e) métamère thoracique. La mue des isopodes s’accomplit en deux temps : approximativement la moitié postérieure d’abord, puis la moitié antérieure. On trouve donc sur le terrain des animaux ayant l’arrière du thorax nettement plus large que l’avant, cette apparente anomalie dénote simplement le fait que l’animal n’a encore accompli que la moitié de son exuviation.
Pléon
Il est constitué de 6 segments dont le dernier est soudé au telson. Éventuellement d’autres segments peuvent fusionner plus ou moins complètement avec cet ensemble pour constituer un pléotelson. Tous les segments sont fusionnés chez les aselles d’eau douce. En principe chaque segment porte une paire d’appendices typiquement biramés, les pléopodes. La dernière paire cependant, d’une forme particulière, est appelée uropodes. Une partie au moins des pléopodes possède une fonction respiratoire, ce sont les branchies des espèces aquatiques. Les espèces terrestres possèdent des cavités plus ou moins complexes (pseudotrachées par exemple) permettant la respiration aérienne. Cette fonction respiratoire des pléopodes constitue le caractère unitaire le plus important de l’ordre [3].
Les pléopodes ont également une fonction natatoire chez les espèces aquatiques. Le partage entre les deux fonctions ci-dessus se réalise de manière variable selon les espèces, soit latéralement (endopodite plutôt respiratoire, exopodite plutôt natatoire) soit antéropostérieurement (les antérieurs natatoires, les postérieurs respiratoires). Certains pléopodes peuvent aussi se différencier en plaques operculaires protégeant les branchies sous-jacentes.
Chez les mâles l’endopodite du 2e pléopode porte une excroissance en forme de bâtonnet creusé en gouttière, l’appendix masculina. Le premier pléopode est parfois modifié également. Les uropodes sont en général biramés et, sauf quelques exceptions, les rames sont constituées d’un seul article. Ils prennent l’aspect de plaques operculaires recouvrant, comme deux battants d’un portail, les pléopodes chez les valvifères (g. Idotea par exemple).
Reproduction
Les sexes sont séparés mais il y a quelques cas d’hermaphrodisme protandrique (cymothoidés) ou protogynique (anthuridés : g. Cyathura). Le dimorphisme sexuel est généralement discret mais parfois très spectaculaire dans des espèces libres marines comme "Dynamene bidentata" ou Campecopea hirsuta ou chez les parasites comme les bopyres (par exemple Bopyrus fougerouxi, parasite de la crevette rose Palaemon serratus) ou les gnathidés (exemple Paragnathia formica, commun dans les zones estuariennes). Dynamene bidentata occupe souvent la cavité formée par la muraille de balanes mortes. La partie postérieure du corps du mâle, pourvue de deux épines obture l’entrée et le fond de la cavité est occupé par un " harem " de plusieurs femelles.
Lors de l’accouplement le mâle chevauche la femelle et recourbe l’arrière de son corps pour le glisser sous le thorax de la femelle, déposant son sperme à l’un des orifices génitaux puis il accomplit la même manœuvre pour inséminer l’autre côté. Les appendix masculina servent probablement à conduire le sperme depuis les « pénis » jusqu’aux orifices femelles.
Les œufs se développent dans la cavité incubatrice qui libère des jeunes pratiquement identiques aux adultes (développement direct), hormis qu’il leur manque la dernière paire de pattes thoraciques.
Chez certains sphéromes l’incubation des œufs se déroule dans des poches formées par une invagination du tégument thoracique ventral à l’intérieur du corps de la femelle, les oostégites sont présents ou non. Les femelles de gnathidés incubent également leurs œufs à l’intérieur de leur corps qui en est extrêmement dilaté.
La sexualité de nombreux isopodes, surtout terrestres, peut être fortement perturbée par l’action de la protéobactérie intracellulaire Wolbachia dont le principal effet est de transformer les mâles en femelles fonctionnelles[4].
Classification
Compte tenu de la variété des espèces contenues dans l’ordre des isopodes leur classification n’est pas encore définie de manière définitive. Elle comporte la subdivision en une douzaine de sous-ordres comprenant près de 100 familles. Sous-ordres et familles :
Selon World Register of Marine Species (20 janvier 2018)[5] (liste incomplète pour les taxons terrestres) :
- sous-ordre Asellota Latreille, 1802
- super-famille Aselloidea Latreille, 1802
- super-famille Gnathostenetroidoidea Kussakin, 1967
- famille Gnathostenetroididae Kussakin, 1967
- famille Protojaniridae Fresi, Idato & Scipione, 1980
- super-famille Janiroidea G.O. Sars, 1897
- famille Acanthaspidiidae Menzies, 1962
- famille Dendrotionidae Vanhöffen, 1914
- famille Desmosomatidae G.O. Sars, 1897
- famille Echinothambematidae Menzies, 1956
- famille Haplomunnidae Wilson, 1976
- famille Haploniscidae Hansen, 1916
- famille Ischnomesidae Hansen, 1916
- famille Janirellidae Menzies, 1956
- famille Janiridae G.O. Sars, 1897
- famille Joeropsididae Nordenstam, 1933
- famille Katianiridae Svavarsson, 1987
- famille Lepidocharontidae Galassi & Bruce, 2016
- famille Macrostylidae Hansen, 1916
- famille Mesosignidae Schultz, 1969
- famille Microparasellidae Karaman, 1933
- famille Mictosomatidae Wolff, 1965
- famille Munnidae G.O. Sars, 1897
- famille Munnopsidae Lilljeborg, 1864
- famille Nannoniscidae Hansen, 1916
- famille Paramunnidae Vanhöffen, 1914
- famille Pleurocopidae Fresi & Schiecke, 1972
- famille Santiidae Kussakin, 1988
- famille Thambematidae Stebbing, 1912
- famille Urstylidae Riehl, Wilson & Malyutina, 2014
- famille Xenosellidae Just, 2005
- super-famille Stenetrioidea Hansen, 1905
- famille Pseudojaniridae Wilson, 1986
- famille Stenetriidae Hansen, 1905
- sous-ordre Calabozoidea Van Lieshout, 1983
- famille Brasileirinidae Pervorčnik, Ferreira & Sket, 2012
- famille Calabozoidae Van Lieshout, 1983
- sous-ordre Cymothoida Wägele, 1989
- super-famille Anthuroidea Leach, 1914
- famille Antheluridae Poore & Lew Ton, 1988
- famille Anthuridae Leach, 1814
- famille Expanathuridae Poore, 2001
- famille Hyssuridae Wägele, 1981
- famille Leptanthuridae Poore, 2001
- famille Paranthuridae Menzies & Glynn, 1968
- super-famille Cymothooidea Leach, 1814
- famille Aegidae White, 1850
- famille Anuropidae Stebbing, 1893
- famille Barybrotidae Hansen, 1890
- famille Cirolanidae Dana, 1852
- famille Corallanidae Hansen, 1890
- famille Cymothoidae Leach, 1818
- famille Gnathiidae Leach, 1814
- famille Protognathiidae Wägele & Brandt, 1988
- famille Tridentellidae Bruce, 1984
- infra-ordre Epicaridea Latreille, 1825
- super-famille Bopyroidea Rafinesque, 1815
- famille Bopyridae Rafinesque, 1815
- famille Colypuridae Richardson, 1905
- famille Entoniscidae Kossmann, 1881
- famille Ionidae H. Milne Edwards, 1840
- super-famille Cryptoniscoidea Kossmann, 1880
- famille Asconiscidae Bonnier, 1900
- famille Cabiropidae Giard & Bonnier, 1887
- famille Crinoniscidae Bonnier, 1900
- famille Cryptoniscidae Kossmann, 1880
- famille Cyproniscidae Giard & Bonnier, 1887
- famille Dajidae Giard & Bonnier, 1887
- famille Entophilidae Richardson, 1903
- famille Hemioniscidae Bonnier, 1900
- famille Podasconidae Giard & Bonnier, 1895
- super-famille Bopyroidea Rafinesque, 1815
- super-famille Anthuroidea Leach, 1914
- sous-ordre Limnoriidea Brandt & Poore in Poore, 2002
- super-famille Limnorioidea White, 1850
- famille Hadromastacidae Bruce & Mueller, 1991
- famille Keuphyliidae Bruce, 1980
- famille Limnoriidae White, 1850
- super-famille Limnorioidea White, 1850
- sous-ordre Microcerberidea Lang, 1961
- famille Atlantasellidae Sket, 1979
- famille Microcerberidae Karaman, 1933
- sous-ordre Oniscidea Latreille, 1802
- infra-ordre Ligiamorpha Vandel, 1943
- section Crinocheta Legrand, 1946
- super-famille Armadilloidea Brandt, 1831
- super-famille Oniscoidea Latreille, 1802
- section Diplocheta Vandel, 1957
- super-famille Trichoniscoidea
- section Crinocheta Legrand, 1946
- famille Agnaridae Schmidt, 2003
- famille Alloniscidae Schmidt, 2003
- famille Armadillidae Brandt, 1831
- famille Armadillidiidae Brandt, 1833
- famille Balloniscidae Vandel, 1963
- famille Bathytropidae Vandel, 1952
- famille Berytoniscidae Vandel, 1955
- famille Bisilvestriidae Verhoeff, 1938
- famille Cylisticidae Verhoeff, 1949
- famille Delatorreiidae Verhoeff, 1938
- famille Detonidae Budde-Lund, 1904
- famille Dubioniscidae Schultz, 1995
- famille Eubelidae Budde-Lund, 1899
- famille Halophilosciidae Verhoeff, 1908
- famille Hekelidae Ferrara, 1977
- famille Irmaosidae Ferrara & Taiti, 1983
- famille Ligiidae Leach, 1814
- famille Mesoniscidae Verhoeff, 1908
- famille Olibrinidae Budde-Lund, 1913
- famille Oniscidae Latreille, 1802
- famille Paraplatyarthridae Javidkar & King, 2015
- famille Philosciidae Kinahan, 1857
- famille Platyarthridae Verhoeff, 1949
- famille Porcellionidae Brandt, 1831
- famille Pudeoniscidae Lemos de Castro, 1973
- famille Rhyscotidae Budde-Lund, 1904
- famille Schoebliidae Verhoeff, 1938
- famille Scleropactidae Verhoeff, 1938
- famille Scyphacidae Dana, 1852
- famille Spelaeoniscidae Vandel, 1948
- famille Stenoniscidae Budde-Lund, 1904
- famille Styloniscidae Vandel, 1952
- famille Tendosphaeridae Verhoeff, 1930
- famille Titanidae Verhoeff, 1938
- famille Trachelipodidae Strouhal, 1953
- famille Trichoniscidae Sars, 1899
- famille Turanoniscidae Borutzky, 1969
- famille Tylidae Dana, 1852
- infra-ordre Ligiamorpha Vandel, 1943
- sous-ordre Phoratopidea Brandt & Poore, 2003
- famille Phoratopodidae Hale, 1925
- sous-ordre Phreatoicidea Stebbing, 1893
- famille Amphisopidae Nicholls, 1943
- famille Hypsimetopidae Nicholls, 1943
- famille Mesamphisopidae Nicholls, 1943
- famille Phreatoicidae Chilton, 1891
- famille Phreatoicopsidae Nicholls, 1943
- famille Ponderellidae Wilson & Keable, 2004
- sous-ordre Sphaeromatidea Wägele, 1989
- super-famille Seroloidea Dana, 1852
- famille Basserolidae Brandt & Poore, 2003
- famille Bathynataliidae Kensley, 1978
- famille Plakarthriidae Hansen, 1905
- famille Serolidae Dana, 1852
- super-famille Sphaeromatoidea Latreille, 1825
- famille Ancinidae Dana, 1852
- famille Sphaeromatidae Latreille, 1825
- famille Tecticipitidae Iverson, 1982
- famille Archaeoniscidae Haack, 1918
- super-famille Seroloidea Dana, 1852
- sous-ordre Tainisopidea Brandt & Poore, 2003
- famille Tainisopidae Wilson, 2003
- sous-ordre Valvifera G. O. Sars, 1883
- famille Antarcturidae Poore, 2001
- famille Arcturidae Dana, 1849
- famille Arcturididae Poore, 2001
- famille Austrarcturellidae Poore & Bardsley, 1992
- famille Chaetiliidae Dana, 1849
- famille Holidoteidae Wägele, 1989
- famille Holognathidae Thomson, 1904
- famille Idoteidae Samouelle, 1819
- famille Pseudidoteidae Ohlin, 1901
- famille Rectarcturidae Poore, 2001
- famille Thermoarcturidae Poore, 2015
- famille Xenarcturidae Sheppard, 1957
 Oniscus asellus (Oniscidae)
Oniscus asellus (Oniscidae)
 Cyathura carinata (Anthuridae)
Cyathura carinata (Anthuridae) Eurydice pulchra (Cirolanidae)
Eurydice pulchra (Cirolanidae).jpg.webp) Paragnathia formica (Gnathiidae)
Paragnathia formica (Gnathiidae) Idotea pelagica (Idoteidae)
Idotea pelagica (Idoteidae) Jaera albifrons (Janiridae)
Jaera albifrons (Janiridae)
Espèce modèle et bioindication
Certains isopodes, en complément de modèles végétaux[6] et d'autres espèces telles que les vers de terre, collemboles et carabes sont proposés comme modèle biologique pour l'étude des sols pollués et/ou comme bioindicateurs pertinents pour l'évaluation environnementale d'un milieu naturel ou sol, semi-naturel, pollué, etc. en particulier dans les contextes de gestion des risques des sites et sols pollués[7].
Dans la culture
Les isopodes sont présents dans la franchise de jeux Pokémon à travers deux créatures : Wimpod et Golisopod, en français Sovkipou et Sarmuraï[8].
Notes et références
- Integrated Taxonomic Information System (ITIS), www.itis.gov, CC0 https://doi.org/10.5066/F7KH0KBK, consulté le 26 mars 2015
- Calman, W.T., 1909. Crustacea. In, Sir R. Lankester ed. A treatise on zoology. A & C. Black, London.346 p.
- Schram, F. R.,1986. Crustacea. Oxford University Press, 606p.
- Didier Bouchon, Thierry Rigaud et Pierre Juchault, « Evidence for widespread Wolbachia infection in isopod crustaceans: molecular identification and host feminization », Proc. R. Soc. Lond., vol. 265, (lire en ligne)
- World Register of Marine Species, consulté le 20 janvier 2018
- Thèse de Géraldine Bidar (2007), portant sur les modèles herbacés ray grass et trèfle, et modèles arborés (Robinia pseudoacacia, Alnus glutinosa et Acer pseudoplatanus
- Jean-Philippe Godet, Intérêt des isopodes terrestres dans l'évaluation de la qualité des sols : Recherche de paramètres indicateurs et mise au point d'un outil écotoxicologique de terrain permettant le diagnostic et le suivi des milieux terrestres pollués par les métaux lourds ; thèse en écologie
- « Sarmuraï », sur pokepedia.fr.
Voir aussi
Références taxinomiques
- (en) Référence World Register of Marine Species : taxon Isopoda Latreille, 1817 (+ liste familles + liste genres)
- (en) Référence Paleobiology Database : Isopoda Latreille 1817
- (fr+en) Référence ITIS : Isopoda Latreille, 1817
- (en) Référence Arthropoda Species Files : Isopoda
- (en) Référence Tree of Life Web Project : Isopoda
- (en) Référence Animal Diversity Web : Isopoda
- (en) Référence uBio : Isopoda Thorell 1881
- (en) Référence Catalogue of Life : Isopoda (consulté le )
- (en) Référence Fauna Europaea : Isopoda
- (en) Référence NCBI : Isopoda (taxons inclus)
Bibliographie
- Pruvot C., Godet J.P., Waterlot C., Douay (2008) Acquisition de paramètres biométriques sur des isopodes tererrestres au moyen d’un téréomicroscope Zeiss. Ensemble, innovons. Institut catholique de Lille. 13p.
- Godet J.P., Demuynck S., Waterlot C., Follet A., Douay F., Leprêtre A., Pruvot C.(2009) Les isopodes terrestres comme indicateurs de la contamination des sols par les éléments métalliques. ADEME 2e rencontres nationales de la recherche sur les sites & sols pollués. 20, , Paris, France.
Liens externes
 Portail de la carcinologie et des crustacés
Portail de la carcinologie et des crustacés  Portail de la biologie marine
Portail de la biologie marine