Dinosauria
Les dinosaures (parfois dinosauriens), de leur nom scientifique Dinosauria (du grec ancien δεινός / deinós « terrible » et σαῦρος / saûros « lézard »[3],[alpha 1]), forment un super-ordre ainsi qu'un clade extrêmement diversifié de sauropsides de la sous-classe des diapsides et dont les uniques représentants actuels sont les oiseaux. Ce sont des archosauriens ovipares, ayant en commun une posture érigée et partageant un certain nombre de synapomorphies telles que la présence d'une crête deltopectorale allongée au niveau de l'humérus et un acetabulum perforant le bassin. Présents dès le milieu du Ladinien (deuxième étage du Trias moyen), il y a environ 240 Ma, la branche non avienne des dinosaures disparait entièrement lors de l'extinction Crétacé-Paléogène il y a 66 Ma. Les oiseaux, la branche avienne des dinosaures, a émergé de petits dinosaures théropodes du Jurassique supérieur et est la seule à avoir survécu à cette extinction, ce qui a permis leur diversification considérable à partir du Cénozoïque.
Ce groupe de vertébrés majoritairement terrestres connut un succès évolutif considérable au Mésozoïque, dominant les faunes continentales entre le Trias supérieur et le Crétacé supérieur pendant plus de 170 Ma. Présents sur l'ensemble des continents dès la fin du Trias, ils comprennent des formes très diverses d'animaux terrestres et volants, bipèdes et quadrupèdes, carnivores et herbivores, ayant développé toute une série d'innovations squelettiques et tégumentaires telles que des cornes, des crêtes, des plaques et des plumes. Les dinosaures non aviens comptent parmi eux les animaux les plus grands et les plus lourds ayant existé sur la terre ferme. Néanmoins, un grand nombre de dinosaures ne dépassait pas la taille d'un être humain et certains d'entre eux étaient plus petits qu'une poule.
La classification standard des dinosaures distingue deux grands clades selon la morphologie de leur bassin : les Ornithischia et les Saurischia. Les Ornithischia (ou Ornithischiens) ne comprennent que des dinosaures herbivores que les paléontologues divisent en trois groupes majeurs : les Ornithopoda qui regroupent des dinosaures majoritairement bipèdes dont les fameux dinosaures à « bec de canard » (ou Hadrosauridae), les Marginocephalia qui incluent des dinosaures à collerette et à dôme osseux sur le haut de la tête (respectivement les Ceratopsia et les Pachycephalosauria), et enfin les Thyreophora qui englobent des dinosaures quadrupèdes surmontés d'armures, de piques et de plaques osseuses sur le dos et la queue (les Ankylosauria et les Stegosauria). Les Saurischia (ou Saurischiens) sont divisés en deux clades bien distincts, les Theropoda qui comprennent des dinosaures bipèdes et l'entièreté des dinosaures carnivores et piscivores ainsi que les dinosaures à plumes et les dinosaures volants (les oiseaux dans leur grande majorité), et les Sauropodomorpha, des animaux généralement quadrupèdes et de très grande taille munis d'un long cou, d'une petite tête et d'une longue queue.
En 2017, une révision de cette classification est proposée par Matthew Baron et ses collègues, en regroupant d'une part les théropodes et les ornithischiens au sein d'un même clade (les ornithoscélidés[alpha 2]) et d'autre part les sauropodomorphes et les herrérasauridés au sein du clade des saurischiens (qui n'inclut donc plus les théropodes)[4],[5],[6].
Le terme Dinosauria fut proposé par le paléontologue anglais Richard Owen en avril 1842, qui donna l'étymologie de « lézard terriblement grand », même s'il est souvent affirmé que « dinosaure » signifie « terrible lézard » (Éric Buffetaut[7]). Dès le milieu du XIXe siècle et jusqu'à la fin des années 1960, les dinosaures non aviens furent considérés par les scientifiques comme des lézards géants, des reptiles (groupe aujourd'hui reconnu comme paraphylétique) à sang froid, patauds et lents, ayant disparu à la fin du Mésozoïque victimes de leur stupidité. Amorcée par le paléontologue américain John Ostrom, en 1969, la « renaissance des dinosaures » se caractérise par un regain d'intérêt pour l'étude des dinosaures disparus, qui furent dès lors reconnus comme des animaux actifs, probablement endothermes même si toutes les études ne sont pas d'accord, et ayant des comportements sociaux complexes.
Les restes des dinosaures non aviens suscitèrent dès leur découverte un grand intérêt auprès du public et les reconstitutions de squelettes dévoilées à l'occasion des expositions ont toujours été des attractions majeures dans les musées du monde entier. Les grands dinosaures disparus sont d'ailleurs devenus partie intégrante de la culture populaire aux XXe et XXIe siècles, figurant dans une pléthore de livres et des films à succès tels que Jurassic Park et L'Âge de glace 3 : Le Temps des dinosaures. Aujourd'hui, si une « dinomania » touche notamment certains enfants, la curiosité populaire ne s'est jamais démentie et les nouvelles découvertes régulièrement rapportées par les médias entretiennent une certaine fascination pour ces animaux. Le mot « dinosaure » est largement passé dans le langage courant et son usage souvent inconsidéré tend à attribuer faussement ce terme à d'autres animaux disparus comme les ptérosaures, les mosasaures, les plésiosaures voire même certains synapsides comme Dimetrodon ou Edaphosaurus qui étaient très éloignés des dinosaures.
Dinosauriens • Dinosaures
Cet article possède des paronymes, voir Dinosaur et Dinosaurus.
« Dinosaure » redirige ici. Pour les autres significations, voir Dinosaure (homonymie).
Pour les articles homonymes, voir Dinosauria (homonymie).

| Règne | Animalia |
|---|---|
| Embranchement | Chordata |
| Sous-embr. | Vertebrata |
| Super-classe | Tetrapoda |
| Classe | Sauropsida |
| Sous-classe | Diapsida |
| Clade | Archosauria |
| Clade | Ornithodira |
| Clade | Dracohors |
Super-ordre ou Clade
Ordres de rang inférieur
- † Ornithischia
- Saurischia
- † Herrerasauria[1],[2] ?
- † Sauropodomorpha
- Theropoda (inclut Aves)
Historique et étymologie
Étymologie

Le substantif masculin[8],[9],[10] dinosaure est emprunté[8],[9], par l'intermédiaire de l'anglais[8] dinosaur[9], au latin scientifique moderne dinosaurus[8],[9].
Le taxon Dinosauria a été introduit et défini par le paléontologue britannique Richard Owen en 1842 afin de regrouper une « tribu ou sous-ordre distinct de Reptiles Sauriens »[11]. Le terme dérive de deux racines grecques δεινός, deinos, signifiant, selon Owen, « effroyablement grand », et σαύρα, sauros, voulant dire « reptile » ou « lézard ». Dès l'apparition du nom, la première racine deinos fut incorrectement traduite par « terrible » ou « effrayant », bien qu'Owen n'utilisât pas cette racine comme un adjectif mais bien sa forme superlative, comme l'eût employée Homère dans l'Iliade, afin de révéler la nature impressionnante de ces animaux[12]. Le paléontologue anglais incluait alors dans ce nouveau groupe de reptiles au moins trois genres, Megalosaurus, Iguanodon et Hylaeosaurus, tous trois caractérisés, entre autres, par un large bassin (sacrum) formé de cinq vertèbres ankylosées, la grande hauteur des épines neurales des vertèbres dorsales et la double articulation des côtes des vertèbres[11]. Trois autres dinosaures préalablement nommés, le sauropode Cetiosaurus, le théropode Poekilopleuron et le sauropodomorphe basal Thecodontosaurus, ne furent cependant pas classés parmi les Dinosauria par Owen qui les considérait alors comme des reptiles indéterminés.
Interprétations mythologiques et courses aux ossements
.jpg.webp)

Les fossiles de grands animaux mésozoïques sont connus depuis l'aube de l'humanité, mais leurs véritables identifications n'ont émergé qu'après des millénaires d'interprétations mythologiques, et après plusieurs décennies de postulats fantaisistes même au sein d'une géologie et d'une paléontologie scientifiques, mais manquant, aux débuts, d'un nombre suffisant de données.
Pour les Chinois c'étaient des os de dragons, pour les Européens des restes des Géants bibliques et d'autres créatures tuées par le Déluge. Georges Cuvier décrit un lézard marin, le mosasaure (contemporain des dinosaures, mais qui n'en était pas un), dès 1808. Les premières espèces identifiées et baptisées sont l’iguanodon, découvert en 1822 par le géologue anglais Gideon Mantell, qui a remarqué des similitudes entre ses fossiles et les os de l’iguane contemporain. Le premier article scientifique sur les dinosaures paraît deux ans plus tard. Il est publié par le révérend William Buckland, professeur de géologie à l'université d’Oxford, et concerne Megalosaurus bucklandii, dont un fossile avait été découvert près d’Oxford. L'étude de ces « grands lézards fossiles » fait l'objet d'un grand intérêt dans les cercles scientifiques européens et américains, et le paléontologue anglais Richard Owen invente le terme « dinosaure » en 1842. Il remarque que les restes précédemment trouvés (Iguanodon, Megalosaurus et Hylaeosaurus) ont de nombreux caractères en commun, et décide de créer un nouveau groupe taxonomique. Avec l'aide du prince Albert de Saxe-Cobourg-Gotha, mari de la reine Victoria, il crée le musée d'histoire naturelle de Londres, à South Kensington (Natural History Museum), pour exposer la collection nationale de fossiles de dinosaures, ainsi que quelques autres objets d'intérêt botanique et géologique.
Vulgarisateur bénéficiant de gros budgets pour faire voir et faire toucher ce qu'il croit être une reconstitution exacte, Richard Owen rend populaire sa dénomination qui atteint un apogée dans les foules curieuses vers 1890. Le naturaliste Thomas Henry Huxley propose ornithoscélidés, littéralement « à la cuisse d'oiseaux ». Hermann de Meyer préfère pachypodes c'est-à-dire « aux pieds épais ». Ces dénominations ne quittent pas le cercle de lecteurs savants. Mais, après 1887, les spécialistes de sciences naturelles lassés du spectacle d'Owen, ordonnent avec rigueur les différentes familles de Saurischiens et d'Ornithischiens[réf. nécessaire].
En 1858 le premier fossile de dinosaure américain est découvert, dans des marnières près de la petite ville de Haddonfield, dans le New Jersey (ce n'est pas le premier fossile de dinosaure trouvé en Amérique, mais le premier identifié comme tel). L’animal est nommé Hadrosaurus foulkii, du nom de la ville et de son découvreur : William Parker Foulke. Cette découverte est capitale car il s'agissait du premier squelette presque complet découvert, et Joseph Leidy met en évidence sans doute possible que l'animal était bipède. Jusqu’alors la plupart des scientifiques croyaient que les dinosaures marchaient à quatre pattes comme les lézards.
La découverte de l'Hadrosaurus bipède marque le début effervescent d'une chasse aux fossiles de dinosaures aux États-Unis. La lutte acharnée entre Edward Drinker Cope, amateur fortuné de géologie et de paléontologie, natif de Philadelphie et Othniel Charles Marsh, heureux new-yorkais héritier de George Peabody à 21 ans et prospecteur infatigable, est connue sous le nom de « guerre des os » (Bone Wars), ou « ruée vers l'os », expression qui fait référence à la ruée vers l'or au XIXe siècle. Leur querelle irrémédiable après 1870, née d'une rivalité dans la chasse aux dinosaures démultipliée par de nombreuses brouilles dont Como Bluff, gisement mis au jour par deux ouvriers de l'Union Pacific et finalement exploité par O.C. Marsh, dure presque 30 ans, et finit en 1879 quand Cope mourut après avoir dépensé toute sa fortune dans cette quête. Marsh qui lui survécut vingt années sortit vainqueur, mais n'évita la ruine que grâce à l’aide financière inespérée de l’Organisation Géologique des États-Unis (United States Geological Survey). La collection d'Edward Cope, auteur prolifique de plus de 1800 articles scientifiques depuis ses dix-huit ans, ce qui ne l'empêcha pas de placer la tête sur la queue d'un plésiosaure, se trouve aujourd'hui au Musée américain d'histoire naturelle, à New York, celle de Marsh au musée Peabody d'histoire naturelle, à l'université Yale.
L'assistant de Cope, Charles Hazelius Sternberg aidé de ses trois fils, reprend le flambeau en entreprenant, au début du siècle suivant, des missions en Alberta, au Kansas, dans le Montana et le Wyoming. Dans ce dernier État, il relève en 1908 la première empreinte de peau momifiée de ce qui s'avère être un Anatosaurus. Une seconde découverte similaire est faite en 1910. Une partie de son équipe rejoint en 1913 Barnum Brown, qui, parfois avec son ami Henry Fairfield Osborn, prospecte depuis 1910 les berges de la Red Deer river entre l'Alberta et le Montana. Les barges des chercheurs, perturbés dans leur repérage au fil de l'eau par d'agaçants moustiques, dérivent avec le courant. Mais la moisson de trouvailles en vaut la peine : squelettes de théropodes, cératopsiens, hadrosaures[réf. nécessaire]...
Les chercheurs de l'Empire allemand annoncent la généralisation de la chasse lointaine et systématique, par leurs succès dans les colonies allemandes d'Afrique occidentale allemande et orientale. Dans l'actuelle Tanzanie, le site de Tendaguru livre des grands sauropodes de la famille des Brachiosauridés (Giraffatitan) et des petits stégosauriens genre Kentrosaurus aux paléontologues allemands. Werner Janensch supervise la collection rapatriée au Musée d'histoire naturelle de Berlin. Edwin Hennig poursuit les investigations en Afrique entre 1908 et 1912.
Si les chasseurs de dinosaures Cope et Marsh sont les véritables héros de la « guerre des os », cette bataille se poursuit de 1870 à 1910 avec plusieurs pays émergents comme les États-Unis et l'Allemagne qui veulent se mesurer aux grandes nations de l'époque, en finançant des expéditions dans le monde entier pour découvrir les dinosaures les plus spectaculaires par leur taille et leur étrangeté[14].
Le temps de l'analyse scientifique en Chine et dans le reste du monde
C'est grâce à une exploration asiatique à la quête de l'origine de l'homme que la Smithsonian Institution, par l'intermédiaire de son musée d'histoire naturelle, le Musée national d'histoire naturelle des États-Unis, cumule les trouvailles décisives dans le désert de Gobi entre 1922 et 1925. La mission du Musée américain d'histoire naturelle, à la suite d'une initiative d'Osborn et de Granger et commandée par Roy Chapman Andrews, découvre des œufs et des nids de Protoceratops andrewsi.
La Chine et ses confins avaient déjà attiré une expédition russe entre 1915 et 1917. Les missions russes, source de prestige, reprennent en 1946 et 1948. Les Chinois rentrent tôt dans la course et lancent leurs propres missions scientifiques après 1933. Après le refroidissement des relations sino-soviétiques, elles se cantonnent à la Mongolie. Les missions russo-mongoles et polono-mongoles se succèdent dans les années 1960 et 1970. Le Gobi est alors l'éden des chasseurs de dinosaures. Plus que les rares revues spécialisées, journaux et radio relatent les péripéties aventureuses.
La Chine investit avec force la spécialité de la paléontologie des vertébrés. Le temps du terrible dragon Kong-Hong reconnu dans les entrailles de la terre et vénéré laisse la place à l'analyse scientifique. Des spécialistes de renom émergent tels C. C. Young alias Yang Zhongjian et le professeur Dong Zhiming qui découvrent plus de cent espèces inconnues en Chine : Sauropode Mamenchisaurus, Hadrosaure Tsintaosaurus, Prosauropode Lufengosaurus, Stegosaure Tuojiangosaurus…
La connaissance de la faune du Mésozoïque s'accroît aussi avec l'expédition franco-norvégienne du Spitzberg en 1968, les paléontologues français y sont avantagés par la connaissance préalable de cette période géologique au Sahara et en Afrique du Nord[réf. nécessaire].
La recherche de fossiles s'est étendue à toute la surface du globe, y compris en Antarctique, où un ankylosaure, Antarctopelta, est découvert en 1986 sur l'île James-Ross, mais dont la description n'intervient que vingt ans après[15]. C'est cependant en 1994 qu'un dinosaure habitant réellement l'Antarctique, Cryolophosaurus ellioti, est décrit dans un journal scientifique[16]. Les zones particulièrement intéressantes sont aujourd'hui l'Amérique du Sud, surtout l'Argentine, et la Chine, dont le sous-sol a révélé de nombreux squelettes très bien conservés.
Dinosaures de France
- Trias de l'ouest et de l'est de la France :
- Airel en Manche : squelette de Coelurosaure, Halticosaurus.
- Pierreclos en Saône-et-Loire : restes de Plateosaurus
- La Chapoux (anciennement Le Chappou) à Cerin dans le département de l'Ain : Thecodontosaurus.
- Salins, Arbois, Poligny, Moissey et Domblans dans le Jura : Plateosaurus, Prosauropodes.
- Violot et Provenchères-sur-Meuse dans la Haute-Marne : Plateosaurus
- Lunéville et Saint-Nicolas-de-Port dans l'est de la Meurthe-et-Moselle : prosauropodes du genre Plateosaurus et Thecodontosaurus, dents de Coelurosaure.
- Jurassique :
- rivages normands, embouchure de la Seine et falaises du Pays de Caux, en particulier au voisinage de Caen, Villers-sur-Mer, Dives, Honfleur, du Havre : dinosaures, gros théropodes, Megalosaurus, Piveteausaurus ou Streptospondylus, coelurosaure, stégosaure Lexovisaurus, etc.,
- Boulogne-sur-Mer et le Boulonnais : restes des sauropodes, mégalosauridés, coelurosaures, Camptosaurus (ornithopode primitif),
- Damparis dans le Jura : restes de gros sauropode du genre Vouivria[17], mégalosauridés, etc.,
- Castelbouc, en Lozère : empreintes de sauropodes, au plafond d'une grotte ;
- Transition Jurassique-Crétacé :
- Canjuers dans le muséum départemental du Var : squelette du petit Compsognathus dans les calcaires lithographiques du Lagerstätte de Canjuers (1971), daté du Tithonien inférieur.
- Crétacé inférieur :
- Angeac en Charente : restes de sauropodes et ornithomimosaures ("dinosaures autruches").
- Pays de Bray et Villers-Saint-Barthélemy dans l'Oise : restes de sauropodes
- Wimereux en Pas-de-Calais : mégalosauridés
- Wassy en Haute-Marne, Cousancelles, Ville-sur-Saulx en Meuse, Louppy-le-Château, Varenne, Grandpré en Ardennes : mégalosauridés type Erectopus, iguanodons et iguanodontidés…
- Bedoin et Mondragon dans le Vaucluse : petits sauropodes
- Fons et Serviers-et-Labaume dans le Gard : dents de thérapode de la famille des deinonychosauridés.
- Crétacé supérieur du Languedoc, de la Provence ou de la façade ouest :
- Saint-Martory dans la Haute-Garonne : fragment de mandibule de hadrosauridés (identifiés après 1975)
- Villeveyrac, Saint-Chinian, Montpellier et Argelliers dans l'Hérault, Albières et Fa dans l'Aude : iguanodondidés Rhabdoton, hadrosauridés Orthomerus, sauropodes Hypselosaurus et Titanosaurus, ankylosaure Struthiosaurus, petits et grands thérapodes…
- Campagne-sur-Aude dans l'Aude : Squelette relativement complet d'Ampelosaurus atacis (Titanosauridae) (2000)
- Aix-en-Provence et contrées voisines dans les Bouches-du-Rhône et le Var : gisements d'œufs de dinosaures
- Près du Phare de Chassiron, île d'Oléron, Veillon en Vendée : empreintes de dinosaures, spectaculaires sur le dernier site…
- Terrains tertiaires comportant des sédiments mésozoïques remaniés par la mer des faluns
Définition
Définition phylogénétique

Afin de distinguer les dinosaures de leurs plus proches cousins (les dinosauromorphes), les scientifiques cladistes, qui définissent des groupes d'organismes sur base de leurs ancêtres plutôt que sur la possession des caractères anatomiques particuliers, ont défini les dinosaures comme étant les « membres du clade le moins inclusif comprenant Triceratops horridus et Passer domesticus » (le moineau domestique)[18],[19],[20].

Cette définition, dite phylogénétique ou cladiste, a besoin d'un contexte phylogénétique et ne peut être comprise qu'en utilisant un cladogramme, ou arbre phylogénétique, un arbre schématique illustrant les liens de parentés entre les êtres vivants et les groupes au sein desquels ils se classent. Ainsi, un animal est un dinosaure s'il se positionne, dans un cladogramme, dans ce groupe d'organismes comprenant à la fois le Triceratops, le moineau, et l'ensemble des descendants de leur ancêtre commun. En d'autres termes, toute espèce descendant de cet ancêtre particulier est par définition un dinosaure. Cet ancêtre peut être positionné dans le cladogramme en trouvant le Triceratops et le moineau (Passer) puis en suivant leurs branches jusqu'à ce qu'elles se rejoignent en un nœud particulier correspondant au nœud du clade des dinosaures.
Des variantes de cette définition phylogénétique des dinosaures existent dans la littérature. Fernando Novas fut le premier à donner une définition du taxon Dinosauria, un clade incluant l'ancêtre commun des Herrerasauridae et des Saurischia + Ornithischia, nommant le clade comprenant les Saurischia et Ornithischia Eudinosauria, un taxon qui n'a jamais été repris par la suite[21]. Un an plus tard, Kevin Padian et Cathleen May définirent le clade des Dinosauria comme étant l'ensemble des descendants du plus récent ancêtre commun des oiseaux et de Triceratops[18], la définition actuellement reconnue des dinosaures, reprise parfois de manière légèrement différente par d'autres auteurs comme le célèbre paléontologue américain Paul Sereno (Triceratops, Neornithes, leur plus récent ancêtre commun et l'ensemble de ses descendants)[22], George Olshevsky, qui souhaitait faire honneur à l'inventeur des dinosaures Richard Owen (le plus récent ancêtre commun de Megalosaurus et Iguanodon et l'ensemble de ses descendants)[23] et Fraser et ses collègues (Triceratops (représentant les Ornithischia) plus Aves (représentant les Saurischia) et l'ensemble des descendants de leur dernier ancêtre commun)[24].
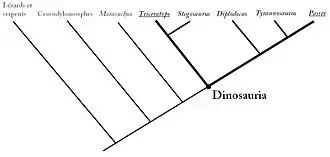
L'ensemble de ces définitions permet de regrouper des animaux définis comme étant des dinosaures (Dinosauria = Saurischia + Ornithischia) et comprenant les théropodes (dinosaures bipèdes, parfois carnivores, incluant les oiseaux), les sauropodomorphes (dinosaures herbivores majoritairement quadrupèdes munis d'un long cou), les ornithopodes (dinosaures herbivores majoritairement bipèdes comprenant les dinosaures à « bec de canard »), les ankylosauriens (dinosaures herbivores quadrupèdes munis d'armures osseuses sur le dos), les stégosauriens (dinosaures herbivores quadrupèdes portant des plaques osseuses et parfois des épines sur le dos), les cératopsiens (dinosaures herbivores majoritairement quadrupèdes possédant une crête osseuse à l'arrière du crâne et parfois des cornes sur le haut de la tête) et les pachycéphalosauridés (dinosaures herbivores bipèdes au crâne épaissi).
Il y a un consensus presque total chez les paléontologues sur le fait que les oiseaux sont les descendants des dinosaures, et plus précisément des dinosaures théropodes appartenant au clade des Maniraptora[25]. Suivant la définition phylogénétique (aussi appelée cladiste), les oiseaux sont donc des dinosaures et les dinosaures n'ont par conséquent pas disparu. Néanmoins, dans le langage courant, ainsi que pour les systématiciens de l'école évolutionniste[26], le mot « dinosaure » n'inclut pas les oiseaux. Par souci de clarté, cet article utilise ainsi le mot « dinosaure » comme synonyme de « dinosaure non avien ».
Caractères anatomiques identifiant les dinosaures
Les premiers scientifiques qui étudièrent les dinosaures ne leur avaient pas donné une telle définition puisque la théorie de l'évolution, impliquant la notion d'ancêtre commun, était vue à leur époque comme une hérésie. Ainsi, suivant la coutume de son époque, Richard Owen définit les dinosaures comme étant des animaux possédant un certain nombre de caractères anatomiques particuliers. Ces derniers sont actuellement nommés synapomorphies, des caractères dérivés, ou nouveautés évolutives, hérités d'un ancêtre commun et unissant l'ensemble des membres d'un clade. Les dinosaures possèdent plusieurs synapomorphies listées ci-dessous. Ces caractères dérivés partagés par l'ensemble des dinosaures permettent, non pas de définir les dinosaures, mais de les identifier, comme un médecin utiliserait une liste de symptômes pour diagnostiquer une maladie particulière[27].

- Musculature temporale s'étendant antérieurement sur le toit du crâne (= fosse supratemporale présente dans la partie antérieure de la fenêtre supratemporale)[20],[27],[28],[29]. Les muscles adducteurs mandibulaires, des muscles temporaux permettant la mastication chez les vertébrés, sont particulièrement larges et développés chez les dinosaures. Ainsi, la zone osseuse où s'attachent ces muscles sur le haut du crâne, dans la partie antérieure de la fosse temporale, est particulièrement étendue, alors qu'elle est courte chez les autres archosaures et les cousins très proches des dinosaures comme Silesaurus[27].
- Processus postérieur du jugal bifurque pour s'articuler avec le quadratojugal[20],[27],[28],[29],. Chez les archosaures autres que les dinosaures, le processus du jugal s'étendant postérieurement et s'articulant avec le quadratojugal est unique alors que chez les dinosaures, il est divisé en deux processus distincts[20],[27].
- Présence d'épipophyses chez les vertèbres cervicales antérieures (et non axiales)[20],[27],[28],[29]. Les épipophyses sont des projections osseuses positionnées dans la partie postérodorsale des épines neurales des vertèbres cervicales (sur la partie dorsale des postzygapophyses plus précisément). Elles permettent à certains muscles du cou de s'attacher afin de renforcer le cou et le dos. Chez les dinosaures, ces muscles devaient être particulièrement développés et capables d'une large variété de mouvements[27].
- Une crête deltopectorale allongée au niveau de l'humérus (occupant plus de 30 % la longueur de l'humérus)[20],[27],[28],[29]. La crête deltopectorale est une ride osseuse située sur la partie antérieure de l'humérus. Elle permet aux muscles deltoïdes de l'épaule et pectoralis de la poitrine de s'insérer. Chez les archosaures autres que les dinosaures, cette crête est discrète, alors qu'elle est longue et proéminente chez les dinosaures, témoignant de la grande mobilité et de la puissance de leurs membres antérieurs[27].
- Un acetabulum ouvert au niveau du bassin (= marge ventrale de l'acétabulum de l'ilion concave)[20],[27],[28],[29]. L'acetabulum est une cavité osseuse du bassin où s'insère et s'articule la tête du fémur. Chez l'homme, cette surface articulaire est circulaire afin de recevoir la tête arrondie du fémur, permettant une grande amplitude de mouvement. Chez les dinosaures, et contrairement à leurs proches cousins archosaures, l'acetabulum est ouvert, ou perforé. Une perforation de l'acetabulum témoigne d'une posture dressée, où les membres postérieurs se situent sous le corps. Chez les animaux à posture semi-dressée ou rampante, la surface médiale de l'acetabulum est renforcée par de l'os afin de dissiper les contraintes soumises par le poids de l'animal qui se propagent médialement vers le bassin, depuis la tête du fémur. Chez les dinosaures qui ont une posture dressée, la contrainte soumise par le poids de l'animal et exercée sur la surface articulaire de l'acetabulum est, non plus médiale, mais dorsale. Ainsi, la surface médiale osseuse de l'acetabulum, devenue inutile chez les archosaures à posture dressée, s'est vue disparaître au cours de l'évolution[27].
 Bassin d'Allosaurus (Theropoda) montrant l'acétabulum ouvert.
Bassin d'Allosaurus (Theropoda) montrant l'acétabulum ouvert. - Le quatrième trochanter du fémur forme une crête forte et asymétrique[20],[27],[28],[29]. Le quatrième trochanter est une crête osseuse située sur la partie postérieure du fémur et présent chez l'entièreté des archosaures. Ce trochanter permet l'insertion des muscles caudo-fémoraux de la jambe pour la locomotion. Chez la plupart des archosaures, le quatrième trochanter est peu développé et symétrique alors que chez l'entièreté des dinosaures, ce trochanter est protubérant et asymétrique, avec une partie distale plus étendue que la partie proximale. Bien que l'utilité d'un quatrième trochanter asymétrique est incertaine, le développement important de cette arête osseuse témoigne de membres postérieurs puissants et doués de mouvements efficients[27].

- Une facette articulaire de la fibula occupant moins de 30 % la largeur de l'astragale[20],[27],[28],[29]. L'astragale et le calcanéus sont les os proximaux du tarses (des os du pied) chez les archosaures. Chez les dinosaures et ses proches cousins, l'astragale est beaucoup plus grand que le calcanéum et les deux os sont fermement articulés entre eux, ainsi qu'avec le tibia et la fibula, l'ensemble de ces quatre os formant ainsi un complexe fonctionnel unique ne permettant aucun mouvement de rotation entre ceux-ci. Chez ces animaux, les mouvements entre la jambe et la patte se font entre le fémur, le complexe tarsal proximal (tibia + fibula + astragale + calcanéum) et le pied lui-même, une condition dite « mésotarsale » partagée par la lignée avienne des archosaures (Avemetatarsalia), qui se distingue de celle des pseudosuchiens où l'astragale et le calcanéum sont de taille semblable et où les deux os s'articulent de manière mobile. Les dinosaures se distinguent des autres métatarsaliens par un contact très réduit entre la fibula et l'astragale puisque la surface de contact entre les deux os n'occupe seulement que 30 % de la largeur de l'astragale[27].
Et selon Nesbitt (2011)[28] :
- Les exoccipitaux ne se rejoignent pas le long de la ligne du milieu de la surface de la cavité endocrânienne.
- Les radius occupent moins de 80 % la longueur la longueur de l'humérus.
- La crête cnémiale du tibia s'arque antérolatéralement.
- Présence d'une ride longitudinale distincte sur la face postérieure de la terminaison distale du tibia.
- La surface d'articulation avec la fibula est concave sur le calcanéus.
D'autres caractères ostéologiques ne se retrouvent que chez les dinosaures, mais puisque leur présence est incertaine chez leurs cousins les plus proches (dinosauromorphes), ils ne peuvent être utilisés avec certitude afin d'identifier un dinosaure d'un autre archosaure.
- Absence d'un os postfrontal au niveau du crâne[20],[28],[29].
- Plaques sternales ossifiées et doubles[20].
- Doigts IV et V de la main réduits[20].
- Trois phalanges ou moins au niveau du quatrième doigt de la main[20],[28],[29].
- Présence d'un foramen post-temporal[20],[28].
- Trois dents au niveau du prémaxillaire[28].
- Fosse nasale du prémaxillaire s'étendant dans le coin antéroventral de la narine[28].
- L'échancrure ventrale de la tête du fémur est concave[28].
- Absence d'un sillon postérieur sur l'astragale[28].
- Absence d'une tubérosité sur le calcanéum[28].
- Absence d'interclavicules[28].

- Une crête/fosse brevis sur la surface ventrale de l'ilion[20],[29]. La plupart des dinosaures possèdent une fosse distincte sur la surface latérale ou ventrale de l'ilion. C'est à cet endroit que s'attachaient une partie de la musculature caudofémorale dites "caudofemoralis brevis". La distribution de cette fosse est complexe puisqu'il n'est pas certain qu'il s'agisse d'un caractère homologue. Ainsi, les dinosaures basaux Herrerasaurus et Plateosaurus ne possèdent pas une telle fosse, au contraire de leur proche cousin Silesaurus et des dinosaures basaux Eocursor, Lesothosaurus, Panphagia et Saturnalia[20].
- Trois vertèbres sacrées ou plus[27]. La plupart des archosaures tels que les dinosauromorphes Lagerpeton et Marasuchus possèdent deux vertèbres sacrées au niveau du bassin. La grande majorité des dinosaures comprennent trois ou plus de trois vertèbres sacrées mais les saurischiens basaux Herrerasaurus et Staurikosaurus montrent deux vertèbres sacrées alors que Silesaurus, un taxon sœur des dinosaures en possèdent trois également[20],[27].
Plusieurs caractères anatomiques avaient été utilisés par plusieurs auteurs afin d'identifier les dinosaures. Cependant, avec la découverte de dinosaures basaux et de nouveau taxons proches des dinosaures, certains de ces caractères se sont révélés être partagés par des proches cousins des dinosaures, ou absent chez certains dinosaures basaux[20].
- Position dorsale de l'ectoptérygoïde par rapport à la crête transversale du ptérygoïde.
- Cavité glénoïde orienté postéro-ventralement au niveau de la scapula (omoplate) et du coracoïde.
- Contact entre le pubis et l'ischion réduit.
- Lamine médioventrale de l'ischion réduite.
- Tête du fémur tournée vers l'intérieur.
- Partie proximale du fémur avec une tubérosité médiale réduite.
- Présence d'un trochanter antérieur au niveau du fémur.
- Processus descendant du tibia s'insérant postérieurement par rapport au processus ascendant de l'astragale.
- Calcanéum proximal plat et concave.
- Présence d'une cheville métotarsale.
- Métatarses II et IV de longueur subégale.
- Terminaison distale du métatarse IV et plus long que large.
Description
Généralités
Bien que nécessaire, la liste des traits anatomiques énoncée ci-dessus et permettant de différencier un dinosaure d'un autre animal est rébarbative et emploie un grand nombre de termes anatomiques complexes et peu intéressants pour un public non spécialisé. Ainsi, un animal d'origine archosaurienne ayant vécu au Mésozoïque, ne volant pas et se différenciant des reptiles par une posture « érigée en contrefort » est très souvent, un dinosaure (non avien).

Puisque les dinosaures sont des archosaures, les mammifères disparus tels le Mammouth ne sont pas des dinosaures, de même que tout animal ayant vécu avant le Trias comme le Dimetrodon, un synapside non mammalien datant du Permien. Les dinosaures furent des animaux majoritairement terrestres et seuls quelques dinosaures à plumes proches de la lignée avienne comme Microraptor étaient capables de voler et d'occuper ainsi la niche écologique des airs. Par conséquent, les animaux aquatiques ayant vécu à la même époque que les dinosaures, comme les plésiosaures, les ichtyosaures et les mosasaures, n'étaient pas des dinosaures. De la même manière, et bien qu'ayant également vécu uniquement au Mésozoïque également, les ptérosaures, des proches cousins des dinosaures, exclusivement volants et dont l'aile est formée par une voile de peau soutenue par le quatrième doigt de la main qui s'est considérablement allongé, ne sont pas des dinosaures. Puisque les dinosaures possédaient des membres dressés, maintenus sous le corps, les animaux proches des dinosaures à posture semi-dressée ou rampante comme les reptiles, tels les crocodiles ou les lézards, ne sont pas non plus des dinosaures.
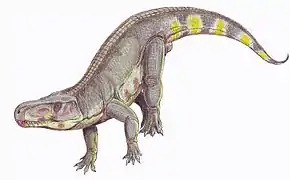
La posture dressée n'est pas une exclusivité dinosaurienne au sein de la lignée archosaurienne puisqu'elle est également partagée, en plus des proches cousins des dinosaures les dinosauromophes, par d'autres archosaures contemporains des premiers dinosaures : les Rauisuchia. La posture érigée de ces archosaures est néanmoins différente de celle des dinosaures (et des mammifères) dans la configuration anatomique du bassin. Chez les dinosaures (et les ornithodires d'une manière générale), la stature dressée fut acquise en gardant une position verticale des os du bassin et en tournant la tête du fémur médialement afin qu'elle s'articule dans la cavité de l'acetabulum qui s'ouvre latéralement[31], une posture dite « érigée en contrefort » ('buttress-erect' posture)[32]. Chez les rauisuchiens, la stature érigée, qui est apparue indépendamment de celle des dinosaures[33],[34], s'est développée grâce à une rotation de l'ilion vers une position horizontale, orientant ainsi l'acetabulum presque ventralement[33],[35],[36]. Cette configuration est connue sous le nom de posture « érigée en pilier » ('pillar-erect' posture)[32].
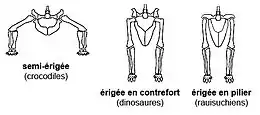
Il est nettement plus difficile de différencier les tout premiers dinosaures de leur proches cousins les dinosauromorphes puisque ces derniers sont également des archosaures bipèdes à posture « érigée en contrefort » ayant vécu à la même époque que les premiers dinosaures. Comme le montre la liste de caractères anatomiques propre aux dinosaures donnée ci-dessus, les dinosaures ne se différencient véritablement de leurs proches cousins que par une posture dressée plus rapide et plus effective, où les muscles du cou et des membres antérieurs et postérieurs furent plus développés et plus puissants, permettant aux premiers dinosaures d'avoir une bipédie effective et un mode de vie très actif.

Seule une posture dressée et des membres antérieurs et postérieurs pourvus de muscles puissants permettent de caractériser l'ensemble des dinosaures, qui furent de taille et de morphologie extrêmement diverses. En effet, les dinosaures, comprennent des animaux bipèdes et quadrupèdes, carnivores et herbivores, avec ou sans dents, à écaille, à poils (des proto-plumes) ou à plumes, aux membres antérieurs très développés ou très courts, des cous démesurément longs ou presque absents, et munis ou non de becs, collerettes, armures osseuses, plaques, cornes, épines, voiles, crêtes et dômes osseux.
Crâne et mâchoire

Le crâne et la mâchoire des dinosaures sont certainement les parties du squelette dont la forme est la plus variée. En effet, la morphologie de la tête, que ces deux entités anatomiques constituent, est directement liée au régime alimentaire, si bien que les dinosaures qui se nourrissaient de plantes coriaces, de feuilles, de viandes, de poissons ou filtraient les eaux pour se sustenter avaient des crânes et des mâchoires de forme très différente. Ainsi, les dinosaures théropodes qui comprennent des prédateurs puissants comme Tyrannosaurus[37], Acrocanthosaurus[38] et Torvosaurus[39] avaient des crânes massifs et robustes munis de dents tranchantes dentelées, alors que leur proches cousins piscivores les Spinosauridae comme Irritator[40] et Spinosaurus[41] avaient un crâne étroit et allongé portant de dents coniques dépourvues de denticules. Beaucoup de théropodes étaient également exclusivement herbivores[42] et possédaient des têtes à bec édenté comme certains Oviraptoridae tel que Citipati[43] ou des mâchoires à bec munies de petites dents triangulaires comme les Therizinosauroidea tel Erlikosaurus[44],[45],[46],[27],[47],[48].

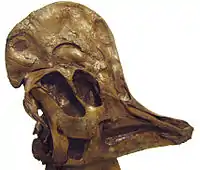
Parmi les dinosaures ornithischiens, qui furent exclusivement herbivores, le crâne était également souvent muni de becs osseux comme la plupart des cératopsiens et des hadrosaures dont les mâchoires portaient de véritables batteries dentaires[27]. Néanmoins, la forme du crâne des ornithischiens, en plus d'être liée au régime alimentaire, variait également fortement suivant les attributs de protections ou de parades comme les cornes, les dômes osseux, les crêtes et les collerettes qu'il portait. Ainsi, la tête des cératopsiens comme Styracosaurus[49], Pachyrhinosaurus[50] et Triceratops[51], qui fut munie d'une collerette (courte ou longue) et parfois de cornes nasales, frontales ou encore de cornes à l'extrémité de la collerette, se distinguait fortement de la tête épaisse et étroite des pachycéphalosauridés comme Stegoceras[52] ou Pachycephalosaurus[53] dont le crâne était épaissi et parfois bombé à son sommet. De même, la tête des hadrosaures, souvent pourvue de longues crêtes osseuses à l'extrémité du crâne comme chez Corythosaurus[54] et Parasaurolophus[55], ne pouvait être confondue avec la tête épaisse et courte des ankylosauridés comme Ankylosaurus[56] et Euoplocephalus[57] dont le sommet était protégé de protubérances osseuses et parfois rehaussé de courtes épines[45],[46],[27],[47],[48].

Malgré cette large variabilité de forme que déploie la tête des dinosaures, le crâne et la mâchoire sont toujours constitués des mêmes os et montrent toujours les mêmes ouvertures. En effet, les dinosaures sont des reptiles diapsides, des animaux dont le crâne est perforé par deux ouvertures crâniennes, ou fenêtres (également appelées fosses), dans la partie postérieure du crâne, la fenêtre temporale inférieure et la fenêtre temporale supérieure. De même, comme la majorité des vertébrés, les dinosaures possèdent une ouverture à l'emplacement de l’œil nommée orbite au milieu duquel se trouve l'anneau sclérotique permettant le support de l’œil, ainsi qu'une ouverture au niveau des narines, la fosse nasale d'où se terminaient les conduits respiratoires jusqu'aux narines. L'entièreté des saurischiens, et certains ornithischiens, montrent enfin une dernière fenêtre, généralement large, au niveau du crâne et située entre l'orbite et la fosse nasale. Cette ouverture, appelée fenêtre antéorbitaire, ce qui signifie "en avant de l'orbite", permettait le passage de plusieurs muscles reliant le toit du crâne aux mâchoires supérieures et inférieures. La mandibule est également pourvue d'une ouverture nommée fenêtre mandibulaire externe, présente chez la plupart des dinosaures[45],[46],[27],[47],[48].


Les os formant le crâne et la mâchoire, en grande majorité double (présents du côté gauche et droit) puisque la tête est symétrique, sont nombreux et leur morphologie est également très diverse au sein des différents groupes de dinosaures. Seuls les plus caractéristiques sont ici exposés. Le prémaxillaire se situe à l'extrémité antérieure du crâne et porte des dents ou un bec corné appelé ramphothèque, ou les deux. Néanmoins, chez les hadrosaures, il existe deux prémaxillaires qui contribuent à la crête osseuse portée par bon nombre d'entre eux. Le maxillaire est également, chez les dinosaures dentés, un os porteur de dents qui constitue la majeure partie de la mâchoire supérieure. Les os formant le toit du crâne sont l'os nasal (délimitant en partie la fosse nasale et formant parfois des crêtes osseuses de toutes sortes), l'os frontal, et l'os pariétal et squamosal qui se trouvent à l'arrière de la tête. Ces deux derniers sont extrêmement développés chez la plupart des cératopsiens puisqu'ils forment la large crête permettant de protéger le cou de ces animaux. La mandibule est majoritairement constituée de l'os dentaire qui est le seul à porter des dents au niveau de la mâchoire inférieure. Chez les dinosaures à bec comme les cératopsiens, les stégosauridés et les hadrosauridés, l'extrémité antérieure de la mâchoire se termine par un os supplémentaire directement relié au dentaire, l'os prédentaire, qui est également renforcé par une ramphothèque chez ces animaux à bec. De la même manière, chez les cératopsiens le prémaxillaire de la mâchoire supérieure est prolongé par un os supplémentaire portant le bec corné et appelé os rostral[45],[46],[27],[47],[48].
Squelette post-crânien

Si le squelette post-crânien (le squelette mis à part la tête) des dinosaures est moins différencié que le crâne et la mandibule, il n'en reste pas moins que sa morphologie est très variable au sein des différents clades de dinosaures. Directement lié à la locomotion et à l'alimentation, le squelette des dinosaures pouvait également déployer toute une série d'innovations squelettiques jouant des rôles de protection, de défense et de parade. Les dinosaures carnivores étaient ainsi exclusivement bipèdes, ce qui leur permettait de se déplacer avec agilité et rapidité pour attraper des proies. Leurs membres postérieurs furent nettement plus développés et robustes que les membres antérieurs, et les doigts de ces animaux se terminaient par des griffes acérées et parfois rétractiles. Au contraire, une majorité de dinosaures herbivores fut quadrupède et beaucoup d'entre eux étaient dotés de protubérances osseuses qui les protégeaient. Si la tête de certains Ceratopsidae déployaient des cornes, le corps des Stegosauridae portait des plaques osseuses triangulaires ou losangiques sur le haut du dos et parfois de longues épines à l'extrémité de la queue et sur les bas côté du ventre[45],[46],[27],[47],[48].

Les ankylosauriens adoptèrent une autre stratégie pour se prémunir des prédateurs. Chez ces animaux quadrupèdes, le corps était recouvert d'un bouclier osseux fait d'une imbrications de nodules, de piques et parfois de larges épines pointant vers les côtés. La queue de ces herbivores se terminait parfois également par une massue osseuse que l'animal était capable de balancer pour repousser les assaillants les plus téméraires. Un autre moyen de se protéger des prédateurs et vers lequel les Sauropodomorpha évoluèrent fut l'accroissement démesuré du corps, où le cou et la queue prirent des longueurs gigantesques. La grande dimension du cou permit à ces dinosaures quadrupèdes d'atteindre la cime des arbres les plus hauts pour se nourrir alors qu'une queue excessivement longue aida à contrebalancer le reste du corps et parfois également à jouer un rôle de fouet supersonique capable de claquer face aux carnassiers qui les attaquaient. Des nodules et épines osseuses se trouvaient également sur le dos de certains sauropodes titanosaures tels qu'Agustinia et Ampelosaurus, de même que certains théropodes carnivores comme Ceratosaurus. La longue queue de certains sauropodes se terminait également par une massue, comme le montre Shunosaurus et Omeisaurus, et parfois même de courtes épines telle la queue de Spinophorosaurus[45],[46],[27],[47],[48].

Les dinosaures herbivores bipèdes, comme la plupart des Ornithopoda, des Pachycephalosauridae et un grand nombre de théropodes maniraptoriformes, comptèrent surtout sur la robustesse et la vélocité de leurs membres postérieurs pour échapper aux prédateurs, mais certains herbivores comme Iguanodon portèrent également de larges éperons au niveau des pouces afin de poinçonner le cou des prédateurs. Bien que la plupart des théropodes comprenaient trois à quatre doigts à chaque main, quelques-uns d'entre eux comme Tyrannosaurus n'étaient pourvus que de deux doigts développés et un troisième doigt vestigial totalement réduit. Par ailleurs, les membres antérieurs de certains théropodes Alvarezsauridae ne furent terminés que par un doigt unique muni d'une large griffe. Les membres antérieurs des théropodes furent généralement bien développés, parfois même robustes comme chez les Spinosauridae, mais certains d'entre eux au sein de la lignée des Abelisauridae, Tyrannosauridae et Alvarezsauridae montraient des membres antérieurs extrêmement courts, leur réduction au cours du temps témoignant parfois d'une absence d'utilité. Dinosaures herbivores et carnivores pouvaient également déployer une voile osseuse sous-tendue par des épines neurales hypertrophiées sur le haut du dos. C'est le cas des dinosaures ornithopodes Ouranosaurus et Hypacrosaurus ainsi que les dinosaures théropodes Acrocanthosaurus, Concavenator et Spinosaurus[45],[46],[27],[47],[48].

Une fois encore, cette large variabilité de forme du corps des dinosaures ne dérive que de la transformation d'un certain nombre d'os, et la toute grande majorité des os du squelette des dinosaures fut partagée par l'ensemble d'entre eux et configurée de la même manière. Ainsi, la colonne vertébrale comprend toujours un nombre (parfois variable) de vertèbres cervicales (vertèbres du cou), dorsales (du dos), sacrales (du bassin) et caudales (de la queue). La cage thoracique est protégée par les côtes (cervicales et dorsales) et des os fins et parallèles nommés gastralias au niveau du bas-ventre, et les petits os situés sous les vertèbres caudales et permettant aux muscles de la queue de s'y attacher sont appelés chevrons ou arcs hémaux. Les membres antérieurs incluent un humérus (partie proximale de l'avant-bras), un radius et une ulna (partie distale de l'avant-bras) et enfin les os de la main que sont les carpes, métacarpes et phalanges. De même, les membres postérieurs se divisent en fémur (partie proximale de la jambe), tibia et fibula (partie distale de la jambe) et enfin des os du pied, à savoir les tarses, métatarses et phalanges. Le complexe scapulaire formant l'épaule comprend la scapula et le coracoïde ainsi que deux petits os, le sternum et la clavicule (cette dernière étant fusionnée chez les théropodes pour former un os en forme de 'V' nommé furcula) et le bassin se constitue d'un ilion, d'un ischion et d'un pubis. Les piques, plaques et nodules que portent certains dinosaures sont enfin des ostéodermes, des protubérances osseuses dérivant de l'ossification de certaines parties du derme[45],[46],[27],[47],[48].
Organes internes


Si l'anatomie osseuse des dinosaures est relativement bien connue grâce à la découverte de leurs os fossilisés, il en est tout à fait autrement de leur anatomie interne. En effet, les organes internes que sont par exemple le cerveau, les organes respiratoires et digestifs (poumons, estomacs, intestins, etc.) et les muscles, se dégradent rapidement une fois l'animal mort et seuls des cas exceptionnels de dépôt rapide et de fossilisations permettent à ces parties molles du corps d'être préservées dans la roche.
Un des plus beaux exemples est donné par le théropode juvénile Scipionyx dont les organes et tissus mous ont été particulièrement bien conservés[58]. La préservation unique des ligaments, des cartilages et des muscles du corps, des vaisseaux sanguins et de la totalité des organes digestifs chez ce petit coelurosaure a permis la découverte d'éléments cruciaux sur l'anatomie générale des dinosaures, qui semble se rapprocher beaucoup plus de celle des oiseaux que des reptiles contemporains[59].
Bien qu'à un degré inférieur de préservation, des restes de viscères et de muscles fossilisés d'autres dinosaures ont été mis au jour : Mirischia[60] préservant des traces d'intestins, Santanaraptor[61] et Pelecanimimus[62] montrant des restes de peau et de muscles, ainsi que plusieurs exemples d'hadrosauridés momifiés comme Brachylophosaurus (nommé «Leonardo»)[63] et Edmontosaurus (nommé «Dakota»)[64] chez lesquels on a retrouvé des restes tégumentaires, ligamentaires et viscéraux.
La découverte d'un cœur fossilisé de Thescelosaurus, un petit dinosaure appartenant au sous-ordre des ornithopodes, a fait l'objet d'un article dans Science en avril 2000. Après une étude par tomodensitométrie, il semblerait que le cœur de Thescelosaurus était divisé en quatre chambres (deux aortes et deux ventricules)[pas clair], comme celui des mammifères et des oiseaux[65]. Un examen minutieux de ce cœur pétrifié, qui aurait été conservé par saponification et transformation en goethite selon certains[65], a révélé qu'il s'agit-là plus vraisemblablement d'une concrétion rocheuse[66],[67].
Muscles
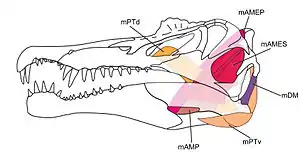
La musculature des dinosaures a été particulièrement étudiée ces dernières années et les reconstitutions musculaires se basent moins sur les tissus musculaires préservés, qui restent très rares dans le registre fossile, mais sur les attaches musculaires visibles sur les os et une comparaison minutieuse avec l'anatomie musculaire des proches parents des dinosaures vivants de nos jours, à savoir les oiseaux et les crocodiles. Ainsi, les muscles du crâne et de la mâchoire, qui comprennent les muscles orbito-temporaux [68], palataux [69], temporaux [70] et mandibulaires [71], ont été récemment reconstitués chez Edmontosaurus, Diplodocus et Majungasaurus[72]. Quant aux muscles du cou, une reconstitution de leur anatomie a pu être donnée chez certains théropodes[73],[74],[75], marginocéphales[76] et sauropodes[77]. De la même manière, des données nouvelles sur la musculature des membres antérieurs et postérieurs chez les dinosaures ont été récemment apportées, par exemple chez Tyrannosaurus[78],[79] et les Dromaeosauridae[80], contribuant à développer de manière importante les connaissances sur la locomotion des dinosaures[81],[82],[83].
Cerveau
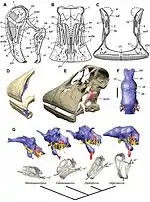
Du fait de sa complexité, la boîte crânienne (ou neurocrâne) renfermant le cerveau a été pendant longtemps une des régions du squelette des dinosaures les moins connues[45]. Néanmoins, les connaissances relativement faibles de son anatomie aux siècles derniers se sont considérablement améliorées au XXIe siècle avec l'avènement du CT-scan. Cet outil utilisé dans les hôpitaux a permis de scanner avec précision et dans les moindres détails la cavité céphalique des boîtes crâniennes relativement bien conservées de nombreux dinosaures. Des reconstructions en 3 dimensions des hémisphères cérébraux, des bulbes olfactifs et des organes de l'oreille interne ont pu être ainsi faites chez plusieurs dinosaures comme Majungasaurus[84], Tyrannosaurus[85], Alioramus[86], Hypacrosaurus[87], Pachyrhinosaurus[88], Diplodocus[89], Camarasaurus[89] et Nigersaurus[90]. Ces études ont permis de comprendre beaucoup mieux l'anatomie du cerveau des dinosaures, mais également d'avoir une idée de leur intelligence et de leurs facultés sensorielles (ouïe, odorat, vue). L'on sait à présent que la morphologie du cerveau est très variable au sein du groupe des dinosaures et les ornithischiens, les sauropodomorphes et les théropodes basaux possédèrent généralement un cerveau proche des reptiles actuels dans sa configuration anatomique, alors que les théropodes dérivés avaient un cerveau ressemblant beaucoup plus à celui des oiseaux. Le cerveau des dinosaures peut se diviser en deux parties, le bulbe olfactif antérieurement et les hémisphères cérébraux au centre, auxquels s'ajoutent, de part et d'autre du cerveau, les 12 nerfs majeurs du cerveau (notés de I à XII), les veines cérébrales et enfin l'oreille interne, constitué du labyrinthe osseux. L'évolution du cerveau chez les théropodes correspond ainsi a un accroissement important du volume cérébral chez les coelurosaures, de même qu'un alignement du bulbe olfactif, des hémisphères cérébraux (télencéphale) et du rhombencéphale sur un même plan[91],[92]. La découverte exceptionnelle d'un cerveau fossilisé qui aurait appartenu à un iguanodonte a permis de constater que la structure de celui était proche des oiseaux modernes et des crocodiliens. En outre, l'étude associée, publiée en 2016, a permis de suggérer que la boîte crânienne et le cerveau chez ces animaux étaient en contact l'un avec l'autre et non pas séparés par un système sanguin. Cette hypothèse indiquerait une taille plus importante du cerveau que celle qui était considérée jusqu'alors[93],[94].
Peaux, poils et plumes

Les dinosaures furent classés pendant longtemps comme des reptiles couverts d'écailles tels nos crocodiles et lézards actuels. Cette vision a fortement changé de nos jours depuis la découverte de structures osseuses typiques des animaux homéothermes (à sang chaud) chez de nombreux dinosaures, et depuis que la province du Liaoning, en Chine, a livré dans les années 1990 et 2000 un grand nombre de fossiles de dinosaures couverts de poils et de plumes. Si certains dinosaures portaient ce genre de téguments filamenteux, d'autres semblent avoir eu la peau lisse, et d'autres encore avoir été couverts d'écailles ; plusieurs types de phanères ont pu coexister, comme chez les oiseaux actuels. En effet, des traces d'écailles ont été découvertes chez de nombreux groupes de dinosaures[95] comme les hadrosauridés (par exemple, Saurolophus[96], Brachylophosaurus[63], Edmontosaurus[64]), les cératopsidés (p. ex. Centrosaurus[97], Chasmosaurus[98]), les stégosauridés (p. ex., Gigantspinosaurus[99], Hesperosaurus[100]), les ankylosauridés (p. ex. Scelidosaurus[101], Tarchia[102]), les sauropodes[103] (p. ex., Saltasaurus[104], Tehuelchesaurus[105]) et les théropodes (p. ex., Carnotaurus[106], Juravenator[107]). Il s'agit dans la plupart des cas d'impressions de peau dans le sédiment mais il existe des cas exceptionnels de peaux fossilisées dont la structure moléculaire a été préservée, comme les momies de dinosaures dont la plupart sont des hadrosaures.


La très grande majorité des paléontologues s'accorde à présent à dire que les dinosaures, et plus exactement les théropodes, furent les ancêtres directs des oiseaux, et les preuves de plumes chez certains théropodes non aviens ont grandement conforté cette théorie. En effet, l'évolution de la plume se suit très bien tout au long du clade des dinosaures, depuis les dinosaures relativement basaux portant un duvet pileux, jusqu'aux théropodes dérivés déployant de véritables rémiges. La présence de poils est avérée chez aux moins deux ornithischiens, l'Heterodontosauridae Tianyulong[108] et le Psittacosauridae Psittacosaurus[109], et un certain nombre de théropodes dont le Megalosauridae Sciurumimus[110], le Tyrannosauroidea Dilong[111], les Compsognathidae Sinosauropteryx[112] et Juravenator[113], et l'Alvarezsauridae Shuvuuia[114]. Ces poils de structures très simples se différencient déjà des téguments filamenteux plus larges et plus longs du Therizinosauroidea Beipiaosaurus[115],[116]. L'Oviraptoridae Caudipteryx[117] et le Dromaeosauridae Sinornithosaurus[118],[119],[120] portent quant à eux des proto-plumes, des structures tégumentaires branchues comprenant une tige centrale à laquelle se joignent des filaments latéraux. Finalement, plusieurs coelurosaures dérivés dont les Oviraptoridae Caudipteryx[117],[121] et Similicaudipteryx[122], les Dromaeosauridae Microraptor[123] et les Troodontidae Anchiornis[124] furent couverts de plumes de type pennes tout à fait homologues à celles de nos oiseaux actuels. Des plumes asymétriques, caractéristiques des oiseaux modernes pouvant voler, ont été découvertes chez Jianianhualong, cette découverte n'impliquant toutefois pas nécessairement la capacité de ce troodontidé au vol[125]. Bien qu'aucun fossile de Velociraptor n'ait conservé des empreintes de plumes, les os des membres antérieurs, et plus spécifiquement l'ulna, montrent de petites protubérances alignées de manière régulière le long de l'os et interprétées comme étant le lieu de fixation de plumes modernes (formée de rachis, vannes et barbules)[126]. Des structures similaires ont également été découvertes le long de l'ulna du Carcharodontosauridae Concavenator, un théropode beaucoup plus primitif que les maniraptoriformes à plumes, démontrant, s'il ne s'agit pas là de lieux d'attachement des muscles[27], la présence de rémiges le long des bras déjà chez ces théropodes relativement primitifs[127]. En 2016 un article publié dans Current Biology annonce la découverte en Birmanie un bloc d'ambre daté de 99 millions d'années contenant un fragment de queue attribué à un coelurosauria et où apparaissent clairement des plumes[128].

Bien que la plupart des dinosaures semble avoir porté uniquement des écailles, il n'est pas impossible que ceux-ci aient perdu la pilosité de leur ancêtres. En effet, les dinosaures à poils les plus primitifs appartiennent au groupe des Heterodontosauridae[108], des ornithischiens primitifs de petite taille, néanmoins la présence de poils est également avérée chez certains ptérosaures comme Sordes, des reptiles volants proches parents des dinosaures. Il n'est pas encore certain que les structures filamenteuses des ptérosaures et des dinosaures soient homologues mais si cela devait être le cas, cela signifierait que les ancêtres communs des ptérosaures et des dinosaures portaient des poils, et donc que les dinosaures les plus basaux furent déjà à fourrure[108],[27].
Couleurs

La découverte des dinosaures à plumes en Chine à la fin du siècle dernier et de ce siècle-ci a révolutionné nos connaissances des dinosaures et de l'évolution de leur descendants actuels les oiseaux. Mais leur découverte a permis d'amorcer une tout autre révolution dans le domaine de la paléontologie des dinosaures, celle de leurs couleurs. Depuis leur naissance au milieu XIXe siècle jusqu'au début du XXIe siècle, il était impossible de fournir des informations concrètes sur la couleur des dinosaures. En effet, les pigments de couleur ne se préservent pas à l'état fossile, et l'entièreté des fossiles de peau, de poils et de plumes des dinosaures ont perdu leur coloration à jamais[129]. Néanmoins, grâce à un examen minutieux des poils et des plumes extrêmement bien préservés des dinosaures du Liaoning à l'aide d'un microscope électronique à balayage (SEM), les paléontologues ont découvert dans les années 2010 la présence de mélanosomes fossilisés, ces organites de cellules pigmentées comprenant la mélanine et responsables, entre autres, des couleurs arborées par les poils et les plumes[130],[131]. La découverte de ces mélanosomes a permis avant tout de démontrer la nature épidermique des plumes des dinosaures[130], qui furent vues par quelques rares paléontologues comme des fibres de collagènes[132],[133],[134].
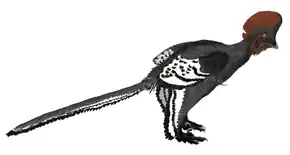
La combinaison de forme, taille, densité et distribution des mélanosomes a également permis de reconstituer les motifs de couleurs déployés par les plumes. Les mélanosomes qui produisent des couleurs noires et grises sont ainsi longs et étroits alors que ceux donnant des couleurs brun-rouge et marron sont courts et larges[131]. Cette technique a jusqu'ici été appliquée à trois dinosaures, au Compsognathidae Sinosauropteryx, au Troodontidae Anchiornis et au Dromaeosauridae Microraptor. Les résultats de cette technique ont ainsi révélé, pour Sinosauropteryx, un pelage de la queue fait d'une alternance de bandes de couleurs claires et foncées, ces dernières déployant des tons plutôt brun-rouge[130]. Chez Anchiornis, le plumage qui couvrait le corps et le cou était généralement grisé. Les ailes des membres antérieurs portaient des bandes de couleurs blanches et noires, alors que les ailes des membres postérieurs étaient blanches à leur base et noires à leur extrémité. Enfin, le plumage de la tête fut généralement grisé et tacheté de couleurs brun-rouge, tout comme les plumes de la crête qui semblent avoir été colorées plus largement de brun-rouge[131]. Quant à Microraptor, la présence de mélanosomes de forme allongée et étroite au niveau des plumes a permis de révéler un plumage de couleur noire majoritairement iridescent[135].

Puisque la couleur des dinosaures fut totalement inconnue jusqu'il y a peu, et reste encore ignorée pour un grand nombre de dinosaures, les artistes de la première moitié du XXe siècle dépeignaient les dinosaures dans des tons de brun, vert et gris, basant leur interprétation sur les animaux actuels comme les reptiles (crocodiles, dragons de Komodo, etc.) et les gros mammifères (éléphants, hippopotames, etc.). La palette des artistes actuels s'est néanmoins fortement élargie. Les dinosaures sont de nos jours représentés avec des couleurs vives, portant un large éventail de couleurs comme beaucoup d'oiseaux, de reptiles et de mammifères actuels, reflétant ainsi leur activité sociale complexe, une idée qui a émergé dans les années 1960[129].
Taille et poids

Si les dinosaures fascinent une grande partie du public, c'est surtout en raison de la taille gigantesque que certains d'entre eux ont atteinte. Une majorité de personnes voient d'ailleurs les dinosaures comme des animaux uniformément grands et imposants, oubliant souvent que beaucoup d'entre eux furent aussi très petits comme le montrent beaucoup de dinosaures à plumes d'une trentaine de centimètres de longueur seulement[136],[137]. Néanmoins, les dinosaures furent les géants de leur époque, surpassant en taille tout autre animal contemporain. Ils restent de nos jours les animaux terrestres les plus grands et les plus lourds à avoir foulé notre planète, tant au niveau des herbivores, dont les sauropodes ont les records de poids et de taille[138],[139], que des carnivores, dont les plus grands dépassaient les 12 mètres de longueur[140].
La taille des dinosaures a fluctué durant le Trias, le Jurassique et le Crétacé[141]; néanmoins les dinosaures ont, dès leur apparition, augmenté de taille tout au long du Mésozoïque[142], et atteint une taille particulièrement grande dès la fin du Trias, en témoigne le prosauropode Plateosaurus dont la taille fut estimée à 8 à 10 mètres pour un poids oscillant de 600 à 900 kilogrammes[143]. Cette augmentation de taille s'observe chez tous les groupes de dinosaures mis à part les sauropodes Macronaria et les théropodes coelurosaures[142]. Ces derniers ont donné naissance aux oiseaux qui furent particulièrement petits au début de leur histoire évolutive. La réduction de taille est ainsi une tendance évolutive rare chez les dinosaures[144]. La distribution de taille des dinosaures est également unique parmi les vertébrés, la courbe de taille des dinosaures étant positionnée au niveau d'animaux de grande taille au contraire des poissons, reptiles, mammifères et oiseaux actuels, démontrant ainsi une stratégie évolutive différente entre dinosaures et vertébrés actuels[145]. Ceci s'observe également entre les dinosaures herbivores (sauropodomorphes et ornithopodes) et les carnivores (théropodes), ces derniers ayant autant d'individus de petite, moyenne et grande taille alors que les dinosaures herbivores comptent surtout des animaux de grande taille. Cette divergence s'explique par le fait que les dinosaures herbivores ont très vite augmenté leur taille afin d'échapper aux prédateurs et de maximiser leur efficacité digestive, au contraire des carnivores qui avaient des ressources en nourriture suffisantes parmi les dinosaures juvéniles et d'autres proies de petites tailles[145].
En 1990, avant la découverte d'un grand nombre de très petits dinosaures à plumes, une étude portant sur 63 genres de dinosaures évalua le poids moyen d'un dinosaure à 850 kg, un poids comparable à celui d'un grizzly, et un poids médian (qui devait osciller entre 9 kg et 5 tonnes) de près de deux tonnes, alors que le poids moyen des mammifères est de 863 grammes, soit celui d'un gros rongeur[146].
Plus grands et plus petits dinosaures
Les paléontologues ont de tous temps essayé d'estimer la taille et le poids des dinosaures, particulièrement des plus grands comme Brachiosaurus et des plus féroces tel Tyrannosaurus. La première méthode quantitative sérieuse fut proposée par Gregory en 1905 afin d'évaluer le poids d'Apatosaurus (alors nommé Brontosaurus)[147]. Sur base d'un modèle rigoureux de l'animal en argile et en plastique sculpté à la main, il put extrapoler le volume de l'animal en l'immergeant dans un liquide et en mesurant le volume de liquide déplacé, puis en le multipliant par la densité présumée du dinosaure (à savoir celle de l'eau) et par le facteur d'échelle du modèle. Cette technique fut suivie par d'autres auteurs jusque dans les années 1980 avant l'avènement de techniques informatiques plus robustes. Colbert, en 1962, estima ainsi le poids de Tyrannosaurus à 7 tonnes, 8,5 tonnes pour Triceratops, 28 tonnes pour Apatosaurus et 78 tonnes pour Brachiosaurus[148].
Seule une petite fraction des animaux morts se fossilisent et seuls quelques spécimens découverts sont des fossiles complets ; les impressions de peaux et de tissus mous sont rares.
La reconstruction d'un squelette d'une espèce en comparant la taille et la morphologie des os avec ceux d'une autre espèce similaire mieux connue est un art inexact ; faire la recomposition des muscles et des autres organes d'un spécimen est scientifiquement difficile. On ne sera donc jamais vraiment certain de la taille des plus grands et plus petits dinosaures.
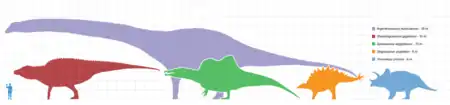
Parmi les dinosaures, les sauropodes étaient gigantesques, les plus grands étaient d'un ordre de grandeur plus massif que tous les animaux ayant marché depuis sur la Terre. Des mammifères préhistoriques comme l'Indricotherium et le mammouth colombien étaient des nains comparés aux sauropodes. Seule une poignée d'animaux aquatiques contemporains les approchent ou les surpassent en taille, telle la baleine bleue, qui pèse 180 tonnes et atteint 31 mètres de long au maximum.
Le plus grand et plus lourd dinosaure connu à partir de squelettes complets ou presque est le Giraffatitan (autrefois connu sous le nom de Brachiosaurus brancai)[149]. Il mesurait 12 m de haut, 22,5 m de long, et aurait pesé entre 30 et 60 tonnes (pour mémoire un éléphant de savane d'Afrique, le plus grand animal terrestre du monde, pèse en moyenne 7,7 tonnes). Le plus long dinosaure issu d'un fossile complet est le Diplodocus qui faisait 27 m (Pittsburgh, musée Carnegie d'histoire naturelle, 1907).
Il y avait de plus grands dinosaures mais les données connues sont estimées sur quelques fossiles fragmentaires. La plupart sont des herbivores découverts dans les années 1970 ou après, parmi lesquels l'énorme Argentinosaurus, qui pourrait avoir pesé entre 80 et 100 tonnes ; le plus long de tous, le Supersaurus de 40 mètres ; et le plus grand, le Sauroposeidon de 18 mètres, qui aurait pu atteindre une fenêtre au 6e étage.
Un dinosaure encore plus grand, le Amphicoelias fragillimus, connu seulement de quelques vertèbres découvertes en 1878, pourrait avoir atteint 58 mètres de long et un poids de 120 tonnes[150]. Le plus lourd aurait pu être le peu connu et encore débattu Bruhathkayosaurus, qui pourrait avoir atteint de 175 à 220 tonnes. On admet actuellement que Bruhathkayosaurus matleyi ne devait pas dépasser 139 tonnes, pour environ 34 mètres de long.
Le plus grand carnivore était le Spinosaurus, qui atteignait une taille de 16 à 18 mètres et pesait 9 tonnes[151]. D'autres grands carnivores incluaient les Giganotosaurus, Mapusaurus, Tyrannosaurus rex et Carcharodontosaurus.
Sans inclure les oiseaux contemporains comme les oiseaux-mouches, les plus petits dinosaures avaient la taille d'un corbeau ou d'un poulet. Les théropodes Microraptor et Parvicursor faisaient moins de 60 cm de long.
Biologie
Comportements

L'interprétation du comportement des dinosaures non aviens est généralement établie sur la disposition des fossiles découverts, leur habitat, les simulations par ordinateur de leurs biomécaniques (basées sur les pistes fossiles et sur des comparaisons avec des animaux actuels situés dans la même niche écologique, ainsi que l'étude des représentants actuels des dinosaures : les oiseaux). On a aussi insufflé de l'air dans les moulages de crânes de certains hadrosaures à crête pour entendre les sons que cette expérience produisit (sans que cela donne la moindre indication sur leurs rythmes et modulations). Comme telle, la compréhension actuelle du comportement des dinosaures fossiles repose donc en grande partie, comme leurs coloris, sur des spéculations, dont certaines resteront probablement controversées pendant encore longtemps. Cependant, il y a un consensus sur le fait que certaines caractéristiques qui sont communes chez les oiseaux et les crocodiles (le groupe le plus proche des dinosaures), telle que la nidification, aient été courantes chez l'ensemble des dinosaures disparus (et ce d'autant plus que des nids fossiles ont été découverts en nombre : voir plus bas).
Troupeaux
La première preuve de l'existence de troupeaux de dinosaures fut découverte en 1878 en Belgique à Bernissart. 31 Iguanodons semblaient avoir péri ensemble après être tombés dans une doline profonde et inondée[152] ; au vu des dernières analyses, ces squelettes semblent s'être déposés lors de trois événements distincts[153].
D'autres sites de morts massives furent découverts. Ceux-ci, avec de nombreuses traces fossiles suggérèrent que les troupeaux ou les hordes étaient communes dans beaucoup d'espèces. Les pistes de centaines, voire de milliers d'herbivores, indiquent que les dinosaures à bec de canard pouvaient se déplacer en grands troupeaux, tel le bison ou le springbok. Des traces de sauropodes permirent de voir que ces animaux voyageaient en groupes composés de plusieurs espèces différentes[154], et d'autres gardaient les jeunes au milieu du troupeau afin de les protéger, d'après les traces au Davenport Ranch au Texas.
Soins parentaux
La découverte en 1978 par Jack Horner du nid du Maiasaura (« dinosaure bonne mère ») au Montana démontra que les soins parentaux duraient bien après l'éclosion chez les ornithopodes[155]. Il y a aussi des preuves que d'autres dinosaures du Crétacé comme le sauropode Saltasaurus (découvert en 1997 en Patagonie) avaient des comportements similaires, et que ces animaux se regroupaient en immenses colonies nidificatrices comme celles des manchots. Ce comportement de nidification groupée et de protection des œufs est confirmé en 2019, avec la découverte en Mongolie de 15 nids (et plus de 50 œufs, dont environ 60 % d'œufs éclos) vieux d'environ 80 Ma et clairement issus de la même saison de nidification[156],[157].
L'Oviraptor de Mongolie a été découvert (1993) dans une position de couvaison comme celle de la poule, ce qui signifie qu'il était recouvert d'une couche de plumes isolantes qui gardait l'œuf au chaud[158]. Des pistes fossiles ont aussi confirmé le comportement maternel parmi les sauropodes et les ornithopodes de l'île de Skye[159]. Des nids et des œufs fossilisés ont été trouvés pour la plupart des principaux groupes de dinosaures, et il apparaît probable que les dinosaures non aviens communiquaient avec leurs petits d'une manière similaire aux dinosaures actuels (oiseaux) et aux crocodiles.
Certains dinosaures installaient leurs nids sur des sites hydrothermaux, ce qui permettait aux œufs de bénéficier d'une température idéale et régulière[160].
Une étude de 2017 basée sur les lignes de croissance incrémentielle de dents fossiles de certains dinosaures ornithischiens (Protoceratops andrewsi et Hypacrosaurus stebingeri) a conclu que l'incubation des œufs était longue (3 à 6 mois avant l'éclosion), au moins chez plusieurs espèces[161]. Ce long délai pourrait avoir contribué à la disparition des dinosaures non aviens face aux oiseaux et mammifères après l'évènement du Crétacé-Paléogène, la question restant de savoir si les théropodes avaient la même physiologie, notamment ceux proches des oiseaux tel Velociraptor[162].
Accouplement et communication
Les crêtes de certains dinosaures, comme les marginocéphales, les théropodes et les hadrosauridae, pourraient avoir été trop fragiles pour une défense active et donc auraient probablement été utilisées pour les parades sexuelles ou à des fins d'intimidation, bien qu'il existe peu d'éléments sur le territorialisme et l'accouplement des dinosaures.

La nature des communications entre dinosaures reste également énigmatique, mais des découvertes récentes suggèrent que la crête creuse des lambeosaurines pourrait avoir fonctionné comme une caisse de résonance utilisée pour une grande variété de vocalisations.
Alimentation
La morphologie et la dentition des dinosaures permettent de distinguer les herbivores des carnivores (ces derniers, munis de dents comprimées en forme de lame, pouvaient être charognards, chasseurs, piscivores ou insectivores) et des omnivores[163]. Les gisements de fossiles montrent souvent une surabondance des carnivores.
Les coprolithes permettent de préciser le régime alimentaire, de même que l'analyse isotopique (42Ca/44Ca et 13C/12C) des dents, qui peut indiquer quels types de plantes étaient consommés[164]. Par exemple, vers 120 et 100 Ma BP en Afrique du Nord, les abélisauridés et les carcharodontosauridés (des théropodes) chassaient préférentiellement des proies terrestres (dont des dinosaures herbivores), alors que les spinosaures étaient piscivores (tandis que les crocodiles géants comme Sarcosuchus — qui n'étaient pas des dinosaures — avaient un régime mixte)[165],[166]. Certains dinosaures herbivores comme Argentinosaurus ingurgitaient avec la végétation des cailloux, gastrolithes qu'ils gardaient dans leur estomac pour faciliter la digestion[167].
Chasse
D'un point de vue comportemental, l'un des fossiles les plus importants de dinosaure a été découvert dans le désert de Gobi en 1971. Il incluait un Velociraptor attaquant un Protoceratops[168], prouvant physiquement que les dinosaures s'attaquaient et se mangeaient entre eux.
Cannibalisme
Bien que le cannibalisme parmi les théropodes ne constitue nullement une surprise[169], il a été confirmé par des traces de dents sur un fossile de Majungasaurus à Madagascar en 2003[170].
Déplacement
Basé sur les preuves fossiles existantes, il n'y avait aucune espèce de dinosaure fouisseur et peu de dinosaures grimpeurs. Puisque l'expansion des mammifères au cénozoïque vit l'apparition de nombreuses espèces fouisseuses et grimpantes, le manque de preuves pour des espèces de dinosaures similaires est quelque peu surprenant.
Une bonne compréhension de la façon dont les dinosaures se déplaçaient est la clef des modèles de comportements des espèces. La biomécanique en particulier a fourni de nombreux éléments comme la détermination de la vitesse de course des dinosaures d'après l'étude des forces exercées par leurs muscles et la gravité sur la structure de leur squelette[171],[172], savoir si les diplodocides pouvaient créer un bang supersonique en balayant l'air avec leur queue en forme de fouet[173], déterminer si les théropodes géants devaient ralentir quand ils poursuivaient leurs proies pour éviter des blessures mortelles[174], et si les sauropodes pouvaient flotter[175].
Cycle d'activité
La théorie scientifique qui prévalait jusqu'ici est que les dinosaures avaient des modes de vie diurnes, alors que les premiers mammifères apparus étaient crépusculaires ou nocturnes[176],[177].
Une étude paléontologique en 2011 de la structure oculaire (longueur de l’orbite et diamètre de l’anneau scléreux) sur 33 espèces d'archosauriens, comparée à celle de 164 espèces actuelles (dont des reptiles, des oiseaux et des mammifères), montre que les dinosaures carnivores étaient partiellement nocturnes, les dinosaures volants et ptérosaures principalement diurnes et les dinosaures herbivores surtout cathéméraux (actifs de jour comme de nuit)[178].
Métabolisme
Une étude française sur la composition isotopique en oxygène des dents et os de 80 dinosaures du Crétacé (théropodes, sauropodes, ornithopodes et cératopsiens[179]) provenant de gisements d'Amérique du Nord, d'Europe, d'Afrique et d'Asie, a montré que ceux-ci devaient être homéothermes. Le rapport 18O/16O — qui dépend de la température interne de l'animal vivant — est identique à celui des mammifères et oiseaux, homéothermes, et diffère nettement de celui des reptiles actuels, ectothermes, et des chéloniens et crocodiliens fossiles du Crétacé.
La présence de structures de Havers (micro-canaux entourés d'une couche d'os concentrique au sein des squelettes) dans les os fossilisés serait également un élément en faveur du caractère endotherme[180].
En 2006, une étude a estimé que la température était proportionnelle à la masse et au taux de croissance, allant de 25 °C pour les petits dinosaures jusqu’à 41 °C pour les plus grands[181]. Un modèle numérique, permettant d'estimer la température corporelle en fonction de la taille et du rythme de croissance, a été appliqué à huit espèces, du psittacosaure (Psittacosaurus mongoliensis, 12 kg) à l'apatosaure (Apatosaurus excelsus, 26 000 kg). D'après cette étude, la température interne de Sauroposeidon proteles, le plus lourd des dinosaures connus (60 tonnes), devait atteindre 48 °C. Ce modèle tendant à prouver que les gros dinosaures étaient chauffés par « homéothermie inertielle », a été remis en cause par R. Eagle et ses collègues. Ces derniers ont en effet estimé la température corporelle de grands sauropodes du Jurassique, tels Camarasaurus et Brachiosaurus, entre 36 et 38 °C, équivalente à celles de mammifères actuels[182].
Cependant, en 2014 une nouvelle étude comparative sur plus de 400 espèces éteintes et vivantes conclut que les dinosaures devaient avoir le sang tiède, dit mésotherme[183]. Une étude en 2015 basée sur la température interne corrélée à la composition chimique des œufs fossilisés estime la température des sauropodes à environ 38 °C contre 32 °C pour celle des oviraptoridés[184].
Histoire évolutive
Phylogénie au sein des archosaures
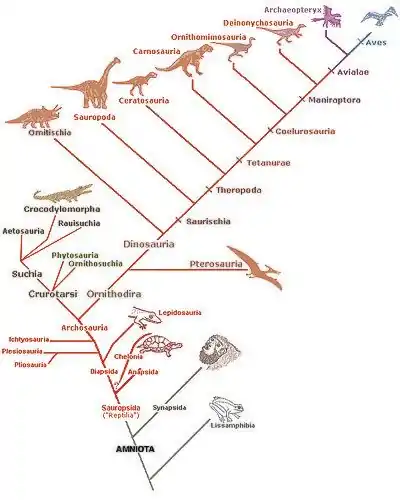
Parmi les amniotes (eux-mêmes issus des reptiliomorphes), les dinosaures sont apparus au sein du super-ordre des archosauriens, un groupe de petits diapsides de la fin du Permien et surtout du début du Trias, qualifiés de reptiles.
L'extinction radicale de la fin du Permien — l'extinction Permien-Trias, il y a environ 251 millions d'années, a balayé de 80 à 96 % des espèces marines, et 70 % des familles de vertébrés terrestres de l'époque[185],[186], permettant à des groupes d'animaux ou de plantes de prendre leur essor par radiations évolutives, notamment les amniotes, parmi lesquels on trouve les reptiles, les ancêtres des dinosaures, et ceux des mammifères anciens et actuels. De nombreux évènements géologiques sont contemporains de cette crise.
Les petits diapsides, c'est-à-dire dont le crâne a deux paires de fosses en arrière des orbites, possèdent des os pré-dentaires à l'avant de la mandibule et des membres transversaux ou semi-dressés. Ils partent à la conquête des écosystèmes et ils divergent en trois grandes familles :
- les ptérosaures qualifiés de reptiles volants ;
- les crocodiliens, dont la filiation actuelle s'est perpétuée dans un milieu amphibie ;
- les dinosaures, qui, aptes à la mobilité terrestre, ont des pattes dressées dans un plan parallèle au plan sagittal de leur corps. Leurs postures redressées sont le terme d'une rapide évolution similaire à celles des mammifères, et très différentes de celle des reptiles.
L'analyse de la structure du bassin, comportant trois os pairs, ilion, pubis, ischion permet de retrouver ces différences. Ainsi le paléontologue Harry Govier Seeley a fondé dès 1887[187],[188] la distinction capitale entre :
- les saurischiens, littéralement « à bassin de reptiles ». Les os du bassin ont une configuration triradiée. Les oiseaux sont issus de ce groupe.
- les ornithischiens, « à bassin d'oiseaux ». La ressemblance invoquée reste ici assez superficielle, mais il existe des conformations, où le pubis est parallèle à l'ischion vers le bas et l'arrière. Les oiseaux ne descendent pas de ce groupe, contrairement à ce que laisse croire ce nom.
Cladogramme de la branche Avemetatarsalia d'après Nesbitt (2011) et Nesbitt et al. (2017) :
| Archosauria |
| |||||||||||||||||||||||||||||||||||||||||||||
Premiers dinosaures
Les premiers dinosaures sont apparus au début du Trias il y a environ 245 millions d'années, ou selon les plus récentes découvertes, il y a 250 millions d'années[189] ; ils étaient peu nombreux cependant, avant le Trias supérieur il y a environ 232 millions d'années, moment où débute l'expansion qui leur confèrera une domination écologique[190].
Les traces de pas fossiles les plus anciennes, et attribuées à des dinosaures, datent de 248 millions d'années environ et concernent « de petits reptiles quadrupèdes, de la taille d’un chat, [qui] ont laissé leurs empreintes dans l’argile il y a environ 250 millions d’années […]. Ces pas ont été découverts dans le sud-ouest [de la] Pologne, dans les Monts Sainte-Croix. […] Les empreintes […] ont été laissées il y a 248 millions d‘années par des reptiles du genre Prorotodactylus, quadrupèdes dotés de pattes à cinq doigts, dont les trois doigts centraux sont les plus fortement imprimés dans le sol. Proches les unes des autres, ces traces se distinguent de celles d’autres reptiles du groupe des crocodiles et des lézards[191] ».
Des « traces […] plus récentes découvertes par l’équipe de Grzegorz Niedzwiedzki de l’université de Varsovie datent de 246 millions d’années. Plus grosses – une quinzaine de centimètres – elles auraient été laissées par des dinosaures du genre Sphingopus, bipèdes[191] ».
Ces traces font probablement remonter la toute première espèce de dinosaures encore indifférenciée entre ornithischiens et saurischiens quelques millions d'années plus tôt, soit dès la fin du permien.
Le plus ancien dinosaure connu par des fossiles est Nyasasaurus âgé de 243 millions d'années, découvert en Tanzanie[192]. Ensuite viennent de fossiles de carnivores âgés de 225 à 230 millions d'années, Eoraptor et Herrerasaurus, découverts dans la Formation d'Ischigualasto en Argentine[20],[193]. Tous deux montrent déjà une certaine spécialisation, puisque ce sont des saurischiens, datant d'après la divergence entre les dinosaures ornithischiens et saurischiens. Notons que tous les dinosaures ont une denture homodonte comme les autres reptiles. Herrerasaurus est non seulement un saurischien, mais peut-être même un théropode déjà affirmé[194].
Expansion des dinosaures
Les dinosaures ont commencé à connaître une diversification rapide dès le Carnien (Trias supérieur), il y a environ 232 millions d'années, et sont sortis triomphants écologiquement du Trias supérieur, après avoir supplanté les rhynchosaures en tant qu'herbivores, et les archosaures basaux en tant que carnivores[190]. Plusieurs causes ont été avancées pour expliquer cette explosion radiative. Selon une première opinion, les dinosaures étaient plus compétitifs que d'autres espèces, du fait de leur posture droite, de leur habileté prédatrice et de leur sang chaud[190]. Selon un autre modèle explicatif, dit "opportuniste", les dinosaures auraient bénéficié d'un changement floral majeur, au cours duquel les flores de conifères ont remplacé les flores dominées par la fougère dicroidium (en), changement qui aurait entraîné l'extinction des herbivores auparavant dominants[190]. Enfin selon l'explication la plus récente (2018), un changement climatique mondial, associé à l'épisode pluvial du Carnien, avec passage d'un climat sec à un climat humide, et retour au climat sec, dans un intervalle court, a favorisé la diversification rapide des dinosaures[190],[195].
Extinction de la plupart des espèces
La disparition de la plupart des espèces des dinosaures donna naissance à de nombreuses théories, certaines farfelues comme la destruction des dinosaures par des extraterrestres, et d'autres plus probables et scientifiquement testables. Il convient toutefois de noter que l'extinction des dinosaures est un problème sémantique : les dinosaures ne sont pas éteints puisqu'il subsiste des oiseaux.
En revanche, il y a bien eu une crise à la fin du Crétacé, il y a 65 millions d'années. Bien que cette dernière ait eu un impact moyen sur la biodiversité en général (si on la compare à la crise Permo-Triasique ou même à celle de l'Ordovicien) et qu'elle ait surtout décimé des organismes marins tels les foraminifères et non des organismes terrestres dont l'extinction relative est beaucoup moins élevée, elle est devenue très célèbre en raison de la surmédiatisation relative aux dinosaures.
La comparaison du taux d'extinction, taxon par taxon, montre que certains des clades contemporains des dinosaures ont été très affectés (tels les plésiosaures et les ptérosaures) et d'autres beaucoup moins (crocodiliens et chéloniens par exemple). Cette crise a éliminé les dinosaures non aviens vivant à cette époque.
Les causes les plus probables ayant induit la crise K-T sont :
- la chute d'un astéroïde ou d'une comète d'une dizaine de kilomètres de diamètre provoquant une catastrophe majeure qui, par le biais de débris dus à la collision, plongea la Terre dans l'obscurité et le froid pendant plusieurs années, empêchant ainsi la photosynthèse, ce qui induisit un appauvrissement massif en plantes et surtout en plancton et conduisit à l'extinction de nombreuses espèces dépendant de ces ressources quel que soit leur niveau trophique. Cette théorie, de nos jours très argumentée (cratère de Chicxulub au Mexique daté de –65 Ma), permet notamment d'expliquer dans une certaine mesure les survies différentielles des taxons. De plus, à la suite de découvertes récentes (avril 2008) de cénosphères (sphères microscopiques se formant à la suite de la combustion de charbon et de pétrole brut) dans les sédiments de la crise K-T, les chercheurs pensent que la météorite aurait pu s'écraser dans une réserve pétrolière, et projeter dans l’atmosphère de grandes quantités d’hydrocarbures enflammés expliquant la disparition de la majorité des espèces[196]. Une étude publiée le dans la revue Science et s'appuyant sur des techniques de datation radiométriques de haute précision, indique que la météorite se serait écrasée sur Terre il y a 66 038 000 d'années, soit au maximum 32 000 ans avant l'extinction des dinosaures, ce qui renforcerait d'autant la probabilité d'un lien de causalité entre les deux évènements[197] ;
- des éruptions à la fin du Crétacé d'un "supervolcan", lié à la présence d'un point chaud placé actuellement sous l'île de La Réunion et qui forma les trapps du Deccan, en Inde pendant une période assez courte au niveau des temps géologiques (au moins 1 million d'années), provoquant un cataclysme planétaire en modifiant le climat et réduisant la photosynthèse en plongeant la Terre dans une pénombre par le biais de nuages de cendres et de brume sulfureuse. Cette théorie est aussi argumentée que la théorie précédente, entre autres par la datation de –65 Ma des trapps du Deccan, dont la superficie équivaut à plusieurs fois la France, et par des résultats sur la baisse de la luminosité à la surface de la Terre lors de l'éruption récente du Pinatubo () aux Philippines. Cette théorie recoupe en partie celle de la collision météorique sur les effets atmosphériques ;
- des régressions marines très importantes à la fin du Crétacé changeant la conformation et l'étendue des milieux littoraux et benthiques tout en induisant un important changement climatique. Cette théorie est elle aussi basée sur des preuves solides et on sait maintenant qu'une très grande régression marine a eu lieu au crétacé supérieur.
Ces trois théories sont basées sur des faits et la crise K-T pourrait être la conséquence de la quasi-simultanéité de ces trois événements. Les avis divergent en ce qui concerne l'importance relative de chacun d'eux :
- dans une étude publiée le dans le Proceedings of the National Academy of Sciences (PNAS), plusieurs paléontologues de l'Université de Reading ont révélé que 40 millions d'années avant leur extinction, les dinosaures avaient déjà commencé à disparaître[198]; l'hypothèse à privilégier serait alors celle d'une disparition progressive provoquée par la séparation des continents et une forte activité volcanique.
Représentants actuels

Le premier fossile d'oiseau, l'Archaeopteryx, du Jurassique supérieur, a été découvert en Bavière en 1861. Sa grande ressemblance avec certains petits dinosaures carnivores bipèdes, comme les Compsognathus, a immédiatement fait apparaître la théorie selon laquelle les oiseaux descendraient d'un groupe de dinosaures au sein des Cœlurosauriens.
Pendant un siècle, cette théorie est restée très controversée, voire rejetée. En effet, les oiseaux ont des clavicules, alors que les cœlurosauriens n'en avaient pas.
Depuis les années 1970, cependant, des Cœlurosauriens dotés de clavicules ont été découverts (et même dans des groupes moins dérivés) et la théorie dinosaurienne sur l'origine des oiseaux est redevenue dominante.
Dans les années 1990, de nombreux fossiles de dinosaures à plumes ont été découverts, principalement dans la région du Liaoning, en Chine et ont contribué à réactiver cette théorie. Il s'agit dans certains cas d'oiseaux primitifs, et dans d'autres de dinosaures non aviens à plumes ou proto-plumes.
L'interprétation qui est faite de ces découvertes est qu'une ou plusieurs espèces de dinosaure cœlurosaurien (voire l'ancêtre des Cœlurosauriens lui-même) a développé le caractère « plume », semble-t-il sans l'utiliser pour le vol, et a donné naissance à des espèces assez nombreuses de dinosaures à plumes. Une de ces espèces de dinosaures à plume aurait, des millions d'années plus tard, donné naissance à l'ancêtre commun à tous les oiseaux.
En 2011, au Canada, onze plumes de dinosaures, colorées (du marron au noir) ont été retrouvées dans de l'ambre, datant de 78 à 79 millions d'années (fin du Crétacé) avec barbes et barbules visibles, semblables à ceux d'oiseaux contemporains, avec des indices d'imperméabilité pouvant laisser penser qu'il s'agissait (dans ce cas) d'animaux semi-aquatiques[199]. Divers indices laissent penser que les plumes de dinosaures étaient souvent colorées[200].
Bien que cette théorie faisant des oiseaux un sous-groupe des dinosaures soit très largement dominante, il reste des contestataires, regroupés sous l'acronyme « BAND » : Birds Are Not Dinosaurs[201].
Reptiles contemporains des dinosaures
De nombreuses lignées qualifiées de reptiliennes vivaient aux mêmes périodes géologiques que les dinosaures, et ont parfois été confondues avec les dinosaures par le cinéma ou la littérature. Les plus connues sont :
- les ptérosaures (qualifiés de reptiles volants) incluant le ptéranodon et le ptérodactyle ;
- les nothosaures, plésiosaures, les pliosaures et les ichthyosaures (qualifiés de reptiles marins), ainsi que les mosasaures (également qualifiés de reptiles marins puisque ce sont des lépidosaures, comme les actuels varans).
Aucune de ces lignées n'est scientifiquement classée comme dinosaurienne.
Classification
Il n'y a pas encore si longtemps[Quand ?], on classait dans les dinosaures : les ichthyosaures, les pliosaures, les plésiosaures (trois groupes de reptiles aquatiques) et les ptérosaures (reptiles volants)[réf. souhaitée] qui, aujourd'hui, sont considérés comme des lignées évolutives indépendantes des dinosaures parmi les reptiles ayant vécu au Mésozoïque. Les dinosaures étaient un groupe d'animaux extrêmement divers ; selon une étude de 2006, 527 genres de dinosaures ont été décrits avec certitude, et plus de 1 844 genres sont encore à classer[202],[203]. Certains d'entre eux étaient herbivores, d'autres carnivores. Certains dinosaures étaient bipèdes, d'autres étaient quadrupèdes et certains, tels Ammosaurus et Iguanodon, pouvaient marcher aussi bien sur deux ou quatre pattes.
Les dinosaures sont nommés de façon précise selon leur genre et leur espèce. Souvent, ce nom d'espèce est donné en fonction du nom du lieu de sa découverte comme Iguanodo bernissartensis du nom de la localité de Bernissart, en Belgique, où fut découvert un troupeau de 31 iguanodons, ou bien Saltasaurus, le dinosaure de la rivière Salta en Argentine. Une particularité anatomique peut servir de base pour baptiser une espèce, comme c'est le cas pour le Triceratops, la face à trois cornes. Ou c'est le nom d'un paléontologue connu qui est utilisé, comme pour Othnielia du nom d'Othniel Charles Marsh. Avec l'essor de la paléontologie, les noms d'espèces s'internationalisent. Les racines grecques et latines sont remplacées parfois par des racines chinoises, mongoles ou africaines comme Nqwebasaurus venant du xhosa, une langue d'Afrique du Sud[204].
Tableau récapitulatif
Sur la base du code international de nomenclature zoologique, on peut reconnaitre de 630 à 650 genres de dinosaures, rassemblant plusieurs milliers d'espèces de dinosaures (une trentaine de nouvelles espèces sont décrites tous les ans). Un peu plus de la moitié de ces espèces ne sont représentées que par un seul spécimen (souvent incomplet) et moins de 20 % des espèces sont connues par plus de cinq spécimens. Ce tableau général fournit des liens entre les principaux groupes de dinosaures. On trouvera à droite quelques genres illustrant les ordres et les familles représentés.




| Ordre des Saurischia | Ordre des Ornithischia |
|---|---|
|
Famille des Herrerasauridae Sous-ordre des Theropoda
Sous-ordre des Sauropodomorpha
|
Famille des Heterodontosauridae Sous-ordre des Thyreophora
Neornithischia basaux Sous-ordre des Cerapoda
Infra-ordre des Ornithopoda
|
─o Dinosauria │ ├─o Ornithischia │ ├─o Lesothosaurus │ └─o Genasauria │ │ │ ├─o Thyreophora │ │ ├─o Scutellosaurus │ │ └─o │ │ ├─o Scelidosaurus │ │ └─o Eurypoda │ │ ├─o Stegosauria │ │ └─o Ankylosauria │ │ │ └─o Neornithischia │ ├─o Marginocephalia │ │ ├─o Pachycephalosauria │ │ └─o Ceratopsia │ └─o Ornithopoda │ └─o Saurischia │ ├─o Sauropodomorpha │ ├─o Prosauropoda (paraphylétique) │ └─o Sauropoda │ └─o Theropoda ├─o Herrerasauridae └─o Neotheropoda └─o Coelophysoidea └─o Averostra ├─o Ceratosauria │ ├─o Ceratosauridae │ └─o Abelisauroidea │ ├─o Noasauridae │ └─o Abelisauridae └─o Tetanurae ├─o Megalosauroidea │ ├─o Piatnitzkysauridae │ └─o Megalosauria │ ├─o Megalosauridae │ └─o Spinosauridae └─o Avetheropoda ├─o Allosauroidea │ ├─o Metriacanthosauridae │ └─o Allosauria │ ├─o Allosauridae │ └─o Carcharodontosauria │ ├─o Neovenatoridae │ └─o Carcharodontosauridae └─o Coelurosauria ├─o Tyrannosauroidea │ ├─o Proceratosauridae │ └─o │ ├─o Megaraptora │ └─o Tyrannosauridae └─o ├─o Compsognathidae └─o Maniraptoriformes ├─o Ornithomimosauria │ ├─o Deinocheiridae │ └─o Ornithomimidae └─o Maniraptora ├─o Alvarezsauroidea │ └─o Alvarezsauridae └─o ├─o Therizinosauria │ └─o Therzinosauroidea │ └─o Therizinosauridae └─o Pennaraptora ├─o Oviraptorosauria │ ├─o Caudipteridae │ └─o Caenagnathoidea │ ├─o Caenagnathidae │ └─o Oviraptoridae └─o Paraves ├─o Dromaeosauridae │ ├─o Unenlagiinae │ └─o │ ├─o Microraptorinae │ └─o Eudromaeosauria │ ├─o Dromaeosaurinae │ └─o Velociraptorinae └─o ├─o Troodontidae └─o Avialae └─o Aves
Parc à dinosaures
De très nombreux parcs à dinosaures existent dans le monde. Voici une liste non exhaustive des parcs à thème dans le monde :
- Allemagne :
- Dino Park
- Chine :
- États-Unis :
- Dinosaur Provincial Park
- Dinosaur Valley State Park
- France
- Dinopedia Parc
- Italie :
- Parco della Preistoria
- Pologne :
- Turquie :
- Jurassic Land
Musées
De très nombreux musées accueillent des squelettes ou des reconstitutions de dinosaures. Voici une liste non exhaustive des musées les plus importants pour quelques pays du globe :
- Allemagne :
- Belgique :
- Muséum des sciences naturelles (Bruxelles)
- Musée de l'Iguanodon de Bernissart
- Canada :
- Musée royal Tyrrell de paléontologie (Drumheller, Alberta)
- États-Unis :
- France :
- Muséum national d'histoire naturelle (Paris)
- Musée des Confluences (Lyon)
- Muséum départemental du Var (Toulon)
- Muséum de Toulouse
- Muséum d'histoire naturelle d'Aix-en-Provence
- Musée vert, le musée d'histoire naturelle du Mans
- Dinosauria (à Espéraza, dans l'Aude)
- Musée du Parc de la Plaine des Dinosaures de Mèze (Hérault)
- Musée de l'ACAP de Cruzy (Hérault)
- Royaume-Uni
- Suisse
- Muséum d'histoire naturelle de Genève (Genève) — jusqu'au milieu des années 2010

Dans la culture
Arts et littérature
- Gertie le dinosaure est le premier personnage de dessin animé doté d’une personnalité propre, créé en 1904 par le dessinateur Winsor McCay.
- Le Monde perdu, un livre de sir Arthur Conan Doyle publié en 1912.
- Brontosaurus gigantomir roman fantastique de Félicie de Baillehache, dans le supplément littéraire du Figaro du 20 novembre 1921
- Tumak, fils de la jungle (1940) et son remake, Un million d'années avant J.C. (1966), films mettant en scène, sans grand souci de cohérence historique, des hommes préhistoriques confrontés à des dinosaures.
- Casimir : personnage principal de L'Île aux enfants, une émission de télévision pour la jeunesse dans les années 1970 en France.
- Denver, le dernier dinosaure est une série télévisée d'animation franco-américaine mettant en scène un dinosaure né de nos jours.
- Le Petit Dinosaure est une série de dessins animés produits par Steven Spielberg et George Lucas.
- Nabuchodinosaure est le dinosaure improbable et humoristique de la bande-dessinée du même nom.
- Jurassic Park et Le Monde perdu : romans de Michael Crichton puis films de Steven Spielberg, mettant en scène des dinosaures, de façon assez réaliste. Les deux volets ont connu plusieurs suites : Jurassic Park 3 en 2001 ensuite Jurassic World en 2015, Jurassic World: Fallen Kingdom en 2018 et enfin Jurassic World: Dominion en 2022.
- Sur la terre des dinosaures (Walking with dinosaurs) : série en six épisodes de Tim Haines, faisant largement appel aux images de synthèse, produite par la BBC en 1999, souvent rediffusée par la chaîne Planète. Ils apparaissent également en caméo au début de la suite "Sur la Terre des Monstres Disparus".
- Dinosaure est un film des studios Disney qui mêle créations en images de synthèse et décors réels.
- Dinotopia : livres illustrés pour enfants puis nouvelles pour adolescents, romans pour adultes, mini-série en trois épisodes et enfin série télévisée, dont la production a été annulée en cours de saison.
- Kyōryū Sentai Zyuranger : série de sentai dont le premier mot du titre original signifie dinosaure en japonais.
- Gon : bébé tyrannosaure, héros d'un manga de Masashi Tanaka.
- Nick Cutter et les Portes du temps : série comportant cinq saisons mettant en scène des dinosaures venant de portails spatio-temporels.
- Les Dinosaures en bande dessinée : bande dessinée visant à faire découvrir le monde des dinosaures sur un ton humoristique.
- Dinosaur King : série japonaise mettant en scène des enfants avec des dinosaures invoqués par cartes.
- Terra Nova : série américaine mettant en scène des individus venant vivre dans le crétacé supérieur.
- Le Voyage d'Arlo, film d'animation des studios Pixar sorti en 2015.
Jeux vidéo
- Yoshi : personnage de jeux vidéo de l'univers de Mario.
- Horizon Zero Dawn : l'univers post-apocalyptique du jeu, dans lequel évolue l'héroïne Aloy, contient de nombreux dinosaures mécaniques ;
- ARK: Survival Evolved : jeu de survie contenant plus d'une centaine de dinosaures différents ;
- Jurassic World Evolution : jeu de gestion de parc de dinosaures inspiré du film Jurassic World ;
- King Kong : reprenant le film homonyme de 2005, de nombreux dinosaures sont visible.
Notes et références
Notes
- (en) Cet article est partiellement ou en totalité issu de l’article de Wikipédia en anglais intitulé « Dinosaur » (voir la liste des auteurs).
- En raison des découvertes récentes en paléontologie et en phylogénétique, les expressions « reptile préhistorique », « reptile disparu », « âge des reptiles », « reptile marin » ou « reptile volant » sont aujourd'hui l'objet de controverses de neutralité (neutralité du point de vue) entre le point de vue de la systématique évolutionniste (D. Aubert, Classer le vivant : les perspectives de la systématique évolutionniste moderne, Ellipses 2017) qui y inclut les dinosaures, les ichthyosaures, les plésiosaures, les pliosaures, les ptérosaures et d'autres groupes, et le point de vue cladistique qui les en exclut et qui considère les « reptiles » comme un groupe paraphylétique (G. Lecointre dir., C. Fortin, M.-L. Le Louarn Bonnet, G. Guillot, Guide critique de l'évolution, Belin 2009, (ISBN 978-2-7011-4797-0)). Elles sont donc à manier avec précaution, notamment dans le contexte de la vulgarisation scientifique.
- Reprenant ainsi le nom d'un ordre de dinosaures introduit par Thomas Huxley en 1870.
- Benjamin Waterhouse Hawkins qui trône au centre de la table, a convié 21 scientifiques, journalistes et mécènes attablés dans le reptile placé au centre d'une pièce recouverte de larges tentures rouges. L'événement est destiné à célébrer une science nouvelle, la paléontologie, et honorer des grands noms de paléontologistes, inscrits sur quatre plaques : Richard Owen debout dans la tête du dinosaure, William Buckland, Georges Cuvier et Gideon Mantell, ces deux derniers manquant à l'appel. Mantell dont les travaux ont été pillés par Owen, a en effet trouvé la mort un an auparavant, probablement par suicide. cf. (en) Edwin Harris Colbert, Dinosaurs. Their Discovery and Their World, Dutton, , p. 25.
Références
- Matthew G. Baron; Megan E. Williams (2018). "A re-evaluation of the enigmatic dinosauriform Caseosaurus crosbyensis from the Late Triassic of Texas, USA and its implications for early dinosaur evolution". Acta Palaeontologica Polonica. 63. DOI:10.4202/app.00372.2017.
- Andrea Cau (2018). "The assembly of the avian body plan: a 160-million-year long process" (PDF). Bollettino della Società Paleontologica Italiana. 57 (1): 1–25. DOI:10.4435/BSPI.2018.01.
- Encyclopædia Universalis, « DINOSAURES », sur Encyclopædia Universalis (consulté le )
- (en) M.G. Baron, D.B. Norman et P.M. Barrett, « A new hypothesis of dinosaur relationships and early dinosaur evolution », Nature, vol. 543, , p. 501–506 (DOI 10.1038/nature21700).
- Laurent Sacco, « Dinosaures : faut-il changer leur classification ? », sur Futura-Sciences, .
- Hervé Le Guyader, « Les dinosaures changent d'arbre », Pour la science, no 490, , p. 92-94.
- Éric Buffetaut, Idées reçues sur les dinosaures, Le Cavalier Bleu, , 160 p. (ISBN 9791031803562, lire en ligne), p. 9
- « Dinosaure », dans le Dictionnaire de l'Académie française, sur Centre national de ressources textuelles et lexicales (consulté le 4 janvier 2016).
- Définitions lexicographiques et étymologiques de « dinosaure » dans le Trésor de la langue française informatisé, sur le site du Centre national de ressources textuelles et lexicales (consulté le 4 janvier 2016).
- Entrée « dinosaure (ou dinosorien) », sur larousse.fr/dictionnaires/Français, Dictionnaire de français (en ligne), Larousse (consulté le ).
- (en) Owen, R. (1842), Report on British Fossil Reptiles, Part II. Report of the British Association for the Advancement of Science, Plymouth, England.
- (en) Farlow, J.O., and Brett-Surman, M.K. (1997). Preface. In: Farlow, J.O., and Brett-Surman, M.K. (eds.). The Complete Dinosaur. Indiana University Press : Bloomington and Indianapolis, ix-xi. (ISBN 978-0-253-33349-0).
- « Stegosaur Carving on a Cambodian Temple? by Glen Kuban », sur paleo.cc (consulté le )
- (en) Sharon Siamon, Dinosaurs! Battle of the Bones, Dk Pub, , p. 14.
- (en) Leonardo Salgado et Zulma Gasparini, « Reappraisal of an ankylosaurian dinosaur from the Upper Cretaceous of James Ross Island (Antarctica) », Geodiversitas, vol. 28, no 1, , p. 119-136 (lire en ligne [PDF]).
- (en) Willian R. Hammer et William J. Hickerson, « A Crested Theropod Dinosaur from Antarctica », Science, vol. 264, , p. 828-830 (DOI 10.1126/science.264.5160.828).
- (en) Philip D. Mannion, Ronan Allain et Olivier Moine, « The earliest known titanosauriform sauropod dinosaur and the evolution of Brachiosauridae », Peer J., vol. 5, no e3217, (DOI 10.7717/peerj.3217)
- Padian, K. & May, C.L., 1993. The earliest dinosaurs. In: Lucas, S.G., Morales, M. (Eds.), The Nonmarine Triassic: New Mexico Museum of Natural History and Science Bulletin, vol. 3, p. 379–380.
- Sereno, P.C., 2005. The logical basis of phylogenetic taxonomy. Systems Biology 54, 595–619.
- Brusatte, Stephen L.; Nesbitt, S. J.; Irmis, R. B.; Butler, R. J.; Benton, M. J.; Norell, M.A. (2010). "The origin and early radiation of dinosaurs". Earth-Science Reviews 101:68-100.
- Novas, F.E., 1992. Phylogenetic relationships of the basal dinosaurs, the Herrerasauridae. Palaeontology 16, 51–62.
- Sereno, P.C., 1998. A rationale for phylogenetic definitions, with application to the higher-level taxonomy of Dinosauria. Neues Jahrbuch für Geologie und Paläontologie Abhandlungen 210, 41–83.
- Olshevsky, G., 2000. An annotated checklist of dinosaur species by continent. Mesozoic Meanderings 3:1-157.
- Fraser, N.C., Padian, K., Walkden, G.M., Davis, A.L.M., 2002. Basal dinosauriform remains from Britain and the diagnosis of the Dinosauria. Palaeontology 45, 79–95.
- (en) Padian, K. (2004). Basal Avialae. In: Weishampel, D.B., Dodson, P., and Osmólska, H. (eds.). The Dinosauria (second edition). University of California Press:Berkeley, 210–231. (ISBN 978-0-520-24209-8).
- Peter Dodson, « Origin of Birds: The Final Solution? », Integrative and Comparative Biology, vol. 40, no 4, , p. 504-512
- Brusatte, Stephen L., 2012. Dinosaur Paleobiology. Wiley-Blackwell. 322 p.
- Nesbitt S.J., 2011. The early evolution of archosaurs : relationships and the origin of major clades. Bulletin of the American Museum of Natural History 352: 1–292.
- Benton, M.J., 2012. Origin and evolution of dinosaurs. p. 336-351. In Brett-Surman, M.K., Holtz, T.R., Jr., Farlow, J.O., and Walters, B. (eds), The complete dinosaur, 2nd edition. Indiana University Press, Bloomington, 1115 pp.
- Hatcher, J.B. (1903). "Osteology of Haplocanthosaurus with description of a new species, and remarks on the probable habits of the Sauropoda and the age and origin of the Atlantosaurus beds." Memoirs of the Carnegie Museum, 2: 1 -72, pls 1 4.
- Kubo, T. & Benton, M.J., 2007. Evolution of hindlimb posture in archosaurs: limb stresses in extinct vertebrates. Palaeontology 50 (6): 1519–1529.
- Benton & Clark, J., 1988. Archosaur phylogeny and the relationships of the Crocodylia. 295–338. In Benton, M.J. (ed.). The phylogeny and classification of the tetrapods. Systematics Association, Special Volume, 35A, 377 pp.
- Bonaparte, J.F., 1984. Locomotion in rauisuchid thecodonts. Journal of Vertebrate Paleontology 3 (4): 210–218.
- Benton, M.J., 1984. « Rauisuchians and the success of dinosaurs ». Nature 310 (5973): 101.
- Parrish, J.M., 1986. Locomotor adaptations in the hindlimb and pelvis of the Thecodontia. Hunteria, 1, 1–36.
- Parrish, J.M., 1987. The origin of crocodilian locomotion. Paleobiology, 13, 396–414.
- Molnar, R.E. (1991) The cranial morphology of Tyrannosaurus rex. Palaeontographica Abteilung A 217, 137–176.
- Eddy, D.R. & Clarke, J.A. (2011) New information on the cranial anatomy of Acrocanthosaurus atokensis and its implications for the phylogeny of Allosauroidea (Dinosauria: Theropoda). PloS one 6, e17932.
- Britt, B.B. (1991) Theropods of Dry Mesa Quarry (Morrison Formation, Late Jurassic), Colorado, with emphasis on the osteology of Torvosaurus tanneri. Brigham Young University Geology Studies 37, 1–72.
- Sues, H.-D., Frey, E., Martill, D.M. & Scott, D.M. (2002) Irritator challengeri, a spinosaurid (Dinosauria: Theropoda) from the Lower Cretaceous of Brazil. Journal of Vertebrate Paleontology 22, 535–547.
- Dal Sasso, C., Maganuco, S., Buffetaut, E. & Mendez, M.A. (2005) New information on the skull of the enigmatic theropod Spinosaurus, with remarks on its size and affinities. Journal of Vertebrate Paleontology 25, 888–896.
- L. E. Zanno et Peter J. Makovicky (2011) Herbivorous ecomorphology and specialization patterns in theropod dinosaur evolution. Proceedings of the National Academy of Sciences 108, 232–237.
- Clark, J.M., Norell, M.A. & Rowe, T. (2002) Cranial anatomy of Citipati osmolskae (Theropoda, Oviraptorosauria), and a reinterpretation of the holotype of Oviraptor philoceratops. American Museum Novitates, 1–24.
- Clark, J.M., Altangerel, P., Norell, M. & Akademi, M.S.U. (1994) The skull of Erlicosaurus andrewsi, a late Cretaceous“ Segnosaur”(Theropoda, Therizinosauridae) from Mongolia. American Museum novitates, 39 pp.
- Currie, P.J. & Padian, K. eds. (1997) Encyclopedia of Dinosaurs. Academic Press. 869 pp.
- Weishampel, D.B., Dodson, P. & Osmólska, H. (2004) The Dinosauria: Second Edition. University of California Press. 880 pp.
- Brett-Surman, M.K., Holtz, T.R. & Farlow, J.O. (2012) The Complete Dinosaur, Second Edition. Indiana University Press. 1128 pp.
- Fastovsky, D.E. & Weishampel, D.B. (2012) Dinosaurs: A Concise Natural History. 2nd ed. Cambridge University Press. 416 pp.
- Ryan, M.J., Holmes, R. & Russell, A.P. (2007) A revision of the late campanian centrosaurine ceratopsid genus Styracosaurus from the Western Interior of North America. Journal of Vertebrate Paleontology 27, 944–962.
- Currie, P.J., Langston Jr, W. & Tanke, D.H. (2008) A new species of Pachyrhinosaurus (Dinosauria, Ceratopsidae) from the Upper Cretaceous of Alberta, Canada. A new horned dinosaur from an Upper Cretaceous Bone Bed in Alberta. Ottawa: NRC Research Press. p, 1–108.
- Forster, C.A. (1996) New information on the skull of Triceratops. Journal of Vertebrate Paleontology 16, 246–258.
- Sullivan, R.M. (2003) Revision of the dinosaur Stegoceras Lambe (Ornithischia, Pachycephalosauridae). Journal of Vertebrate Paleontology 23, 181–207.
- Horner, J.R. & Goodwin, M.B. (2009) Extreme Cranial Ontogeny in the Upper Cretaceous Dinosaur Pachycephalosaurus. PLoS ONE 4, e7626.
- Brown, B. (1914) Corythosaurus casuarius, a new crested dinosaur from the Belly River Cretaceous; with Provisional classification of the family Trachodontidae. Bulletin of the American Museum of Natural History 33, 559–565.
- Ostrom, J.H. (1963) Parasaurolophus cyrtocristatus: A Crested Hadrosaurian Dinosaur from New Mexico. Fieldiana Geology, 14, 143–168.
- Carpenter, K. (2004) Redescription of Ankylosaurus magniventris Brown 1908 (Ankylosauridae) from the Upper Cretaceous of the Western Interior of North America. Canadian Journal of Earth Sciences, 41, 961–986.
- Vickaryous, M.K. & Russell, A.P. (2003) A redescription of the skull of Euoplocephalus tutus (Archosauria: Ornithischia): a foundation for comparative and systematic studies of ankylosaurian dinosaurs. Zoological Journal of the Linnean Society, 137, 157–186.
- Dal Sasso, C. and Signore, M. 1998. Exceptional soft-tissue preservation in a theropod dinosaur from Italy. Nature, 392:383–387. DOI:10.1038/32884.
- Dal Sasso, C. and Maganuco, S. 2011. Scipionyx samniticus (Theropoda: Compsognathidae) from the Lower Cretaceous of Italy: osteology, ontogenetic assessment, phylogeny, soft tissue anatomy, taphonomy and palaeobiology. Memorie della Società Italiana di Scienze Naturali e del Museo Civico di Storia Naturale di Milano, 37:1–281.
- Naish, D., Martill, D.M., and Frey, E. 2004. Ecology, Systematics and Biogeographical Relationships of Dinosaurs, Including a New Theropod, from the Santana Formation (?Albian, Early Cretaceous) of Brazil. Historical Biology, 16:57–70. doi:10.1080/08912960410001674200.
- Kellner, A.W.A. 1999. Short note on a new dinosaur (Theropoda, Coelurosauria) from the Santana Formation (Romualdo member, Albian), Northeastern Brazil. Boletim do Museu Nacional, N.S., 49:1–8.
- Briggs, D.E.G., Wilby, P.R., Pérez-Moreno, B.P., Sanz, J.L., and Fregenal-Martínez, M. 1997. The mineralization of dinosaur soft tissue in the Lower Cretaceous of Las Hoyas, Spain. Journal of the Geological Society, 154:587–588. doi:10.1144/gsjgs.154.4.0587.
- Murphy, N.L., Trexler, D., and Thompson, M. 2006. “Leonardo”, a mummified Brachylophosaurus from the Judith River Formation, p. 117–133. In Carpenter, K. (ed.), Horns and Beaks: Ceratopsian and Ornithopod Dinosaurs. Indiana University Press, Bloomington and Indianapolis.
- Manning, P.L., Morris, P.M., McMahon, A., Jones, E., Gize, A., Macquaker, J.H.S., Wolff, G., Thompson, A., Marshall, J., Taylor, K.G., Lyson, T., Gaskell, S., Reamtong, O., Sellers, W.I., Dongen, B.E. van, Buckley, M., and Wogelius, R.A. 2009. Mineralized soft-tissue structure and chemistry in a mummified hadrosaur from the Hell Creek Formation, North Dakota (USA). Proceedings of the Royal Society B: Biological Sciences, 276:3429–3437. doi:10.1098/rspb.2009.0812.
- Fisher, P.E., Russell, D.A., Stoskopf, M.K., Barrick, R.E., Hammer, M., and Kuzmitz, A.A. 2000. Cardiovascular Evidence for an Intermediate or Higher Metabolic Rate in an Ornithischian Dinosaur. Science, 288:503–505. DOI:10.1126/science.288.5465.503.
- Rowe, T., McBride, E.F., and Sereno, P.C. 2001. Dinosaur with a Heart of Stone. Science, 291:783–783. doi:10.1126/science.291.5505.783a.
- Cleland, T.P., Stoskopf, M.K., and Schweitzer, M.H. 2011. Histological, chemical, and morphological reexamination of the “heart” of a small Late Cretaceous Thescelosaurus. Naturwissenschaften, 98:203–211. doi:10.1007/s00114-010-0760-1.
- M. tensor periorbitae - mPT, M. levator pterygoideus - mLPt et M. protractor pterygoideus - mPPt.
- M. pseudotemporalis profundus - mPSTp, M. pterygoideus dorsalis - mPTd, M. pterygoideus ventralis - mPTv et M. adductor mandibulae posterior - mAMP.
- M. adductor mandibulae externus superficialis - mAMES, M. adductor mandibulae externus medialis - mAMEM, M. adductor mandibulae externus profundus - mAMEP et M. pseudotemporalis superficialis - mPSTs.
- M. depressor mandibulae - mDM.
- Holliday, C.M. 2009. New Insights Into Dinosaur Jaw Muscle Anatomy. The Anatomical Record: Advances in Integrative Anatomy and Evolutionary Biology, 292:1246–1265. doi:10.1002/ar.20982.
- Snively, E. and Russell, A.P. 2007a. Functional variation of neck muscles and their relation to feeding style in Tyrannosauridae and other large theropod dinosaurs. The Anatomical Record: Advances in Integrative Anatomy and Evolutionary Biology, 290:934–957. doi:10.1002/ar.20563.
- Snively, E. and Russell, A.P. 2007b. Functional morphology of neck musculature in the Tyrannosauridae (Dinosauria, Theropoda) as determined via a hierarchical inferential approach. Zoological Journal of the Linnean Society, 151:759–808. doi:10.1111/j.1096-3642.2007.00334.x.
- Snively, E. and Russell, A.P. 2007c. Craniocervical feeding dynamics of Tyrannosaurus rex. Paleobiology, 33:610–638. doi:10.1666/06059.1.
- Tsuihiji, T. 2010. Reconstructions of the Axial Muscle Insertions in the Occipital Region of Dinosaurs: Evaluations of Past Hypotheses on Marginocephalia and Tyrannosauridae Using the Extant Phylogenetic Bracket Approach. The Anatomical Record: Advances in Integrative Anatomy and Evolutionary Biology, 293:1360–1386. doi:10.1002/ar.21191.
- Schwarz, D., Frey, E., and Meyer, C.A. 2007. Pneumaticity and soft-tissue reconstructions in the neck of diplodocid and dicraeosaurid sauropods. Acta Palaeontologica Polonica, 52:167.
- Carrano, M.T. and Hutchinson, J.R. 2002. Pelvic and hindlimb musculature of Tyrannosaurus rex (Dinosauria: Theropoda). Journal of Morphology, 253:207–228. doi:10.1002/jmor.10018.
- Hutchinson, J.R., Anderson, F.C., Blemker, S.S., and Delp, S.L. 2005. Analysis of hindlimb muscle moment arms in Tyrannosaurus rex using a three-dimensional musculoskeletal computer model: implications for stance, gait, and speed. Paleobiology, 31:676–701. doi:10.1666/0094-8373(2005)031[0676:AOHMMA]2.0.CO;2.
- Jasinoski, S.C., Russell, A.P., and Currie, P.J. 2006. An integrative phylogenetic and extrapolatory approach to the reconstruction of dromaeosaur (Theropoda: Eumaniraptora) shoulder musculature. Zoological Journal of the Linnean Society, 146:301–344. doi:10.1111/j.1096-3642.2006.00200.x.
- Hutchinson, J.R. and Gatesy, S.M. 2000. Adductors, abductors, and the evolution of archosaur locomotion. Paleobiology, 26:734–751. doi:10.1666/0094-8373(2000)026<0734:AAATEO>2.0.CO;2.
- Hutchinson, J.R. 2006. The evolution of locomotion in archosaurs. Comptes Rendus Palevol, 5:519–530. doi:10.1016/j.crpv.2005.09.002.
- Hutchinson, J.R. and Gatesy, S.M. 2006. Dinosaur Locomotion: Beyond the bones. Nature, 440:292–294. doi:10.1038/440292a.
- Sampson, S.D. and Witmer, L.M. 2007. Craniofacial Anatomy of Majungasaurus crenatissimus (theropoda: Abelisauridae) from the Late Cretaceous of Madagascar. Journal of Vertebrate Paleontology, 27:32–104. doi:10.1671/0272-4634(2007)27[32:CAOMCT]2.0.CO;2.
- Witmer, L.M. and Ridgely, R.C. 2009. New Insights Into the Brain, Braincase, and Ear Region of Tyrannosaurs (Dinosauria, Theropoda), with Implications for Sensory Organization and Behavior. The Anatomical Record: Advances in Integrative Anatomy and Evolutionary Biology, 292:1266–1296. doi:10.1002/ar.20983.
- Gabe S. Bever, Stephen L. Brusatte, Amy M. Balanoff, et Mark A. Norell, 2011. "Variation, Variability, and the Origin of the Avian Endocranium: Insights from the Anatomy of Alioramus altai (Theropoda: Tyrannosauroidea)". PloS one, 6:e23393.
- Evans, D.C., Ridgely, R., and Witmer, L.M. 2009. Endocranial Anatomy of Lambeosaurine Hadrosaurids (Dinosauria: Ornithischia): A Sensorineural Perspective on Cranial Crest Function. The Anatomical Record: Advances in Integrative Anatomy and Evolutionary Biology, 292:1315–1337. doi:10.1002/ar.20984.
- Witmer, L.M. and Ridgely, R.C. 2008. Structure of the brain cavity and inner ear of the centrosaurine ceratopsid Pachyrhinosaurus based on CT scanning and 3D visualization, p. 117–144. In Currie, P.J. (ed.), A new horned dinosaur from an Upper Cretaceous bone bed. National Research Council Research Press, Ottawa.
- Witmer, L.M., Ridgely, R.C., Dufeau, D.L., and Semones, M.C. 2008. Using CT to Peer into the Past: 3D Visualization of the Brain and Ear Regions of Birds, Crocodiles, and Nonavian Dinosaurs, p. 67–87. In Endo, H. and Frey, R. (eds.), Anatomical Imaging. Springer Japan, Tokyo.
- Sereno, P.C., Wilson, J.A., Witmer, L.M., Whitlock, J.A., Maga, A., Ide, O., and Rowe, T.A. 2007. Structural Extremes in a Cretaceous Dinosaur. PLoS ONE, 2:e1230. doi:10.1371/journal.pone.0001230.
- Larsson, H.C.E., Sereno, P.C., and Wilson, J.A. 2000. Forebrain enlargement among nonavian theropod dinosaurs. Journal of Vertebrate Paleontology, 20:615–618. doi:10.1671/0272-4634(2000)020[0615:FEANTD]2.0.CO;2.
- Domínguez Alonso, P., Milner, A.C., Ketcham, R.A., Cookson, M.J., and Rowe, T.B. 2004. The avian nature of the brain and inner ear of Archaeopteryx. Nature, 430:666–669. doi:10.1038/nature02706.
- (en) Martin D. Brasier, David B. Norman2, Alexander G. Liu, Laura J. Cotton, Jamie E. H. Hiscocks, Russell J. Garwood, Jonathan B. Antcliffe et David Wacey, « Remarkable preservation of brain tissues in an Early Cretaceous iguanodontian dinosaur », Geological Society, London, Special Publications, vol. 448, (DOI 10.1144/SP448.3).
- Laurent Sacco, « Un cerveau de dinosaure fossilisé découvert pour la première fois ! », .
- Czerkas, S. 1997. Skin, p. 1100–1116. In Currie, P.J. and Padian, K. (eds.), Encyclopedia of Dinosaurs. Academic Press. San Diego, California.
- Bell, P.R. 2012. Standardized Terminology and Potential Taxonomic Utility for Hadrosaurid Skin Impressions: A Case Study for Saurolophus from Canada and Mongolia. PLoS ONE, 7:e31295. doi:10.1371/journal.pone.0031295.
- Brown, B. 1917. A complete skeleton of the horned dinosaur Monoclonius, and description of a second skeleton showing skin impressions. Bulletin of the American Museum of Natural History, 37:298–306.
- Sternberg, C.M. 1925. Integument of Chasmosaurus belli. Canadian Field Naturalist, 39:108–110.
- Xing, L.D., Peng, G.Z., and Shu, C.K. 2008. Stegosaurian skin impressions from the Upper Jurassic Shangshaximiao Formation, Zigong, Sichuan, China: A new observation. Geological Bulletin of China, 27:1049–1053.
- Christiansen, N.A. and Tschopp, E. 2010. Exceptional stegosaur integument impressions from the Upper Jurassic Morrison Formation of Wyoming. Swiss Journal of Geosciences, 103:163–171. doi:10.1007/s00015-010-0026-0.
- Martill, D.M., Batten, D.J., and Loydell, D.K. 2000. A new specimen of the thyreophoran dinosaur cf. Scelidosaurus with soft tissue preservation. Palaeontology, 43:549–559. doi:10.1111/j.0031-0239.2000.00139.x.
- Arbour, V., Lech-Hernes, N.L., Guldberg, T.E., Hurum, J.H., and Currie, P.J. 2012. An ankylosaurid dinosaur from Mongolia with in situ armour and keratinous scale impressions. Acta Palaeontologica Polonica. doi:10.4202/app.2011.0081.
- Coria, R.A. and Chiappe, L.M. 2007. Embryonic skin from Late Cretaceous sauropods (Dinosauria) of Auca Mahuevo, Patagonia, Argentina. Journal of Paleontology, 81:1528–1532. doi:10.1666/05-150.1.
- Bonaparte, J.F. and Powell, J.E. 1980. A continental assemblage of tetrapods from the Upper Cretaceous beds of El Brete, northwestern Argentina (Sauropoda-Coelurosauria-Carnosauria-Aves). Mémoires de la Société Géologique de France, Nouvelle Série, 139:19–28.
- Giménez, O. del V. 2007. Skin impressions of Tehuelchesaurus (Sauropoda) from the Upper Jurassic of Patagonia. Revista del Museo Argentino de Ciencias Naturales, 9:119–124.
- Bonaparte, J.F., Novas, F.E., and Coria, R.A. 1990. Carnotaurus sastrei Bonaparte, the horned, lightly built carnosaur from the Middle Cretaceous of Patagonia. Natural History Museum of Los Angeles County Contributions in Science, 416:1–42.
- Chiappe, L.M. and Göhlich, U.B. 2010. Anatomy of Juravenator starki (Theropoda: Coelurosauria) from the Late Jurassic of Germany. Neues Jahrbuch für Geologie und Paläontologie - Abhandlungen, 258:257–296. doi:10.1127/0077-7749/2010/0125.
- Zheng, X.-T., You, H.-L., Xu, X., and Dong, Z.-M. 2009. An Early Cretaceous heterodontosaurid dinosaur with filamentous integumentary structures. Nature, 458:333–336. doi:10.1038/nature07856.
- Mayr, G., Peters, S.D., Plodowski, G., and Vogel, O. 2002. Bristle-like integumentary structures at the tail of the horned dinosaur Psittacosaurus. Naturwissenschaften, 89:361–365. doi:10.1007/s00114-002-0339-6.
- Rauhut, O.W.M., Foth, C., Tischlinger, H., and Norell, M.A. 2012. Exceptionally preserved juvenile megalosauroid theropod dinosaur with filamentous integument from the Late Jurassic of Germany. Proceedings of the National Academy of Sciences, 109:11746–11751. doi:10.1073/pnas.1203238109.
- Xu, X., Norell, M.A., Kuang, X., Wang, X., Zhao, Q., and Jia, C. 2004. Basal tyrannosauroids from China and evidence for protofeathers in tyrannosauroids. Nature, 431:680–684. doi:10.1038/nature02855.
- Chen, P., Dong, Z., and Zhen, S. 1998. An exceptionally well-preserved theropod dinosaur from the Yixian Formation of China. Nature, 391:147–152. doi:10.1038/34356.
- Chiappe, L.M. and Göhlich, U.B. 2010. Anatomy of Juravenator starki (Theropoda: Coelurosauria) from the Late Jurassic of Germany. Neues Jahrbuch für Geologie und Paläontologie - Abhandlungen, 258:257–296. doi:10.1127/0077-7749/2010/0125.
- Schweitzer, M.H., Watt, J.A., Avci, R., Knapp, L., Chiappe, L., Norell, M., and Marshall, M. 1999. Beta-keratin specific immunological reactivity in feather-like structures of the Cretaceous Alvarezsaurid, Shuvuuia deserti. Journal of Experimental Zoology, 285:146–157. doi:10.1002/(SICI)1097-010X(19990815)285:2<146::AID-JEZ7>3.0.CO;2-A.
- Xu, X., Tang, Z., and Wang, X. 1999. A therizinosauroid dinosaur with integumentary structures from China. Nature, 399:350–354. doi:10.1038/20670.
- Xu, X., Zheng, X., and You, H. 2009. A new feather type in a nonavian theropod and the early evolution of feathers. Proceedings of the National Academy of Sciences, 106:832–834. doi:10.1073/pnas.0810055106.
- Qiang, J., Currie, P.J., Norell, M.A., and Shu-An, J. 1998. Two feathered dinosaurs from northeastern China. Nature, 393:753–761. doi:10.1038/31635.
- Xu, X., Wang, X.-L., and Wu, X.-C. 1999. A dromaeosaurid dinosaur with a filamentous integument from the Yixian Formation of China. Nature, 401:262–266. doi:10.1038/45769.
- Ji, Q., Norell, M.A., Gao, K.-Q., Ji, S.-A., and Ren, D. 2001. The distribution of integumentary structures in a feathered dinosaur. Nature, 410:1084–1088. doi:10.1038/35074079.
- Xu, X., Zhou, Z., and Prum, R.O. 2001. Branched integumental structures in Sinornithosaurus and the origin of feathers. Nature, 410:200–204. doi:10.1038/35065589.
- Dyke, G.J. and Norell, M.A. 2005. Caudipteryx as a non-avialan theropod rather than a flightless bird. Acta Palaeontologica Polonica, 50:101–116.
- Xu, X., Zheng, X., and You, H. 2010. Exceptional dinosaur fossils show ontogenetic development of early feathers. Nature, 464:1338–1341. doi:10.1038/nature08965.
- Xu, X., Zhou, Z., Wang, X., Kuang, X., Zhang, F., and Du, X. 2003. Four-winged dinosaurs from China. Nature, 421:335–340. doi:10.1038/nature01342.
- Hu, D., Hou, L., Zhang, L., and Xu, X. 2009. A pre-Archaeopteryx troodontid theropod from China with long feathers on the metatarsus. Nature, 461:640–643. doi:10.1038/nature08322.
- (en) Xing Xu, Philip Currie, Michael Pittman, Lida Xing, Qingjin Meng, Junchang Lü, Dongyu Hu et Congyu Yu, « Mosaic evolution in an asymmetrically feathered troodontid dinosaur with transitional features », Nature Communications, (DOI 10.1038/ncomms14).
- (en) A. H. Turner, Peter J. Makovicky et M. A. Norell, « Feather Quill Knobs in the Dinosaur Velociraptor », Science, vol. 317, , p. 1721–1721 (DOI 10.1126/science.1145076).
- Ortega, F., Escaso, F., and Sanz, J.L. 2010. A bizarre, humped Carcharodontosauria (Theropoda) from the Lower Cretaceous of Spain. Nature, 467:203–206. doi:10.1038/nature09181.
- (en) Lida Xing et al., « A Feathered Dinosaur Tail with Primitive Plumage Trapped in Mid-Cretaceous Amber] », Current Biology, vol. 26, no 24, , p. 3352-3360 (DOI 10.1016/j.cub.2016.10.008).
- Ryan, M.J. and Russell, A.P. 1997. Color, p. 134–135. In Currie, P.J. and Padian, K. (eds.), Encyclopedia of Dinosaurs. Academic Press.
- Zhang, F., Kearns, S.L., Orr, P.J., Benton, M.J., Zhou, Z., Johnson, D., Xu, X., and Wang, X. 2010. Fossilized melanosomes and the colour of Cretaceous dinosaurs and birds. Nature, 463:1075–1078. doi:10.1038/nature08740.
- Li, Q., Gao, K.-Q., Vinther, J., Shawkey, M.D., Clarke, J.A., D’Alba, L., Meng, Q., Briggs, D.E.G., & Prum, R.O. 2010. Plumage Color Patterns of an Extinct Dinosaur. Science, 327:1369–1372. doi:10.1126/science.1186290.
- Lingham-Soliar, T. 2003. Evolution of birds: ichthyosaur integumental fibers conform to dromaeosaur protofeathers. Naturwissenschaften, 90:428–432. doi:10.1007/s00114-003-0448-x.
- Feduccia, A., Lingham-Soliar, T., & Hinchliffe, J.R. 2005. Do feathered dinosaurs exist? Testing the hypothesis on neontological and paleontological evidence. Journal of Morphology, 266:125–166. doi:10.1002/jmor.10382.
- Lingham-Soliar, T., Feduccia, A., and Wang, X. 2007. A new Chinese specimen indicates that “protofeathers” in the Early Cretaceous theropod dinosaur Sinosauropteryx are degraded collagen fibres. Proceedings of the Royal Society B: Biological Sciences, 274:1823–1829. doi:10.1098/rspb.2007.0352.
- Li, Q., Gao, K.-Q., Meng, Q., Clarke, J.A., Shawkey, M.D., D’Alba, L., Pei, R., Ellison, M., Norell, M.A. & Vinther, J. 2012. Reconstruction of Microraptor and the Evolution of Iridescent Plumage. Science 335:1215–1219. doi:10.1126/science.1213780.
- Godefroit, P., Demuynck, H., Dyke, G., Hu, D., Escuillié, F. & Claeys, P. 2013. Reduced plumage and flight ability of a new Jurassic paravian theropod from China. Nature Communications 4:1394. doi:10.1038/ncomms2389.
- Xu, X., Zhao, Q., Norell, M., Sullivan, C., Hone, D., Erickson, G., Wang, X., Han, F. and Guo, Y. 2009. A new feathered maniraptoran dinosaur fossil that fills a morphological gap in avian origin. Chinese Science Bulletin 54:430–435. doi:10.1007/s11434-009-0009-6.
- Upchurch, P., Barrett, P.M. & Dodson, P. 2004. Sauropoda, p. 259–322. The Dinosauria. Second edition. University of California Press, Berkeley, CA.
- Sander, P.M., Christian, A., Clauss, M., Fechner, R., Gee, C.T., Griebeler, E.-M., Gunga, H.-C., Hummel, J., Mallison, H., Perry, S.F., Preuschoft, H., Rauhut, O.W.M., Remes, K., Tutken, T., Wings, O. & Witzel, U. 2011. Biology of the sauropod dinosaurs: the evolution of gigantism. Biological Reviews of the Cambridge Philosophical Society 86:117–155. doi:10.1111/j.1469-185X.2010.00137.x.
- Therrien, F. and Henderson, D.M. 2007. My theropod is bigger than yours… or not: estimating body size from skull length in theropods. Journal of Vertebrate Paleontology 27:108–115. doi:10.1671/0272-4634(2007)27[108:MTIBTY]2.0.CO;2.
- Sereno, P.C. 1999. The Evolution of Dinosaurs. Science 284:2137–2147. doi:10.1126/science.284.5423.2137.
- Carrano, M.T. 2006. Body-Size Evolution in the Dinosauria, p. 225–268. In Carrano, M., Gaudin, T.J., Blob, R.W., and Wible, J.R. (eds.), Amniote Paleobiology: Perspectives on the Evolution of Mammals, Birds, and Reptiles. University of Chicago Press.
- Gunga, H.-C., Suthau, T., Bellmann, A., Friedrich, A., Schwanebeck, T., Stoinski, S., Trippel, T., Kirsch, K. & Hellwich, O. 2007. Body mass estimations for Plateosaurus engelhardti using laser scanning and 3D reconstruction methods. Naturwissenschaften 94:623–630. doi:10.1007/s00114-007-0234-2.
- (en) Jonah N. Choiniere , Xing Xu, James M. Clark, Catherine A. Forster, Yu Guo, Fenglu Han, « A basal alvarezsauroid theropod from the early late Jurassic of Xinjiang, China », Science, vol. 327, no 5965, , p. 571 (DOI 10.1126/science.1182143)
- O’Gorman, E.J. and Hone, D.W.E. 2012. Body Size Distribution of the Dinosaurs. PLoS ONE 7:e51925. doi:10.1371/journal.pone.0051925.
- Erickson, B. 1990. On the Origin of Dinosaurs and Mammals. http://www.frontier-knowledge.com/earth/papers/Origin%20of%20Dinosaurs%20and%20Mammals.pdf
- Gregory, W.K. 1905. The weight of the Brontosaurus. Science 22:572–572.
- Colbert, E.H. 1962. The weights of dinosaurs. American Museum of Natural History 2076:1–16.
- (en) Colbert, E.H. (1968). Men and Dinosaurs: The Search in Field and Laboratory. E. P. Dutton & Company:New York, vii + 283 p. (ISBN 978-0-14-021288-4).
- (en) Carpenter, K. (2006). BIGGEST OF THE BIG: A CRITICAL RE-EVALUATION OF THE MEGA-SAUROPOD Amphicoelias fragillimus. In: Foster, J.R., and Lucas, S.G. (eds.). Paleontology and Geology of the Upper Jurassic Morrison Formation. New Mexico Museum of Natural History and Science Bulletin 36: 131–138.
- (en) dal Sasso, C., Maganuco, S., Buffetaut, E., and Mendez, M.A. (2006). New information on the skull of the enigmatic theropod Spinosaurus, with remarks on its sizes and affinities. Journal of Vertebrate Paleontology 25(4):888–896.
- Johan Yans, Jean Dejax, Denise Pons, Christian Dupuis, Philippe Taquet.(2005) Palaeontological and geodynamical implications of the palynological dating of the wealden facies sediments of Bernissart (Mons Basin, Belgium). C. R. Palevol 4 (2005) 135–150.
- (en) Deposition of Iguanodon skeletons occurred in at least 3 different events. Bernissart Iguanodons and their significance (long) date : 4 Jul. 2001.
- Day, J.J. and Upchurch, P. (2002). Sauropod Trackways, Evolution, and Behavior. Science 296:1659. voir commentaires dans l'article
- J.R. Horner, R. Makela, « Nest of Juveniles Provides Evidence of Family-Structure Among Dinosaurs », dans Nature, 1979, 282 (5736), p. 296–298.
- (en) Jonathan Lambert, « Ancient Mongolian nests show that dinosaurs protected their eggs », Nature, vol. 571, , p. 308-309 (DOI 10.1038/d41586-019-02174-7).
- (en) Kohei Tanaka, Yoshitsugu Kobayashi, Darla K. Zelenitsky, François Therrien, Yuong-Nam Lee et al., « Exceptional preservation of a Late Cretaceous dinosaur nesting site from Mongolia reveals colonial nesting behavior in a non-avian theropod », Geology, (DOI 10.1130/G46328.1)
- Oviraptor nesting Oviraptor nests or Protoceratops ?
- Dinosaur family tracks
- Gerald Grellet-Tinner& Lucas E. Fiorelli ; « A new Argentinean nesting site showing neosauropod dinosaur reproduction in a Cretaceous hydrothermal environment », Nat. Commun., Volume 1, Article 32 ; DOI:doi:10.1038/ncomms1031 ; 29 June 2010, consulté le 2010/07/02.
- (en) Carolyn Gramling, « Dinosaur babies took a long time to break out of their shells », sur Science, .
- (en) Gregory M. Erickson, Darta K. Zelenitski, David Ian Kay et Mark A. Norell, « Dinosaur incubation periods directly determined from growth-line counts in embryonic teeth show reptilian-grade development », PNAS, (DOI 10.1073/pnas.1613716114, résumé).
- (en) Enrique Fiesta, John Davidson, Introduction to Dinosaurs, Mendon Cottage Books, , p. 17-19.
- (en) Oliver Wings, Thomas Tütken, Denver W. Fowler, Thomas Martin, Hans-Ulrich Pfretzschner & Ge Sun, « Dinosaur teeth from the Jurassic Qigu and Shishugou Formations of the Junggar Basin (Xinjiang/China) and their paleoecologic implications », Paläontologische Zeitschrift, vol. 89, no 3, , p. 485-502 (DOI 10.1007/s12542-014-0227-3).
- « Le menu des dinosaures révélé par le calcium », sur CNRS, (consulté le ).
- (en) A. Hassler, J. E. Martin, R. Amiot, T. Tacail, F. Arnaud Godet, R. Allain et V. Balter, « Calcium isotopes offer clues on resource partitioning among Cretaceous predatory dinosaurs », Proceedings of the Royal Society B, (DOI 10.1098/rspb.2018.0197)
- T. Haines, P. Chambers, The Complete Guide to Prehistoric Life, Firefly Books Ltd, , p. 118–119.
- Joined forever in death The discovery of two fossil dinosaurs entangled together proved many theories.
- Cannibalistic Dinosaur The mystery of a dinosaur cannibal.
- Rogers, R.R., Krause, D.W. and Rogers, K.C. (2003). Cannibalism in the Madagascan dinosaur Majungatholus atopus. OLPA Dinosaur Cannibal Unearthed in Madagascar. Nature 422:515-518.
- (en) Gait and Dinosaur speed formules de détermination de la vitesse des dinosaures
- (en) Calculate your own Dinosaur speed autres formules
- (en) Douglas, K. and Young, S. (1998). The dinosaur detectives. MEGALANIA DINOSAUR NEWS PAGE New Scientist 2130:24.
- (en) Hecht, J. (1998). The deadly dinos that took a dive. New Scientist 2130. BBC Dino-sore.
- (en) Henderson, D.M. (2003). Effects of stomach stones on the buoyancy and equilibrium of a floating crocodilian: A computational analysis. The “Inflatable” Dinosaurs of the Mesozoic. Canadian Journal of Zoology 81:1346–1357.
- « Certains dinosaures vivaient surtout la nuit », sur MaxiSciences,
- « Les dinosaures: diurnes ou nocturnes? », sur DinoNews,
- (en) Lars Schmitz & Ryosuke Motani, « Nocturnality in Dinosaurs Inferred from Scleral Ring and Orbit Morphology », Science, (lire en ligne)
- (en) Des dinosaures à sang chaud au Crétacé ?
- (en) Les dinosaures avaient le sang chaud, R. Amiot, Pour la science, octobre 2006, p. 70-74.
- (en) J. L Gillooly, A. P. Allen et E. L. Charnov, « Dinosaur Fossils Predict Body Temperatures », PLoS Biology, vol. 4, no 8, , e248 (DOI 10.1371/journal.pbio.0040248, lire en ligne)
- (en) R. A. Eagle, T. Tütcken, T. S. Martin, A. K. Tripati, H. C. Fricke, M. Conneli, R. L. Cifelli et J. M. Eiler, « Dinosaur Body Temperatures Determined from Isotopic (13C-18O) Ordering in Fossil Biominerals », Science, vol. 333, , p. 443-445 (DOI 10.1126/science.1206196)
- (en) Robert A. Eagle, Marcus Enriquez, Gerald Grellet-Tinner, Alberto Pérez-Huerta, David Hu, Thomas Tütken, Shaena Montanari, Sean J. Loyd, Pedro Ramirez, Aradhna K. Tripati, Matthew J. Kohn, Thure E. Cerling, Luis M. Chiappe & John M. Eiler, « Isotopic ordering in eggshells reflects body temperatures and suggests differing thermophysiology in two Cretaceous dinosaurs », Nature Communications, vol. 6, no 8296, (DOI 10.1038/ncomms9296)
- Sarda Sahney et Michael J. Benton, « Recovery from the most profound mass extinction of all time », Proceedings of the Royal Society B, vol. 275, , p. 759-765 (DOI 10.1098/rspb.2007.1370).
- (en) Douglas H. Erwin, « The Permo-Triassic extinction », dans Shaking the Tree: Readings from Nature in the History of Life, (ISBN 0-226-28496-4, lire en ligne), p. 189
- (en) H. G. Seeley, « On the Classification of the Fossil Animals Commonly Named Dinosauria », Proceedings of the Royal Society of London, vol. 43, 1887 - 1888, p. 165-171 (lire en ligne)
- Ronan Allain et Émilie Läng, « Origine et évolution des saurischiens », Comptes Rendus Palevol, vol. 8, nos 2-3, , p. 243 - 256 (ISSN 1631-0683, DOI DOI: 10.1016/j.crpv.2008.09.013, lire en ligne)
- Science-et-vie.com, « Paléontologie : l'apparition des dinosaures est plus vieille qu'on ne le pensait ! - Science & Vie », sur www.science-et-vie.com, (consulté le )
- (en) Michael J. Benton, Massimo Bernardi et Cormac Kinsella, « The Carnian Pluvial Episode and the origin of dinosaurs », Journal of the Geological Society, vol. 175, no 6, , p. 1019–1026 (ISSN 0016-7649, DOI 10.1144/jgs2018-049, lire en ligne, consulté le )
- « Dans les petits pas des premiers dinosaures terrestres », Sciences et Avenir, site en ligne du magazine, article daté du 07/10/10, citant l'article « Footprints pull origin and diversification of dinosaur stem lineage deep into Early Triassic » publié dans le Proceedings of the Royal Society B par Stephen L. Brusatte, Grzegorz Niedźwiedzki et Richard J. Butler.
- « Le plus vieux des dinosaures ? », Le Monde.fr, (lire en ligne, consulté le )
- (en) Max C. Langer, Martin D. Ezcurra, Jonathas S. Bittencourt et Fernando E. Novas, « The origin and early evolution of dinosaurs », Biological Reviews, vol. 84, , p. 1-56 (DOI 10.1111/j.1469-185X.2009.00094.x).
- New evidence on dinosaur and mammal origins from the Ischigualasto Formation (Upper Triassic, Argentina, Journal of Vertebrate Paleontology volume 8, issue 3, par P.C. Sereno, F.E. Novas ; A.B. Arcucci et C. Yu, 1988.
- (en) « Dinosaurs ended -- and originated -- with a bang! », sur ScienceDaily (consulté le )
- Nouvel Obs.com (12/06/2008)La fin des dinosaures.
- « La chute d'une météorite a bien été fatale aux dinosaures », sur www.lemonde.fr, (consulté le )
- Sans l'astéroïde géante, les dinosaures auraient quand même disparu
- Ryan C. McKellar, Brian D. E. Chatterton, Alexander P. Wolfe et Philip J. Currie, A Diverse Assemblage of Late Cretaceous Dinosaur and Bird Feathers from Canadian Amber ; Science 16 September 2011: 1619-1622
- R. A. Wogelius, P. L. Manning, H. E. Barden, N. P. Edwards, S. M. Webb, W. I. Sellers, K. G. Taylor, P. L. Larson, P. Dodson, H. You, L. Da-qing et U. Bergmann, Trace Metals as Biomarkers for Eumelanin Pigment in the Fossil Record ; Science 2011-09-16, no 1622-1626 ; online 30 juin 2011
- (en) Darren Naish, « The "Birds Are Not Dinosaurs" Movement », sur blogs.scientificamerican.com, (consulté le ).
- (en) Fountain, Henry. "Many more dinosaurs still to be found." New York Times : 12 septembre 2006. Member Center
- (en) Wang, S.C., and Dodson, P. (2006).NCBI Estimating the Diversity of Dinosaurs. ; PNAS Estimating the Diversity of Dinosaurs. Proceedings of the National Academy of Sciences, États-Unis 103:37, p. 13601-13605.
- Dinosauria, le musée des dinosaures à Espéraza, dans l'Aude.
Voir aussi
Bibliographie
- (en) Michael K. Brett-Surman, Thomas Holtz Jr. & James O. Farlow, The Complete Dinosaur (2nd edition), Indiana University Press, , 1128 p.
- (en) Steve Brusatte, Dinosaur Paleobiology, Wiley-Blackwell., , 322 p.
- (en) David E. Fastovsky & David B. Weishampel, Dinosaurs: A Concise Natural History. (2nd edition), Cambridge University Press., , 416 p.
- (en) David Weishampel, Peter Dodson & Halszka Osmólska, The Dinosauria (2nd edition), University of California Press, , 861 p..
- (en) Philip J. Currie & Kevin Padian, Encyclopedia of Dinosaurs., Academic Press., , 869 p.
- Patrice Lebrun, Dinosaures Carnivores (Tome 1: histoire des découvertes, anatomie et physiologie, phylogénèse, théropodes basaux et carnosauriens), Minéraux et Fossiles (Hors-Série no 18), , 128 p.
- Patrice Lebrun, Dinosaures Carnivores (Tome 2: tétanuriens et coelurosauriens basaux), Minéraux et Fossiles (Hors-Série no 20), , 271 p.
- La Fin des dinosaures : comment les grandes extinctions ont façonné le monde vivant. Éric Buffetaut, Fayard, 2003 (ISBN 978-2-213-61489-2) ;
- L’Arbre à remonter le temps : les rencontres de la systématique et de l'évolution. Pascal Tassy, Éd. C. Bourgois, 1991 (ISBN 978-2-267-00712-1) ;
- Comme les huit doigts de la main : réflexions sur l'histoire naturelle. Stephen Jay Gould, Éd. du Seuil, 1996, (ISBN 978-2-02-019426-6 et 0-393-03416-X) ;
- Jean-Guy Michard, Le Monde perdu des dinosaures, coll. « Découvertes Gallimard / Sciences et techniques » (no 69), 1989 (ISBN 2-07-053087-6).
- Pascal Hachet, Dinosaures sur le divan. Psychanalyse de Jurassic Park, éditions Aubier, 1998 (ISBN 2-7007-2405-4).
- Ronan Allain, "Histoire des dinosaures", éditions Perrin, 2012
Articles connexes
Cryptozoologie
En cryptozoologie, certains cryptides sont présentés comme des dinosaures survivants, par exemple le Mokele-mbembe ou le Ngoubou, deux cryptides d'Afrique centrale qui seraient respectivement un sauropode et un cératopsien. Cependant, l'immense majorité des paléontologues et des zoologistes réfutent cette hypothèse, notamment parce qu'il n'y a aucune preuve et qu'aucun fossile de dinosaure non avien postérieur à l'extinction K/T n'a été découvert.
Références taxinomiques
- (en) Référence Tree of Life Web Project : Dinosauria
- (en) Référence FishBase :
- (en) Référence Paleobiology Database : Dinosauria Owen 1842
- (en) Référence NCBI : Dinosauria (taxons inclus)
- (en) Référence uBio : Dinosauria
Liens externes
- (fr) DinoNews: Le site d'un professeur de sciences naturelles passionné par les dinosaures.
- (fr) Le monde des dinosaures: Description de 150 dinosaures et de leur cadre de vie.
- (fr) : Site de l'exposition du Muséum : Dinosaure, la vie en grand
- (fr) Les dinosaures: Le dossier vidéo de francetv éducation.
- (fr) Dinosauria: Site canadien complet sur les dinosaures.
- [vidéo] Que sait-on vraiment des dinosaures ? sur YouTube, chaine boussole.
- Ressources relatives au vivant :
- (cs + en) BioLib
- (en) Paleobiology Database
- (en) Universal Biological Indexer and Organizer
- Ressource relative à la littérature :
- Ressource relative à la santé :
- Ressource relative à la bande dessinée :
- (en) Comic Vine
- Notices dans des dictionnaires ou encyclopédies généralistes :
 Portail des dinosaures
Portail des dinosaures  Portail de l'ornithologie
Portail de l'ornithologie