Lambeosaurus
Lambeosaurus ou lambéosaure (ce qui signifie « lézard de Lambe ») est un genre éteint de dinosaures ornithopodes de la famille des hadrosauridés et de la sous-famille des lambéosaurinés. Il a vécu au Crétacé supérieur (Campanien) en Amérique du Nord, il y a environ entre 76 et 75 Ma (millions d'années)[1].

autrefois attribués à des espèces différentes,
mais aujourd'hui considérés comme appartenant
à l'espèce L. lambei des deux sexes
et à différents stades de sa croissance.
Espèces de rang inférieur
- † L. lambei (espèce type)
Parks, 1923 - † L. magnicristatus
Sternberg, 1935
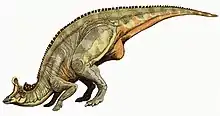
Ce dinosaure herbivore bipède ou quadrupède, est surtout connu pour sa crête crânienne creuse qui, chez l'espèce la plus connue (L. lambei) ressemblait à une hache. Plusieurs espèces ont été créés pour les fossiles trouvés dans l'Alberta (Canada), le Montana (États-Unis), et la Basse-Californie (Mexique), mais seules les deux espèces canadiennes sont actuellement reconnues comme valides. Mesurant environ 12,5 mètres de long[réf. nécessaire], l'espèce mexicaine L. laticaudus, depuis reclassée dans son propre genre Magnapaulia, a été l'un des plus grands genres d'ornithischiens. Les autres espèces étaient de taille plus modeste.
Lambeosaurus a été décrit tardivement en 1923 par William Arthur Parks, plus de vingt ans après l'étude des premiers fossiles découverts par Lawrence Lambe. Le genre a eu une histoire taxonomique compliquée, en partie parce que les petits hadrosauridés à aigrettes, maintenant connus comme étant des jeunes, étaient autrefois considérés comme des genres et espèces à part entière. Actuellement, les divers crânes attribués à l'espèce type L. lambei sont interprétés comme des crânes d'âges différents et/ou montrant un dimorphisme sexuel. Lambeosaurus était étroitement lié au genre mieux connu Corythosaurus, dont les fossiles se trouvent dans des sédiments un peu plus âgés[1],[2]. Sa proximité avec les genres moins connus Hypacrosaurus et Olorotitan, mise en avant un temps, a été réfutée en 2016[2]. Tous possédaient cette crête originale, qui est maintenant généralement considérée comme ayant servi à des fonctions sociales, de communication phonique et de reconnaissance visuelle.
Description

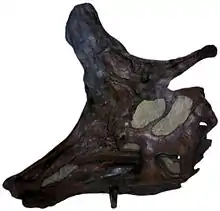
Lambeosaurus, au moins pour le plus connu d'entre eux, L. lambei, était assez similaire au célèbre Corythosaurus, n'en différant que par la forme de l'ornement de la tête. Comparée à celle de Corythosaurus, la crête de Lambeosaurus était placée plus vers l'avant, et les tuyaux nasaux creusés en son sein étaient placés à l'avant de la crête et empilés verticalement[3]. Il peut également être distingué de Corythosaurus par l'absence de bifurcation de ses processus nasaux qui formaient une partie des côtés de la crête, ce qui est la seule façon de différencier les juvéniles des deux genres. Les crêtes prenaient ensuite des formes distinctives chez les animaux plus âgés[4].
Dans l'ensemble, Lambeosaurus était un hadrosauridé comme les autres ; il pouvait se déplacer soit sur ses deux pattes arrière soit sur ses quatre pattes, comme retrouvé par des empreintes fossilisées de leur passage. Il avait une longue queue raidie par des tendons ossifiés qui l'empêchait de tomber. Les mains avaient quatre doigts, ayant perdu le pouce retrouvé normalement chez les tétrapodes, tandis que les deuxième, troisième et quatrième doigts étaient regroupés et formaient des sabots, ce qui évoque que l'animal pouvait utiliser ses membres antérieurs pour se soutenir. Le cinquième doigt était libre et pouvait être utilisé pour manipuler des objets. Chaque pied avait seulement les trois orteils centraux[5]
.
La crête, la caractéristique la plus distinctive, était différente chez les deux espèces les mieux connues. Chez L. lambei, elle avait une forme de hache lorsque le dinosaure était adulte, et était un peu plus courte et plus arrondie chez les fossiles considérés comme des femelles[3]. La « lame de la hache » se projetait devant les yeux, et le « manche » était une tige osseuse solide qui surplombait l'arrière du crâne. La lame de la hache avait deux parties : La partie supérieure était une mince crête osseuse qui apparaissait assez tard dans la vie, quand un individu approchait l'âge adulte, et la partie inférieure contenait les espaces creux qui étaient en continuation avec les voies nasales[3]. Chez L. magnicristatus, le « manche » était considérablement réduit, et la « lame » fortement élargie[6], formant une très grande banane. Cette crête est endommagée chez la plupart des spécimens entiers, et on n'en retrouve généralement que la moitié antérieure[7]. Chez les espèces moins connues L. laticaudus et L. paucidens la crête n'est pas encore connue. Toutefois, L. laticaudus peut se reconnaître par sa grande taille et l'importante hauteur de sa queue, qui avait de longs arcs hémaux ou chevrons et processus épineux analogues à ceux des hypacrosaures[8].
Les espèces canadiennes de Lambeosaurus semblent avoir eu la même taille que les Corythosaurus, et donc mesurer environ 9 à 9,40 mètres de long[9],[10]. On a des empreintes d'écailles de plusieurs spécimens ; un spécimen aujourd'hui classé comme L. lambei avait une peau fine avec des écailles de même taille, polygonales réparties sans ordre particulier sur le cou, le torse et la queue[11]. Une structure similaire est connue pour le cou, les membres antérieurs, et les pieds d'un spécimen de L. magnicristatus[7].
Classification
Lambeosaurus est le genre type des Lambeosaurinae, la sous-famille des hadrosauridés qui avait une crête creuse sur la tête. Dans les lambéosaurinés, il est étroitement lié à Corythosaurus, n'en différant de très peu, que par la forme de la crête[5]. La position phylogénétique de Lambeosaurus au sein des lambeosaurinés a évolué avec le temps ; dans les années 2000, il a été placé dans une tribu, les Corythosaurini, dont l'usage s'est perdu aux dépens du clade des Lambeosaurini au début des années 2010[12],[13].
Les grandes synthèses phylogénétiques sur les Hadrosauroidea réalisées par Albert Prieto-Márquez et ses collègues à compter de 2010 le placent dans la tribu des Lambeosaurini[12],[1],[2].
Cladogramme
Le cladogramme d'Albert Prieto-Márquez et ses collègues, réalisé en 2013, indique sa position entre la tribu des Parasaurolophini et le genre Corythosaurus[1] :
| Lambeosaurinae |
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Un cladogramme plus large, plus complet et plus récent, établi par les mêmes auteurs en 2016, se trouve dans l'article Hadrosauridae[2].
Découverte et histoire

Lambeosaurus a une histoire taxonomique compliquée, qui commence en 1902 lorsque Lawrence Lambe donne le nom de Trachodon marginatus a du matériel composé d'os des membres et d'autres os d'hadrosauridés (à l'origine CGC 419) de l'Alberta[14]. Les paléontologues ont commencé à trouver de meilleurs restes d'hadrosauridés dans les mêmes couches sédimentaires dans les années 1910, dans ce qui est maintenant connu comme un niveau stratigraphique datant de la fin du Campanien (Crétacé supérieur) dans la formation de Dinosaur Park. Lambe a attribué deux nouveaux crânes à T. marginatus puis, sur la base des nouvelles informations, a créé le genre Stephanosaurus pour cette espèce en 1914[15]. Malheureusement, il n'y avait que très peu de moyens d'associer ces crânes avec le matériel disparate de T. marginatus ; donc, en 1923, William Parks a proposé de créer un nouveau nom de genre et d'espèce pour les crânes, en leur donnant à la fois un nom générique et spécifique honorant Lambe : Lambesaurus lambei (spécimen type NMC 2869, à l'origine de la CGC 2869)[16].
De nouvelles espèces et les « prochénéosaures »
Bien que les premiers chercheurs de l'Alberta ne s'en soient pas rendu compte à l'époque, ils avaient découvert aussi les restes de Lambeosaurus juvéniles. Ces fossiles ont été interprétés comme des adultes d'une lignée distincte d'hadrosauridés, la sous-famille des Cheneosaurinae[17]. Le premier animal à être nommé a été Didanodon altidens, une mâchoire supérieure gauche (CGC 1092) provenant de la formation de Dinosaur Park et décrite par Henry Fairfield Osborn en 1902[18]. Elle a été renommée Trachodon altidens par Lambe[14], et le nom de Didanodon semble être rapidement tombé dans l'oubli, car il n'est pas mentionné dans la monographie sur les hadrosauridés d'Amérique du Nord de Richard Swann Lull et Nelda Wright en 1942.

En 1920, William Matthew Diller utilise le nom de genre Procheneosaurus (sans épithète spécifique) comme légende d'une photo d'un squelette de l’American Museum of Natural History, trouvé dans la formation de Dinosaur Park (AMNH 5340)[19]. Parcs estimant la procédure et la description insuffisantes pour que le nom soit considéré comme valide, veut remédier à la situation et invente le nom de genre Tetragonosaurus. C'est dans ce genre qu'il place l'espèce type T. praeceps (basée sur ROM 3577), une seconde espèce T. erectofrons (basé sur ROM 3578) pour des petits crânes trouvés également dans la formation de Dinosaur Park, et place Procheneosaurus dans T. praeceps[20]. Charles Mortram Sternberg complète la collection en 1935 par l'ajout du légèrement plus grand T. cranibrevis (basé sur CGC (aujourd'hui NMC 8633)[21]. L'utilisation du nom de genre Tetragonosaurus a été rejetée par Lull en faveur de Procheneosaurus. En 1942, avec Wright, il transfère les différentes espèces de Tetragonosaurus dans le genre Procheneosaurus, y ajoutant provisoirement Trachodon altidens et faisant de P. praeceps l'espèce type[22]. Ce choix a été généralement adopté jusqu'en 1975, quand Peter Dodson a montré que les « chénéosaures » étaient en réalité des jeunes d'autres espèces de dinosaures[3]. Procheneosaurus praeceps et P. altidens sont considérés comme des synonymes probables de Lambesaurus lambei[5], alors que les deux premiers noms sont antérieurs au troisième.
Les « prochénéosaures » n'ont pas été les seuls lambéosaurinés à être étudiés et nommés dans les années 1900. On a ensuite accepté la pratique de donner de nouveaux noms de genres et d'espèces pour ce qui est maintenant considéré comme de très probables variations individuelles, variation due à l'âge ou au sexe, ou à une distorsion de fossilisation. En 1935, trois autres noms d'espèces ont ainsi été créés pour le seul genre Lambeosaurus. Sternberg, dans un même article crée en plus de T. cranibrevis, L. magnicristatum (corrigé en magnicristatus), à partir d'un crâne et d'un squelette post-crânien partiel (CGC-maintenant NMC-8705) et L. clavinitialis, à partir d'un plus petit crâne (CGC-maintenant NMC-8703) avec une crête moins importante et des processus épineux plus réduits au niveau du dos[21]. Parcs a, pour sa part, créé Corythosaurus frontalis, (basé sur le crâne CGC 5853, aujourd'hui ROM 869), qui différait des célèbres grandes crêtes, érigées, arrondies des autres spécimens de Corythosaurus par une simple crête vers l'avant[23].
Réexamen et réorganisation du genre
Il n'y aura pas de nouveaux spécimens décrits pendant de nombreuses années. En 1964, John Ostrom note qu'une espèce nommée jadis par Othniel Charles Marsh, Hadrosaurus paucidens, (basée sur USNM 5457 composée d'un maxillaire partiel et d'un squamosal) trouvés dans la formation de Judith River dans le comté de Fergus, au Montana, était probablement un spécimen de Lambeosaurus[24].
En 1975, Peter Dodson, après avoir cherché à comprendre comment il y a pu voir tant d'espèces et de genres de lambéosaurinés vivant dans sur un laps de temps géologique aussi court et une aussi petite surface, publie les résultats d'une étude morphométrique basée sur la mesure de dizaines de crânes. Il constate que de nombreuses espèces ont été décrites à partir de restes qui doivent êtreinterprétés comme des jeunes ou des individus de l'autre sexe. Ainsi, pour Lambeosaurus, il constate que L. clavinitialis était probablement une femelle de L. lambei et Corythosaurus frontalis et Procheneosaurus praeceps des juvéniles de la même espèce. L. magnicristatus était cependant suffisamment différent pour justifier sa propre espèce. Il considère Procheneosaurus cranibrevis et P. erectofrons comme de jeunes corythosaures[3]. Reprenant l'étude du genre Procheneosaurus / Tetragonosaurus il y trouve beaucoup de confusion et notamment que le spécimen type de P. cranibrevis était un jeune Lambeosaurus, tandis que d'autres étaient des Corythosaurus, cela en se basant sur la jonction caractéristique de l'os nasal avec le prémaxillaire[4].

Pendant les années 1970, Bill Morris étudie les restes de lambeosaurinés géants de Basse Californie. Il les nomme ? L. laticaudus en 1981 (spécimen type LACM 17715). Morris place un point d'interrogation avant le nom, car il n'a découvert aucune crête entière pour son espèce, et ne peut donc en lui attribuer une affectation définitive. À partir de ce qu'il connait du crâne, il considère toutefois qu'il s'agit d'un Lambeosaurus. Il interprète cette espèce comme aquatique, en raison de caractéristiques comme sa grande taille, sa haute queue étroite (interprétée comme une adaptation à la natation), et la faiblesse des articulations de ses hanches ainsi que par le fait qu'un fémur cassé avait pu guérir. Il pensait qu'un tel handicap aurait été incompatible avec une aussi longue survie pour un animal terrestre[8].
Liste des espèces
Deux espèces de Lambeosaurus sont actuellement confirmées comme valides, une troisième et une quatrième sont parfois ajoutées.
Lambeosaurus lambei (Parks, 1923)
Elle est connue par au moins 17 individus, avec sept crânes et squelettes partiels et une dizaine de crânes isolés. L. clavinitialis (CM Sternberg, 1935), Corythosaurus frontalis (Parks, 1935), et Procheneosaurus praeceps (Parks, 1931) sont tous considérés comme des synonymes de L. lambei dans une revue de 2001[5]. Il est possible que les crânes de L. clavinitialis retrouvés sans colonne vertébrale associée soient en fait des L. magnicristatus[25], mais il faut savoir cependant que cette hypothèse a été rejetée dans la nouvelle description en 2007 de L. magnicristatus[7].
Lambeosaurus magnicristatus (Sternberg, 1935)
Elle n'est connue qu'à partir de deux individus, tous deux avec leur crâne. Malheureusement, la majorité du squelette, hors la tête, du spécimen type a été perdu. Beaucoup d'os ont été fortement endommagés par l'eau pendant leur entreposage et ont été perdus avant leur description. Ses restes proviennent de séries géologiques légèrement plus jeunes que celle ayant L. lambei[7]. Le nom spécifique est dérivé du latin magnus « grand » et cristatus « aigrettes », en référence à leur crête osseuse[26]. En outre, Jack Horner a identifié des fragments de mâchoires de lambéosaurinés dans la formation de Bearpaw au Montana comme pouvant appartenir à L. magnicristatus ; ils représentent les premiers restes de lambéosaurinés trouvés dans des roches marines[27].
« Lambeosaurus laticaudus » (Morris, 1981)
Ce grand dinosaure, connu à partir de restes dissociés de plusieurs individus, a été un temps accepté comme une espèce appartenant au genre Lambeosaurus[5], ou une espèce d’Hypacrosaurus[7],[28]. Sa validité a été contestée par le descripteurs du genre Velafrons, qui estiment que les restes trouvés ne permettent pas de connaitre le genre mais seulement de dire qu'ils n'appartiennent pas à leur genre[28]. Cette espèce a finalement été attribuée en 2012 à un nouveau genre sous le nom binominal de Magnapaulia laticaudus [29]. Son nom spécifique est dérivé du latin latus « large » et de caudus « queue »[26].
« Lambeosaurus paucidens » (Marsh, 1889)
Elle est considérée comme un nom douteux et est répertorié comme Hadrosaurus paucidens dans une révision de 2004[5], mais au moins un auteur, Donald F. Glut, l'accepte toujours comme une espèce de Lambeosaurus[30]. L'espèce n'est plus prise en compte dans les synthèses phylogénétiques plus récentes entre 2010 et 2016[12],[1],[2]. L'épithète spécifique est dérivé du latin paucus : « quelques, peu » et dens de "dents"[26].
Autres espèces
Les particularités de Procheneosaurus cranibrevis et son identification comme une jeune lambeosauriné, ont été reconnues en 2005[4], mais n'ont pas encore eu une large diffusion. Enfin, Didanodon et Trachodon altidens ont reçu un traitement prêtant à confusion dans la revue de 2004 : Didanodon a été classé, sans commentaire, comme un Lambeosaurus, alors que Trachodon altidens a été inscrit pour sa part dans la liste des noms douteux[5].
Paléoécologie

Lambesaurus lambei et L. magnicristatus, découvertes dans la formation de Dinosaur Park, étaient membres d'une faune diversifiée et bien documentée d'animaux préhistoriques comprenant des animaux aussi bien connus que les dinosaures à cornes Centrosaurus, Styracosaurus et Chasmosaurus, d'autres hadrosauridés comme Prosaurolophus, Gryposaurus, Corythosaurus et Parasaurolophus, le tyrannosauridé Gorgosaurus et les ankylosauriens Edmontonia et Euoplocephalus[31]. La formation de Dinosaur Park est considérée comme une ancienne plaine avec des rivières et des zones inondables qui sont devenues des marécages et ont été soumises progressivement à des conditions marines lorsque la Voie maritime intérieure de l'Ouest a transgressé vers l'ouest[32]. À l'époque, le climat était plus chaud que dans l'Alberta actuel. Il n'y avait pas de gelées, mais une alternance de saisons humides et de saisons plus sèches. Les conifères formaient apparemment la végétation dominante, avec un sous-étage de fougères, de fougères arborescentes, et d'angiospermes[33]. Anatomiquement semblables, L. lambei, L. magnicristatus et Corythosaurus ont vécu à des périodes différentes au cours du dépôt de la formation de Dinosaur Park, comme le montre la stratigraphie. Les fossiles de Corythosaurus sont trouvés dans les deux tiers inférieurs de la formation, ceux de L. lambei sont présents dans le tiers supérieur, et les rares restes de L. magnicristatus sont retrouvés seulement au sommet, au moment où l'influence marine était la plus importante[34].
Paléobiologie
Alimentation
Comme les autres hadrosauridés, Lambeosaurus était un grand herbivore bipède et quadrupède, qui, pour broyer des plantes avait un crâne suffisamment évolué pour que les mâchoires puissent effectuer des mouvements de mastication analogues à ceux des mammifères. Ses dents étaient constamment remplacées et étaient regroupées dans des « batteries » dentaires contenant chacune plus de 100 dents, seule une poignée d'entre elles étant utilisées à un moment donné. Il utilisait son bec pour cueillir les végétaux, qui étaient ensuite maintenus entre les mâchoires par un organe analogue à des joues. Il pouvait se nourrir à partir du sol jusqu'à environ 4 mètres de hauteur[5]. Comme le fait remarquer le paléontologue Robert Bakker, les lambéosaurinés ont leur bec plus étroit que celui des saurolophinés (ex-hadrosaurinés), ce qui implique que Lambeosaurus et ses parents pouvaient se nourrir de manière plus sélective que leurs homologues à large bec[35].
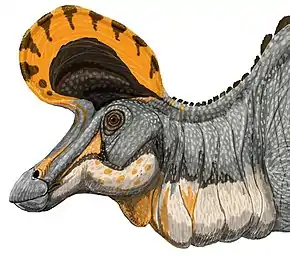
Crête
Comme les autres lambéosaurinés tels que Parasaurolophus et Corythosaurus, Lambeosaurus avait une crête caractéristique sur le dessus de la tête. Ses cavités nasales se prolongeaient en arrière dans cette crête, dont la plus grande partie était creuse. De nombreuses suggestions ont été faites pour la ou les fonctions de la crête, y compris d'avoir servi de réceptacle aux glandes à sel, à l'amélioration du sens de l'odorat, à servir de tuba ou de piège à air, agissant comme une caisse de résonance pour amplifier des sons, ou à permettre aux différentes espèces ou aux sexes différents de la même espèce de se reconnaître[25],[36]. À l'heure actuelle, le rôle social d'avertisseur sonore et de signe de reconnaissance est devenu le plus largement accepté parmi ces différentes hypothèses[5].
La grande taille des orbites et la présence d'anneaux sclérotiques impliquent qu'ils avaient une vue perçante et des habitudes diurnes, la vue étant importante pour ces animaux. Le sens de l'ouïe des hadrosauridés semble également avoir été très développé. Il y a au moins un exemple, dans le genre voisin Corythosaurus, de la présence d'un mince étrier (os retrouvé dans l'oreille des reptiles) qui, combiné à un grand tympan implique l'existence d'une oreille moyenne sensible, et le labyrinthe membraneux des hadrosauridés est allongé comme celui des crocodiliens. Cela indique que la partie auditive de l'oreille interne était bien développée[25]. Si elle était utilisée comme caisse de résonance, la crête pourrait également avoir permis d'émettre des sons différents reconnaissables par les différentes espèces ou sexes, car les dispositions différentes des voies nasales correspondant aux différentes formes de crête auraient permis de produire des sons intrinsèquement différents[37].
Notes et références
Notes
- (en) Cet article est partiellement ou en totalité issu de l’article de Wikipédia en anglais intitulé « Lambeosaurus » (voir la liste des auteurs).
Références
- (en) Albert Prieto-Márquez, Fabio M. Dalla Vecchia, Rodrigo Gaete et Àngel Galobart, « Diversity, Relationships, and Biogeography of the Lambeosaurine Dinosaurs from the European Archipelago, with Description of the New Aralosaurin Canardia garonnensis », PLoS ONE, vol. 8, no 7, , e69835 (DOI 10.1371/journal.pone.0069835)
- (en) A. Prieto-Marquez, G.M. Erickson et J.A. Ebersole, « A primitive hadrosaurid from southeastern North America and the origin and early evolution of 'duck-billed' dinosaurs », Journal of Vertebrate Paleontology, vol. 36, no 2, , e1054495 (DOI 10.1080/02724634.2015.1054495)
- (en) Peter Dodson, « Taxonomic implications of relative growth in lambeosaurine dinosaurs », Systematic Zoology, vol. 24, no 1, , p. 37–54 (DOI 10.2307/2412696, JSTOR 2412696)
- (en) David C. Evans, Dinosaur Provincial Park : A Spectacular Ancient Ecosystem Revealed, Bloomington, Indiana University Press, , 648 p. (ISBN 978-0-253-34595-0, lire en ligne), « The type specimen of Tetragonosaurus erectofrons (Ornithischia: Hadrosauridae) and the identification of juvenile lambeosaurines », p. 349–366
- (en) John R. Horner, Weishampel, David B. (éditeur), Dodson, Peter (éditeur) et Osmólska, Halszka (éditeur), The Dinosauria, Berkeley, University of California Press, , 2e éd. (ISBN 0-520-24209-2), « Hadrosauridae », p. 438–463
- (en) Richard Swann Lull, Hadrosaurian Dinosaurs of North America, Geological Society of America, , p. 193–194
- (en) David C. Evans, « Anatomy and relationships of Lambeosaurus magnicristatus, a crested hadrosaurid dinosaur (Ornithischia) from the Dinosaur Park Formation, Alberta », Journal of Vertebrate Paleontology, vol. 27, no 2, , p. 373–393 (DOI 10.1671/0272-4634(2007)27[373:AAROLM]2.0.CO;2)
- (en) William J. Morris, « A new species of hadrosaurian dinosaur from the Upper Cretaceous of Baja California: ?Lambeosaurus laticaudus », Journal of Paleontology, vol. 55, no 2, , p. 453–462
- Lull, Richard Swann; and Wright, Nelda E. (1942). Hadrosaurian Dinosaurs of North America. 227–228
- (en) Holtz, Thomas R. Jr. (2011) Dinosaurs: The Most Complete, Up-to-Date Encyclopedia for Dinosaur Lovers of All Ages, Winter 2010 Appendix.
- Lull, Richard Swann; and Wright, Nelda E. (1942). Hadrosaurian Dinosaurs of North America. 112–117
- (en)Prieto-Márquez, A. (2010). "Global phylogeny of Hadrosauridae (Dinosauria: Ornithopoda) using parsimony and Bayesian methods." Zoological Journal of the Linnean Society, 159: 435–502
- (en) R. M. Sullivan, S. E. Jasinski, M. Guenther and S. G. Lucas. 2011. The first lambeosaurin (Dinosauria, Hadrosauridae, Lambeosaurinae) from the Upper Cretaceous Ojo Alamo Formation (Naashoibito Member), San Juan Basin, New Mexico. Fossil Record 3. New Mexico Museum of Natural History and Science Bulletin 405-417
- (en) Lawrence M. Lambe, « On Vertebrata of the mid-Cretaceous of the Northwest Territory. 2. New genera and species from the Belly River Series (mid-Cretaceous) », Contributions to Canadian Paleontology, vol. 3, , p. 25–81
- (en) Lawrence M. Lambe, « On a new genus and species of carnivorous dinosaur from the Belly River Formation of Alberta, with a description of the skull of Stephanosaurus marginatus from the same horizon », Ottawa Naturalist, vol. 28, , p. 13–20
- (en) William A. Parks, « Corythosaurus intermedius, a new species of trachodont dinosaur », University of Toronto Studies, Geological Series, vol. 15, , p. 1–57
- (en) Lull, Richard Swann; and Wright, Nelda E. (1942). Hadrosaurian Dinosaurs of North America. 178–187.
- (en) Henry Fairfield Osborn, « Distinctive characters of the mid-Cretaceous fauna. 1. New genera and species from the Belly River Series (mid-Cretaceous) », Contributions to Canadian Paleontology, vol. 3, , p. 1–21
- (en) William Diller Matthew, « Canadian dinosaurs », Natural History, vol. 20, no 5, , p. 1–162
- (en) William A. Parks, « A new genus and two new species of trachodont dinosaurs from the Belly River Formation of Alberta », University of Toronto Studies, Geological Series, vol. 31, , p. 1–11
- (en) Charles M. Sternberg, « Hooded hadrosaurs of the Belly River Series of the Upper Cretaceous », Canada Department of Mines Bulletin (Geological Series), vol. 77, no 52, , p. 1–37
- (en) Lull, Richard Swann; and Wright, Nelda E. (1942). Hadrosaurian Dinosaurs of North America, 1–242
- (en) William A. Parks, « New species of trachodont dinosaurs from the Cretaceous formations of Alberta », University of Toronto Studies, Geological Series, vol. 37, , p. 1–45
- (en) John H. Ostrom, « The systematic position of Hadrosaurus (Ceratops) paucidens Marsh », Journal of Paleontology, vol. 38, no 1, , p. 130–134
- (en) James A. Hopson, « The evolution of cranial display structures in hadrosaurian dinosaurs », Paleobiology, vol. 1, no 1, , p. 21–43 (JSTOR 2400327)
- (en) D.P. Simpson, Cassell's Latin Dictionary, Londres, Cassell Ltd., , 5e éd. (ISBN 978-0-304-52257-6), p. 883
- (en) John R. Horner, « Upper Cretaceous dinosaurs from the Bearpaw Shale (marine) of south-central Montana with a checklist of Upper Cretaceous dinosaur remains from marine sediments in North America », Journal of Paleontology, vol. 53, no 3, , p. 566–577
- (en) Terry A. Gates, « Velafrons coahuilensis, a new lambeosaurine hadrosaurid (Dinosauria: Ornithopoda) from the Late Campanian Cerro del Pueblo Formation, Coahuila, Mexico », Journal of Vertebrate Paleontology, vol. 27, no 4, , p. 917–930 (DOI 10.1671/0272-4634(2007)27[917:VCANLH]2.0.CO;2)
- (en) A. Prieto-Márquez, L. M. Chiappe et S. H. Joshi, « The lambeosaurine dinosaur Magnapaulia laticaudus from the Late Cretaceous of Baja California, Northwestern Mexico », PLoS ONE, vol. 7, no 6, , e38207 (PMID 22719869, PMCID 3373519, DOI 10.1371/journal.pone.0038207)
- (en) Donald F. Glut, Dinosaurs : The Encyclopedia, Jefferson, North Carolina, McFarland & Co, , 1076 p., relié (ISBN 978-0-89950-917-4), « Lambeosaurus », p. 525–533
- (en) Weishampel, David B.; Barrett, Paul M.; Coria, Rodolfo A.; Le Loeuff, Jean; Xu Xing; Zhao Xijin; Sahni, Ashok; Gomani, Elizabeth, M.P.; and Noto, Christopher R. (2004). "Dinosaur Distribution", in The Dinosauria (2nd), pp. 517–606.
- (en) Eberth, David A. 2005. "The geology", in Dinosaur Provincial Park, pp. 54–82
- (en) Braman, Dennis R., and Koppelhus, Eva B. 2005. "Campanian palynomorphs", in Dinosaur Provincial Park, pp. 101–130
- (en) Michael J. Ryan, Dinosaur Provincial Park : A Spectacular Ancient Ecosystem Revealed, Bloomington, Indiana University Press, , 648 p. (ISBN 978-0-253-34595-0, lire en ligne), « Ornithischian Dinosaurs », p. 312–348
- (en) Robert T. Bakker, The Dinosaur Heresies : New Theories Unlocking the Mystery of the Dinosaurs and their Extinction, New York, William Morrow, , poche (ISBN 978-0-8217-2859-8), p. 194
- (en) David B. Norman, The Illustrated Encyclopedia of Dinosaurs : An Original and Compelling Insight into Life in the Dinosaur Kingdom, New York, Crescent Books, , 208 p. (ISBN 978-0-517-46890-6), « Hadrosaurids II », p. 122–127
- (en) David B. Weishampel, « Acoustic analyses of potential vocalization in lambeosaurine dinosaurs (Reptilia: Ornithischia) », Paleobiology, vol. 7, no 2, , p. 252–261
Annexes
Articles connexes
Liens externes
Références taxinomiques
- (en) Référence Paleobiology Database : Lambeosaurus Parks, 1923 (consulté le )
- (en) Référence uBio : Lambeosaurus Parks, 1923 (consulté le )
 Portail des dinosaures
Portail des dinosaures  Portail de l’Alberta
Portail de l’Alberta