Pterosauria
Ptérosaures

Sous-ordres de rang inférieur
Les ptérosaures (Pterosauria) sont un ordre éteint de reptiles volants[alpha 1] qui ont existé pendant presque toute la durée du Mésozoïque : du Trias supérieur, il y a 230 millions d’années (Ma), jusqu'à la fin du Crétacé, il y a 66 Ma[1],[2],[3].
Ce sont des diapsides archosaures ornithodiriens, mais ce ne sont pas des dinosaures, et donc pas des oiseaux.
Il semble que les ptérosaures aient été les premiers vertébrés volants. Ils possédaient de grandes ailes formées d'une série de membranes : la plus importante, le cheiropatagium (aussi appelé brachiopatagium) fixée au quatrième très long doigt de leurs membres antérieurs, une autre entre le poignet et le tronc, appelée propatagium, soutenue par l'os ptéroïde, et enfin une troisième rattachée à la cuisse et à la queue : le cruropatagium. Leur répartition a été mondiale : un grand nombre de fossiles, souvent mal conservés, ont été découverts sur tous les continents[1],[2].
Plusieurs espèces au moins étaient couvertes de duvet (le premier spécimen fossile qui révéla cela étant Sordes pilosus). Le plus petit fossile connu en 2008 est celui de Nemicolopterus, une espèce de la taille d'un moineau[4]. Les plus grandes espèces n'ont pas d'équivalent en taille dans le monde contemporain : on a découvert des spécimens de Quetzalcoatlus et de Hatzegopteryx de plus de 12 m d'envergure et pesant sans doute plus de 300 kg[5],[6]. Les ptérosaures se sont adaptés pour occuper de nombreuses niches écologiques, aussi bien terrestres que marines, actuellement prises par les oiseaux ou les chauves-souris.
Découvertes
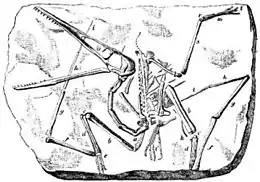
Le premier ptérosaure fossile a été décrit par le naturaliste italien Cosimo Collini en 1784. Il était issu de dépôts calcaires du Jurassique, près de Solnhofen[7]. Collini interpréta ce qu'il voyait de manière erronée et décrivit une créature aquatique marine. Cette interprétation resta longtemps majoritaire ; d'ailleurs en 1830, le zoologiste allemand Johann Georg Wagler suggéra que ces animaux devaient se servir de leurs membres avant comme de nageoires, à la manière des manchots. C'est aussi à cette époque que les ptérosaures sont interprétés comme des « reptiles volants » (Flugsaurier)[8]. Georges Cuvier suggéra dès 1801 que ces animaux pouvaient être volants[9], même s'ils ne pouvaient pas être les ancêtres des oiseaux. Les 29 premiers fossiles sortirent tous du même dépôt ; ce n'est qu'en 1828 qu'un nouveau fossile appelé Dimorphodon et décrit par Mary Anning fut découvert à Lyme Regis, au Royaume-Uni.
Edwin Tully Newton démontre en 1888 que la taille des cerveaux de ptérosaures était très semblable à celle des oiseaux, grâce à un crâne incomplet de Scaphognathus[10]. On suppose dès lors que leur capacité cognitive est comparable. Déjà, en 1901, dans le populaire Dragons of the Air, Harry Govier Seeley montra la convergence évolutive entre ces animaux et des oiseaux, en soulignant notamment que les ptérosaures et les oiseaux possédaient un système ossature-poumons complexe comparable. En effet les os creux évoquent un système de sacs aériens semblable à celui des oiseaux, cette observation sera confirmée plus tard[11]. Il émet les premiers doutes quant au caractère « reptilien » des ptérosaures, en soulignant que ces animaux disposaient vraisemblablement d'un cœur cloisonné à la manière des oiseaux et des mammifères : il existe une grande variété d'anatomies cardiaques et aucun reptile actuel ne possède cette caractéristique. Il a cependant, durant longtemps, aussi affirmé que les ptérosaures étaient les ancêtres des oiseaux, ce que les connaissances actuelles démentent.
Au fil des découvertes des fossiles, des hypothèses naissent ou se confirment ; ainsi, la présence de poils suggère l'homéothermie qui, avec certaines caractéristiques du crâne, suggère le vol actif. Au moins 60 genres de ptérosaures ont été trouvés à ce jour. Cette diversité suggère des capacités d'adaptation, comme les os des pattes recourbées de Nemicolopterus qui leur permettaient sûrement de se poser sur des branches d'arbres.
Le premier œuf fossile de ptérosaure est découvert en 2004 dans les carrières de Liaoning, en Chine[12] ; il est daté à 121 Ma. Deux autres œufs sont découverts la même année, l'un en Chine[13] et l'autre en Argentine[14]. En 2017, de nouvelles fouilles en Chine dégagent un gisement de plusieurs centaines d’œufs d'Hamipterus tianshanensis, avec des embryons en bon état et à divers stades de leur développement[15],[16].
Principales caractéristiques

À l'examen de ces fossiles surprenants, plusieurs questions se sont posées : comment ces animaux pouvaient-ils voler ? Leur vol était-il actif ? Au sol étaient-ils quadrupèdes ou bipèdes ? Étaient-ils endothermes ? Comment des espèces aussi grandes d'animaux volants ont-elles pu voir le jour ?
Les caractéristiques partagées avec les autres archosauriens se situent principalement au niveau de leur crâne diapside, ils ont un troisième « trou » devant l'œil, appelé fenêtre préorbitale, et le foramen pariétal est réduit. En outre le cinquième doigt est de taille réduite. Ils partagent également avec les ornithodires le même type de chevilles.
L'ordre des ptérosaures est divisé en deux sous-ordres (ou super-familles suivant les paléontologues) : les ptérodactyloïdés (absence partielle ou totale de queue et crâne imposant, représentés entre autres par le ptérodactyle ou encore le Quetzalcoatlus qui mesurait jusqu'à 12 m d'envergure) et les rhamphorhynchoïdés (plus archaïques, ils possédaient une queue pour la stabilité et un crâne plus petit, représentés entre autres par Rhamphorhynchus ou encore le célèbre Dimorphodon du film Jurassic World).
On soupçonne les ptérosaures d'avoir été endothermes en raison de leur pelage[17] de phanères appelés pycnofibres, depuis une publication sur les tissus mous du genre Jeholopterus coécrite par le paléontologue Alexander W. A. Kellner en 2009[18].
Certaines espèces du Crétacé, comme Quetzalcoatlus et Hatzegopteryx, ont atteint 12 mètres d'envergure, la taille d'un petit avion.
Ils étaient ovipares et quadrupèdes au sol comme les chauves-souris (résultats notamment des fouilles de la plage aux ptérosaures à Crayssac depuis les années 1990)[19]. Ils venaient sans doute écumer le littoral à la recherche de nourriture ou pour s'y reposer, à l'instar des oiseaux marins actuels[20].
L'alimentation des ptérosaures était variée : ils pouvaient être piscivores, insectivores, filtreurs (à la manière des flamants roses comme Pterodaustro), mangeurs de coquillages (avec des dents plates pour briser la coquille chez Dsungaripterus).
Le squelette
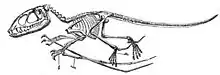

Leur squelette montre de nombreuses particularités uniques, bien sûr très adaptées pour le vol, aussi la convergence évolutive entre ce groupe et celui des oiseaux est certaine sur de nombreux points. Comme les oiseaux et les dinosaures avant eux, leur squelette est pneumatisé, c'est-à-dire qu'il pouvait contenir de l'air qui leur servait à la respiration. Comme ces derniers également, l'intérieur des os est renforcé par une structure en nid d'abeille. À certains égards, notamment en ce qui concerne la solidité et l'économie de poids, les adaptations au vol vont au-delà de celles observées pour les oiseaux. Leurs os semblent en effet étonnamment fins et creux, par conséquent légers et cependant plus solides que ceux des oiseaux grâce à de fins piliers[21]. Seuls les Pterodactyloidea disposent d'un squelette un peu différent, plus adapté à une marche verticale. Le corps est considéré comme la partie la plus parlante pour connaître le mode de vie de ces animaux.
Le nombre de vertèbres du cou varie de sept à neuf. On compte, pour les espèces découvertes, entre 10 et 18 vertèbres, sans compter les vertèbres sacrées. Chez les grands Pterodactyloidea, les vertèbres 3 à 7 ont fusionné en une structure appelée « Notarium », qui rigidifie grandement la colonne vertébrale vis-à-vis des mouvements de torsion qui devaient accompagner les battements d'ailes. Ils disposent de Gastralia. La cavité abdominale est courte, il y a six ou huit côtes abdominales, six à sept côtes sternales à peine esquissées. Les clavicules sont fusionnées avec le sternum ou n'existent pas. La scapula est importante, le coracoïde est bien marqué, et les deux sont fusionnés au sein d'un ensemble appelé scapulocoracoïde, ce qui confère une certaine rigidité à l'animal.
Les Ptérosaures disposent d'un os supplémentaire, le ptéroïde, sur l'avant-bras. Il devait avoir une utilité dans la tension du propatagium. Un autre os qu'ils sont des seuls à posséder est le pré-pubis, situé à l'avant du pubis.
Les os de la ceinture pelvienne semblent beaucoup moins solides et leur organisation moins sophistiquée que ceux de la ceinture scapulaire. De trois à dix vertèbres sacrées sont fusionnées avec les Ilions. Chez les premiers Rhamphorhynchoidea, cette structure est relativement lâche, elle est plus soudée chez les Pterodactyloidea.
Leur crâne est très grand — parfois plus long que leur corps, cou compris. [réf. souhaitée] La plupart des espèces ont une structure crânienne à peu près identique. Les évolutions et donc les différences s'observent principalement dans la partie antérieure de celui-ci, les prémaxillaires, maxillaires supérieurs et les os du front. Les fenêtres temporales ne se sont pas fermées. Les orbites des yeux sont particulièrement grands. Ils disposent d'anneaux sclérotiques. La vision est légèrement stéréoscopique, permettant de voir les reliefs.
On suppose que les conduits du système d'olfaction passaient soit par la fenêtre préorbitale, sous les yeux, soit par un trou au fond de la gueule.
Les Pterodactyloidea disposent d'une spectaculaire crête cartilagineuse, de formes diverses. L'utilité de cet appendice n'est pas connue.
Au cours de leur évolution, plusieurs groupes de Ptérosaures, plus particulièrement des Pterodactyloidea géants, finissent totalement édentés. Cette disparition a eu lieu de manière convergente chez les Pteranodontidae et les Azdharchoidea.
Les ailes et le vol
Il est aisé, par rapport aux espèces contemporaines, d'imaginer que les petites espèces de ptérosaures devaient voler. Les plus grandes devaient simplement pouvoir planer. En effet, le vol actif demande beaucoup d'énergie. De ce fait, on les suppose homéothermes. Il n'y a pas de preuve directe de cette hypothèse, mais d'autres archosauriens comme les dinosaures sont aujourd'hui considérés comme très probablement à sang chaud ; le pelage des ptérosaures est surtout essentiellement interprété comme une façon de conserver la chaleur produite par le corps.

Les ailes étaient formées par un patagium, une membrane de peau semblable à celle des chauves-souris formée le long des os des membres avant. Ce ne sont pas les mêmes os des « mains » que ceux mis en œuvre dans les ailes des oiseaux et des chauves-souris. De ce fait, on peut exclure une filiation directe entre ces groupes d'animaux. Le patagium n'était pas soutenu par les quatre derniers doigts comme pour les chauves-souris, ni le second comme les oiseaux, mais par un allongement du quatrième doigt (le cinquième ayant disparu). On ignore si la membrane se rattachait au corps de l'animal par le bassin ou le genou. Elle devait être renforcée de fibres.
Mode de déplacement au sol
Le mode de déplacement au sol et donc la manière dont les ptérosaures décollent sont depuis longtemps source de controverses. Depuis que ces animaux sont considérés comme volants, ils étaient considérés comme quadrupèdes[22], à quelques nuances près pour les Pterodactyloidea. Dans les années 1980, les paléontologues Kevin Padian et Jeremy Raynor suggèrent que même les petits ptérosaures comme Dimorphodon étaient principalement bipèdes[23], à la manière des oiseaux du genre Geococcyx. Plusieurs empreintes fossiles découvertes depuis montrent cependant distinctement quatre doigts pour les membres arrière et trois doigts pour les membres avant, prouvant que les ptérosaures posaient bien leurs quatre membres sur le sol pour se déplacer[24],[25]. Contrairement à la plupart des vertébrés, les ptérosaures sont plantigrades. Les membres de certaines familles de ptérosaures comme les Azhdarchidae et les Ornithocheiridae sont même suffisamment allongés pour suggérer qu'ils puissent être d'excellents marcheurs sans pour autant être des coureurs rapides du fait de leurs courts membres arrière[26].
La taille relative des membres au contact avec le sol, en comparaison avec les animaux modernes tels que les oiseaux, permet d'imaginer le mode de vie probable de chacune de ces espèces. Les azhdarchidés ont des pattes relativement petites en comparaison de leur taille, ces pattes mesurent environ 25 % à 30 % de la longueur du tibia, cela suggère que ces animaux se déplaçaient mieux sur terrain sec et dur que sur terrain souple ou humide. Les Ptéranodons, quant à eux, ont des pattes légèrement plus grandes, elles mesurent 47 % de la longueur du tibia, tandis que les ptérosaures filtreurs, comme les Ctenochasmatoidae, avaient de très grandes pattes. Celles-ci mesuraient 69 % de la longueur du tibia pour les Pterodactylus et même 84 % pour Pterodaustro. Ces pattes étaient adaptées à la marche dans la boue et les sols mous semblables à celle des échassiers modernes[27].
La morphologie de leur bassin suggère aussi qu'ils étaient capables de se dresser sur leurs pattes arrière comme peuvent le faire certains lézards actuels, cette capacité pourrait être à l'origine d'un bond permettant de décoller[28].
Reproduction
La forme étroite du bassin des fossiles découverts implique que les ptérosaures étaient ovipares, comme la plupart des reptiles. L'œuf découvert en 2004 a été écrasé et la surface de celui-ci présente des signes de fissuration comme cela se produirait sur une coquille à membrane souple chez les reptiles actuels[12]. La composition chimique du fossile tend à prouver que ces œufs devaient être enterrés comme le sont aujourd'hui ceux des tortues et des crocodiliens. On comprend dès lors que ces animaux aient pu se retrouver défavorisés face aux oiseaux[29]. En effet ces derniers utilisent la chaleur de leur corps pour développer leurs embryons ainsi ces derniers sont moins soumis aux aléas climatiques. En outre les oiseaux disposent d'un potentiel de lieux de ponte plus important. Nous ne savons pas si les ptérosaures nourrissaient leurs petits ou si, comme pour certains reptiles modernes, les petits, précoces, devaient tenir sur les réserves de leur œuf jusqu'à l'obtention de leur autonomie alimentaire. Enfin, nous ne connaissons pas la durée d'incubation, elle peut énormément varier, allant jusqu'à l'ovoviviparité.
Le patagium de l'embryon découvert en 2004 est bien développé[alpha 2], cela suggère que les ptérosaures devaient pouvoir voler peu de temps après leur éclosion[30]. Le fossile d'un juvénile découvert dans les limons de Solnhofen corrobore cette conclusion[31]. Plusieurs spécimens juvéniles ont aussi été découverts sur le site du désert d'Atacama au Chili. Les nombreux embryons découverts en 2017 confirment l'absence de calcification de l'enveloppe des œufs (contrairement aux crocodiles, aux dinosaures et aux oiseaux), œufs qui ressemblent à ceux des lézards actuels. Comme pour ceux-ci, il est probable que les œufs des ptérosaures étaient enfouis dans le sol pour éviter leur dessication, et qu'ils n'étaient pas couvés[16],[32].
Régime alimentaire

Une centaine d'espèces de ptérosaures sont actuellement connues. Il existe une grande diversité de modes d'alimentation au sein de ce groupe. Leur dentition (du moins pour ceux qui ont des dents) fournit de nombreuses informations sur le régime alimentaire des ptérosaures. Les caractéristiques les plus intéressantes sont le nombre de dents, leur taille, la courbure ou encore leur espacement. La structure des dents des ptérosaures est simple : une racine surmontée d'une couronne en cône recouverte d'émail. Il n'existe pas de bords coupants ni de pointes supplémentaires sauf en de rares exceptions. Cette dentition simple ne leur permettait probablement pas de mâcher, les ptérosaures devaient donc avaler la nourriture directement, comme les oiseaux actuels[33].
- Des dents écartées, longues à l'avant et réduites à l'arrière indiquent une piscivorie, à l'exemple des Rhamphorhynchoidea.
- Des dents courtes et pointues trahissent l'insectivorie, comme chez Anurognathus.
- Les nombreuses dents serrées, pointues et en crochet de Coloborhynchus devaient servir à la capture de proies importantes.
- Les longues dents très fines de Pterodaustro indiquent un mode alimentaire fondé sur le filtrage, à l'image des canards actuels.
- Les grosses dents bulbeuses de Dsungaripterus pouvaient servir à écraser des coquilles.
- Les ptérosaures édentés à museau court et puissant, comme les Tapejaridae, auraient pu se nourrir de graines.
Dénomination, taxonomie et histoire évolutive
Dénomination et définition
En 1809, Cuvier proposa comme nom Ptérodactyle pour désigner un fossile découvert en Allemagne, une construction à partir du grec ancien pouvant signifier « doigt ailé ». Ce nom allait devenir celui d'un genre particulier, les Pterodactylus. Ce terme de ptérodactyle est quelquefois utilisé comme synonyme de Ptérosaure. Le nom de Pterosauria a été proposé par Johann Jakob Kaup en 1834 et ce terme a rapidement supplanté celui d’Ornithosauria (ou « oiseau-reptile » proposé par Charles-Lucien Bonaparte en 1838. Ptérosaure, construit à partir du grec ancien πτερόσαυρος, [pterosauros], peut se traduire par « reptile ailé ».
Le taxon des Pterosauromorpha regroupe les espèces à la morphologie de ptérosaure, ce taxon regroupe donc les ptérosaures et certains de leurs ancêtres, ainsi qu'un certain nombre d'espèces non découvertes qui auraient pu descendre de ces derniers. Le paléontologue Kevin Padian définissait les Pterosauromorpha par synapomorphie avec Pterodactylus antiquus, c'est-à-dire comme ayant des os du quatrième doigt de la main très développés sur laquelle s'appuie une membrane formant une aile. Ce genre de définition ne fait plus l'unanimité, l'expression « très développés » pouvant être diversement interprétée. En effet, le nombre de genres à inclure dans ce clade peut varier avec une telle définition. En 2000, un paléontologue amateur, David Peters, proposa que l'on inclue dans le clade toutes les espèces ayant pour dernier ancêtre commun, celle des genres Preondactylus (un des plus anciens genres connus) et Quetzalcoatlus (un des plus récents genres connus), deux espèces très différentes, le second ne descendant visiblement pas directement du premier. Cette proposition a été reprise par David Unwin et adaptée par le paléontologue Alexander Kellner en 2003, qui, lui, propose d'y inclure le dernier ancêtre commun des Anurognathidae, Preondactylus et Quetzalcoatlus, et tous leurs descendants. Cependant, ces définitions seront vraisemblablement modifiées avec la découverte de nouveaux fossiles, car elles se fondent sur finalement assez peu de fossiles.
Histoire évolutive

Les plus anciens ptérosaures découverts, datés du Trias supérieur, comme Eudimorphodon ou Preondactylus, disposent déjà de toutes les caractéristiques permettant le vol battu. Leurs ancêtres, à l’instar de ceux des chauves-souris, devaient être arboricoles et planeurs, morphologiquement proches des actuels écureuils volants. Mais tant que des fossiles intermédiaires clairement apparentés aux ptérosaures ne seront pas découverts dans les terrains antérieurs, trois lignées sont en lice pour le titre de précurseurs évolutifs possibles : les ornithodires tels Scleromochlus, les archosauromorphes comme Euparkeria et les prolacertiformes comme les Sharovipteryx.
Les hypothèses :
 Reconstruction de Scleromochlus par Nobu Tamura
Reconstruction de Scleromochlus par Nobu Tamura Squelette de Marasuchus, un autre ornithodire
Squelette de Marasuchus, un autre ornithodire Reconstruction de Euparkeria par Taenadoman
Reconstruction de Euparkeria par Taenadoman Reconstruction de Sharovipteryx par Dimitri Bogdanov
Reconstruction de Sharovipteryx par Dimitri Bogdanov
Une des plus anciennes théories est celle des Ornithodira, entre-temps remise en cause par de nombreux paléontologues comme Peter Wellnhofer, David Unwin, S. Christopher Bennett. Ce dernier suggère, en se fondant sur des analyses synapomorphiques, que les ancêtres des ptérosaures pourraient être des archosauromorphes de la famille des Erythrosuchidae ou des Proterochampsidae[34]. Unwin et d’autres pensent que les ptérosaures descendent des prolacertiformes comme Sharovipteryx[35]. Il apparaît en 2020, sur la base du crâne, de l'oreille interne et des os de la main, que le groupe le plus proche des ptérosaures est celui des Lagerpétidés (des ornithodires)[36],[37].
Plus globalement, la plupart des scientifiques pensent que ces animaux ont évolué à partir d'archosauriens, à partir d'autres espèces que celles qui allaient évoluer en dinosaures.
Disparus en même temps que les dinosaures, les ptérosaures ont laissé plus de points d'interrogation que de fossiles. Or, en 2009, un site chinois a livré un fossile de Darwinopterus — une forme transitionnelle dans l'évolution du ptérosaure — puis un fossile d'adulte du même genre avec un œuf, vieux de 160 millions d'années. Pour David Unwin, de l'université de Leicester, et ses collègues, cette seconde découverte vient étayer la théorie d'une différenciation sexuelle chez les ptérosaures : les femelles avaient un bassin plus large, et seuls les mâles portaient une crête. Mais, estime Kevin Padian, de l'université de Californie à Berkeley, nous ne connaissons pas assez le développement de cette espèce pour savoir si les différences morphologiques constatées sur les fossiles sont dues à l'âge ou au sexe. Des travaux plus poussés pourraient répondre à ces questions.
Cladogramme le plus couramment admis :
├─o Autres groupes de reptiles dont les Plesiosauria, Ichthyosauria, Testudines, ... └─o Archosauria ├─o │ └─o Crurotarsi (dont les crocodiliens) └─o Incertain ├─o Pterosauria └─o Dinosauria └─o Aves (les oiseaux)
Comme les ptérosaures ne semblent pas montrer d'adaptation spécifique à la vie dans les arbres, la plupart des hypothèses supposent que l'aptitude à voler des ptérosaures s'est développée d'une autre manière que chez les oiseaux. Les ancêtres proposés dans ces hypothèses ont donc de longues pattes offrant une aptitude certaine à courir. Scleromochlus ou Sharovipteryx ont de plus, une grande surface de peau qui couvre leurs pattes arrière à la queue, caractéristique rappelant le patagium. Les ptérosaures seraient dès lors issus d'une évolution de ces espèces vivant près de falaises. Cependant, en 2008, une équipe de chercheurs a avancé des indices montrant que les premiers ptérosaures pourraient être de petits insectivores, vivant dans les arbres des forêts tropicales[4].
Des différences morphologiques évidentes différencient les premiers ptérosaures connus des derniers, car ce sont plus de 150 Ma qui séparent les spécimens découverts. Les plus anciens fossiles disposaient d'une longue queue, certaines de leurs dents disposaient de plusieurs cupsides comme les autres achosauriens. Dans la classification classique, on suppose que les ancêtres des ptérosaures ont évolué en deux lignées distinctes, les Pterodactyloidea sans queue et les Rhamphorhynchoidea disposant d'une queue. Cependant il semblerait que, au fil du temps, la queue de ces animaux se serait réduite, aussi la classification classique a perdu de sa pertinence phylogénique. En effet, les fossiles avec une queue courte n'apparaissent qu'à partir de 160 Ma. Les spécimens de Pterodactyloidea sont aussi, en moyenne, beaucoup plus grands.
Les premiers ptérosaures
Liste des familles de ptérosaures du Trias
|
Le genre Eudimorphodon de la famille des Campylognathoididae a été daté du Trias, précisément il y a 217 Ma, c'est le plus ancien fossile de ptérosaure connu. L'origine des ptérosaures est donc bien plus ancienne car les trois familles ayant des représentants connus pour avoir vécu durant le Norien, à savoir Campylognathoididae, Anurognathidae et Dimorphodontidae montrent une diversification déjà considérable. Les ptérosaures se sont adaptés pour occuper dès cette époque des niches écologiques aussi bien terrestres que maritimes. Si ces espèces descendent d'espèces forestières, cela expliquerait la rareté des fossiles plus anciens, car les forêts fournissent peu de fossiles. Aussi l'évolution des premiers groupes est plus incertaine que celles des groupes de la fin du Crétacé. Quoi qu'il en soit, le groupe des ptérosaures comptait les plus grands animaux volants connus de l'époque, certains mesurant déjà environ deux mètres d'envergure. Cette taille, non négligeable, n'est cependant pas démesurée, si on se rappelle d'une part la taille (plus de 12 mètres) que le taxon allait atteindre par la suite, et sachant d'autre part que les libellules de cette époque mesuraient 60 centimètres d'envergure. On ne sait pas si les ptérosaures sont intervenus dans la disparition de ces insectes géants.
Selon David Unwin, la forme la plus primitive serait Preondactylus avec des jambes relativement longues et des ailes courtes, une forme du crâne en dôme. Les Macronychoptera leur sont semblables avec des ailes plus longues. La branche des Dimorphodontidae conserve ses puissantes pattes, même si les Caelidracones, la plus grande de ces espèces a des jambes relativement courtes. Ce groupe s'est ensuite divisé entre les Anurognathidae, qui a conservé le même type de crâne et les Lonchognatha au crâne plus long. Ce dernier groupe se serait scindé à nouveau entre les Campylognathoididae et les Breviquartossa, qui ne se distinguent que par le crâne et de subtiles caractéristiques des ailes.
Ces espèces ne sont connues que par des fragments d'os, des dents assez caractéristiques au sein de l'ordre, les squelettes complets sont rares. Le spécimen le mieux conservé en 2009 est Eudimorphodon ranzi du muséum de Bergame, on y remarque même des restes de poisson dans son estomac. La plupart des fossiles complets viennent des carrières de Zorzino en Italie ou de Preon Valley aux États-Unis.
Leurs dents sont encore bien différenciées, elles sont relativement longues, spécialement chez Rhamphorhynchus. En outre leurs os sont plus épais, moins creux. Ils disposent tous d'une longue queue, munie d'un stabilisateur.
Les Campylognathoididae continueront à se développer durant tout le Trias mais disparaissent à la fin de celui-ci, tandis que vont prospérer les deux autres groupes.
L'émergence des Pterodactyloidea
| Liste des familles de Ptérosaures du Jurassique |
Exceptée une branche de Ptérosaures, les Pterodactyloidea, aucune ne semble avoir profondément évolué au cours du Jurassique, mais le nombre de fossiles est faible, et il est difficile en 2009 de se faire une opinion précise. Ce n'est qu'après la première moitié du Jurassique que les Pterodactyloidea commencent à jouer un rôle prépondérant. Les fossiles de la carrière de Solnhofen et ceux de Qaratai (Karatau) dans les montagnes du Kazakhstan datent de cette époque. Unwin suggère que les Pterodactyloidea sont issus d'un groupe apparu lors de la scission des Breviquartossa et des Rhamphorhynchidae. Du fait de la mobilité de ces espèces, il est impossible de savoir de quelle partie du monde elles sont issues.
Quelques espèces de cette époque ont des dents striées mais pour la plupart elles sont lisses. Alors que les ptérosaures plus anciens se déplaçaient à quatre pattes, en repliant leurs ailes, les Pterodactyloidea devaient pouvoir se tenir sur leur pattes arrière. Ceci s'observe par exemple à la façon dont le crâne est fixé d'une manière plus verticale sur le cou. Les longueurs relatives des os des membres avant changent également. La queue, longue et terminée par un stabilisateur chez les autres espèces, est considérablement réduite. Le cinquième orteil des membres avant, une sorte d'ergot qui chez les autres espèces soutient le patagium, est pratiquement disparu chez eux.
Ces modifications morphologiques sont traditionnellement interprétées comme une amélioration de l'agilité en vol, une facilité à marcher, une augmentation de la vitesse de décollage pour échapper aux rapides prédateurs théropodes. Ces hypothèses sont étayées par le fait que les deux sous-groupes de Pterodactyloidea du Jurassique, les Dsungaripteroidea et les Ctenochasmatoidea, disposent de dents très spécialisées pour la pêche en eau peu profonde, permettant respectivement l'ouverture de coquilles de mollusques et la filtration de l'eau.
Contrairement à ce que l'on a longtemps pensé les autres groupes de ptérosaures comme les Rhamphorhynchidae n'ont pas disparu, il est vrai cependant que la fréquence de ces fossiles diminuent. Leur nombre a peut-être baissé en raison de la concurrence des Pterodactyloidea ou même des ancêtres des oiseaux.
Disparition
Liste des familles de Ptérosaures du Crétacé
|
Le Crétacé est marqué par le développement d'une mégafaune d'espèces géantes. Les exemples sont nombreux, chez les plésiosaures, mosasaures, ichthyosaures, et bien sûr chez les ptérosaures.
Le nombre d'espèces semble peu à peu avoir décliné jusqu'à leur disparition totale il y a 66 Ma. Les oiseaux ont occupé alors leurs niches adaptatives, du moins pour les plus petites espèces[38]. Les seuls fossiles du Maastrichtien, c'est-à-dire de la fin du Crétacé, à avoir été découverts sont ceux d'espèces géantes, plus précisément d’Azhdarchidae et de Pteranodontia. Ceci est d'autant plus surprenant qu'en général les plus grandes espèces sont les plus fragiles du fait de leur population plus réduite et leur reproduction plus lente.
La découverte en 2016 d'un ptérosaure fossile étonnamment petit (1,5 m d'envergure) dans des roches vieilles d'environ 77 Ma remet en cause l'hypothèse suivant laquelle les petits ptérosaures auraient été remplacés par de grandes espèces vers 100 Ma. Le fossile en question est celui d'un jeune adulte, de taille définitive ou presque. Les fossiles de juvéniles des grandes espèces de ptérosaures de la même période étant rares, il est possible que l'enregistrement fossile soit biaisé en défaveur des petits ptérosaures[39].
L'extinction massive de la fin du Crétacé n'a pas touché que les espèces terrestres comme les dinosaures ou les ptérosaures mais aussi la plupart des espèces marines. Il est donc facilement compréhensible que les espèces de ptérosaures liées au milieu marin se soient également éteintes. Toutefois, cela n'explique pas la disparition des azhdarchidés et des istiodactylidés, espèces non inféodées à la mer. Toutes ces disparitions de ptérosaures semblent indépendantes de l'explosion radiative aviaire.
Les plus importants gisements de fossiles de ptérosaures pour cette époque sont ceux qui se situent du Texas au Kansas, ceux de Santana dans le bassin de l'Araripe au Brésil, ceux de Xinjiang en Chine. Les deux derniers ayant la particularité, comme à Solnhofen, de contenir des fossiles avec les empreintes des tissus mous contrairement à ceux du désert nord-américain où les vestiges sont des os fragiles et dispersés. Les sites sud américains ne fournissent pas du tout les mêmes espèces que les autres, à savoir Anhanguera, Ornithocheirus, Tapejara, Thalassodromeus, Ludodactylus et Tupuxuara. En Chine on a découvert par exemple Dsungaripterus et Noripterus, des espèces à fortes mâchoires, mais aussi entre 2014 et 2017, un gisement d'Hamipterus comportant de nombreux fossiles accompagnés de plus de 200 œufs[15].
Classifications
Du fait du nombre relatif de fossiles et surtout de l'absence de forme transitionnelle, une classification basée sur la phylogénie est nécessairement hypothétique. Plusieurs ont été proposées, notamment celle de David Unwin en 2003 puis 2006[40], celle de Kellner en 2003, celle de Wellnhofer en 1991, celle de Peters en 1997.
En outre de nombreuses espèces sont découvertes tous les ans [réf. souhaitée], ce qui rend ces classifications d'autant plus instables.
La classification classique
Felix Plieninger a proposé en 1901 de séparer le groupe en deux taxons[41] :
- Les ramphoryncoïdes, qui sont les premiers ptérosaures apparus au Trias ; ils sont caractérisés par une longue queue, des dents et l'absence de crête. Dans la conception classique, ces ptérosaures ont disparu avant la fin de l'ère. Ce groupe est en fait paraphylétique et certaines de ses espèces ont évolué jusqu'à leur disparition à la fin du Jurassique, d'autres ont survécu.
- Les ptérodactyloïdes : ce groupe de ptérosaures est apparu au Jurassique et se distingue du précédent par une queue courte, l'absence de dents chez les formes les plus évoluées, une réduction du cinquième orteil et une crête sur la tête chez certaines espèces.
Classification actuelle selon BioLib (6 juin 2016)[42] :
- sous-ordre Rhamphorhynchoidea Carroll, 2000 †
- famille Dimorphodontidae Seeley, 1870 †
- famille Eudimorphodontidae Wellnhofer, 1978 †
- famille Campylognathoididae Kuhn, 1967 †
- famille Rhamphorhynchidae Seeley, 1870 †
- famille Anurognathidae Kuhn, 1937 †
- genre Comodactylus Galton, 1981 †
- genre Peteinosaurus Wild, 1978 †
- sous-ordre Pterodactyloidea Plieninger, 1901 †
- super-famille Ornithocheiroidea Seeley, 1901 †
- famille Ornithocheiridae Seeley, 1870 †
- famille Istiodactylidae Howse, Milner & Martill, 2001 †
- famille Anhangueridae Campos & Kellner, 1985 †
- famille Azhdarchidae Nessov, 1984 †
- famille Ctenochasmatidae Nopsca, 1928 †
- famille Dsungaripteridae Young, 1964 †
- famille Lonchodectidae Unwin & al., 2000 †
- famille Pterodactylidae Bonaparte, 1838 †
- famille Pterodaustriidae Bonaparte, 1971 †
- famille Tapejaridae Kellner, 1989 †
- genre Dermodactylus Marsh, 1881 †
- genre Eosipterus Ji & Ji, 1997 †
- genre Herbstosaurus Casamiquela, 1974 †
- genre Kepodactylus Harris & Carpenter, 1996 †
- genre Mesadactylus Jensen & Padian, 1989 †
- genre Mythunga Molnar & Thulborn, 2008 †
- genre Ningchengopterus Lü, 2009 †
- genre Nyctosaurus Marsh, 1876 †
- genre Quetzalcoatlus Lawson, 1975 †
- genre Wyomingopteryx †
- super-famille Ornithocheiroidea Seeley, 1901 †
- genre Changchengopterus Lü, 2009 †
- genre Darwinopterus Lü & al., 2009 †









Ptérosaures dans la culture
Le , la maison de vente aux enchères en ligne Catawiki a vendu un squelette de Ptérosaure vieux de 110 millions d’années, pour 150000 euros, dont il n’existe que quatre squelettes presque complets dans le monde[43].
Les ptérosaures dans la littérature
Un ptérodactyle est au cœur de l'intrigue d’Adèle et la Bête, bande dessinée de Jacques Tardi.
Les ptérosaures au cinéma
Au cinéma et à la télévision, les ptérosaures sont toujours représentés capturant leurs proies avec leurs pattes comme des rapaces. Alors qu'en réalité, les ptérosaures ne possédaient pas de serres.
On peut voir des ptéranodons dans le film Jurassic Park 3, ainsi que dans la série Nick Cutter et les portes du temps (saison 1, épisode 5).
On peut également voir des ptérosaures dans la série américaine Terra Nova, coproduite par Steven Spielberg (saison 1, épisode 3).
Enfin, un des kaijus les plus connus est le ptérosaure mutant géant Rodan qui est apparu dans plusieurs films, jeux vidéo, livres et bandes dessinées, sous son nom ou au coté d'autre kaijus comme Godzilla.
- Voir aussi la Catégorie:Film de ptérosaure.
Dans la musique
Les ptérosaures sont le logo du groupe d'Avant-Prog américain Birdsongs of the Mesozoic apparaissant également dans presque toutes les pochettes des albums du groupe.
Le mythe d'une survie du groupe
Comme pour les plésiosaures, des rumeurs prétendent que les ptérosaures ne seraient pas totalement éteints. En cryptozoologie, certains cryptides sont parfois présentés comme des ptérosaures (comme le kongamato d'Afrique australe ou le ropen de Nouvelle-Guinée)[réf. souhaitée]. Cependant, aucune preuve concrète ne permet de confirmer cette hypothèse.
Notes et références
Notes
- En raison des découvertes récentes en paléontologie et en phylogénétique, les expressions « reptile volant », comme celles de « reptile préhistorique », « reptile disparu », « reptile marin », ou « âge des reptiles » sont aujourd'hui l'objet de controverses de neutralité entre le point de vue de la systématique évolutionniste (D. Aubert, Classer le vivant : les perspectives de la systématique évolutionniste moderne, Ellipses 2017) et le point de vue cladistique (Guillaume Lecointre dir., Corinne Fortin, Marie-Laure Le Louarn Bonnet, Gérard Guillot, Guide critique de l'évolution, Belin 2009, (ISBN 978-2-7011-4797-0)). Elles sont donc à manier avec précaution, notamment dans le contexte de la vulgarisation scientifique.
- L'embryon aurait eu une envergure de 27 centimètres.
Références
- (en) Paul M. Barrett, Richard J. Butler, Nicholas P. Edwards et Andrew R. Millner, « Pterosaur distribution in time and space: an atlas », Zitteliana, vol. 28, , p. 61-107 (lire en ligne [PDF]).
- (TPDB, 2014)
- (en) Buffetaut, E., Laurent, Y., Le Loeuff, J. et Bilotte, M. (1997), A terminal Cretaceous giant pterosaur from the French Pyrenées, Geological Magazine, 134(4), p. 553–556.
- (en) Xiaolin Wang, Alexander W. A. Kellner, Zhonghe Zhou et Diogenes de Almeida Campos, « Discovery of a rare arboreal forest-dwelling flying reptile (Pterosauria, Pterodactyloidea) from China », Proceedings of the National Academy of Sciences, vol. 105, no 6, , p. 1983–1987 (PMID 18268340, DOI 10.1073/pnas.0707728105)
- Le Musée royal de l'Ontario, « Un nouveau moulage de squelette de ptérosaure arrive juste à temps pour les fins de semaines », sur www.rom.on.ca,
- Buffetaut, Eric, Grigorescu, Dan, et Csiki, Zoltan, « A new giant pterosaur with a robust skull from the latest Cretaceous of Romania. », Naturwissenschaften, vol. 89, no 4, , p. 180-184 (résumé)
- Collini, C. A. (1784). Sur quelques Zoolithes du Cabinet d’Histoire naturelle de S. A. S. E. Palatine & de Bavière, à Mannheim. Acta Theodoro-Palatinae Mannheim 5 Pars Physica, pp. 58–103 (1 plate).
- Wagler, J. (1830). Natürliches System der Amphibien, Munich, 1830 : 1-354.
- Cuvier, G. (1801). [Reptile volant]. In : Extrait d’un ouvrage sur les espèces de quadrupèdes dont on a trouvé les ossements dans l’intérieur de la terre. Journal de Physique, de Chimie et d’Histoire Naturelle, 52 : 253–267.
- Newton, E. T. (1888). On the skull, brain, and auditory organ of a new species of pterosaurian (Scaphognathus purdoni) from the Upper Lias near Whitby, Yorkshire. Philosophical Transactions of the Royal Society of London, B, 179 : 503-537.
- Laurent Sacco, « Les ptérosaures respiraient comme les oiseaux ! », sur Futura-Sciences,
- (en) Ji, Q., Ji, S., Cheng, Y., You, H., Lü, J., Liu, Y., et Yuan, C., Pterosaur egg with a leathery shell, vol. 432, (DOI 10.1038/432572a), p. 572
- (en) Xiaolin Wang et Zhonghe Zhou, « Pterosaur embryo from the Early Cretaceous », Nature, vol. 429, , p. 621 (DOI 10.1038/429621a).
- (en) Luis M. Chiappe, Laura Codorniú, Gerald Grellet-Tinner et David Rivarola, « Palaeobiology: Argentinian unhatched pterosaur fossil », Nature, vol. 432, , p. 571-572 (DOI 10.1038/432571a).
- Hervé Morin, « Des œufs de ptérosaures découverts en Chine révèlent la genèse d’un reptile volant », Le Monde, (lire en ligne, consulté le ).
- (en) D. Charles Deeming, « How pterosaurs bred », Science, vol. 358, no 6367, , p. 1124-1125 (DOI 10.1126/science.aao6493).
- Unwin, D. M., et Bakhurina, N. N., 1994. « Sordes pilosus and the nature of the pterosaur flight apparatus », Nature 371, 62-64.
- (en) Kellner, A. W. A., Wang, X., Tischlinger, H., Campos, D., Hone, D. W. E., et Meng, X. (2009). "The soft tissue of Jeholopterus (Pterosauria, Anurognathidae, Batrachognathinae) and the structure of the pterosaur wing membrane." Proceedings of the Royal Society B, publié en ligne avant impression le 5 août 2009 ; DOI:10.1098/rspb.2009.0846.
- (en) Jean-Michel Mazin, Jean-Paul Billon-Bruyat, Pierre Hantzpergue, Gérard Lafaurie, « Ichnological evidence for quadrupedal locomotion in pterodactyloid pterosaurs: trackways from the Late Jurassic of Crayssac (southwestern France) », Geological Society, vol. 217, no 1, , p. 283-296 (DOI 10.1144/GSL.SP.2003.217.01.17)
- René Cubaynes, Les dépôts littoraux du jurassique sur la bordure quercynoise, Université Paul Sabatier, , p. 52
- Witton et Habob, "On the Size and Flight Diversity of Giant Pterosaurs, the Use of Birds as Pterosaur Analogues and Comments on Pterosaur Flightlessness", PLOS One, 2010
- Cuvier, G. (1801). [Reptile volant]. In : Extrait d’un ouvrage sur les espèces de quadrupèdes dont on a trouvé les ossements dans l’intérieur de la terre. Journal de Physique, de Chimie et d’Histoire Naturelle, 52 : 253–267.
- (en) Padian, K., « A Functional Analysis of Flying and Walking in Pterosaurs », Paleobiology, vol. 9, no 3, , p. 218-239
- (en) Padian, K., « Pterosaur Stance and Gait and the Interpretation of Trackways », Ichnos, vol. 10, nos 2-4, 2003., p. 115-126 (DOI 10.1080/10420940390255501)
- (en) Hwang K., Huh M., Lockley M. G., Unwin D. M. et Wright J. L., « New pterosaur tracks (Pteraichnidae) from the Late Cretaceous Uhangri Formation southwestern Korea », Geological Magazine, vol. 139, no 4, 2002., p. 421-435 (DOI 10.1017/S0016756802006647)
- (en) Dyke, G. J., Nudds, R. L., et Rayner, J. M. V., « Limb disparity and wing shape in pterosaurs. », Journal of Evolutionary Biology, vol. 19, no 4, 2006., p. 1339-1342(4) (DOI 10.1111/j.1420-9101.2006.01096.x)
- (en) Witton, M. P., et Naish, D., « A Reappraisal of Azhdarchid Pterosaur Functional Morphology and Paleoecology. », PLoS ONE, vol. 3, no 5, , e2271. (DOI 10.1371/journal.pone.0002271, lire en ligne)
- (Futura-Sciences, Jean Etienne, 2009)
- (en) A statistical study of Rhamphorhynchus from the Solnhofen Limestone of Germany: Year-classes of a single large species., (résumé), p. 569-580
- (en) Wang, X., et Zhou, Z., « Pterosaur embryo from the Early Cretaceous. », Nature, , p. 429 : 621
- (en) Unwin, D. M., et Bakhurina, N. N., « Sordes pilosus and the nature of the pterosaur flight apparatus », Nature, no 371, 1994., p. 62-64
- (en) Xiaolin Wang, Alexander W. A. Kellner, Shunxing Jiang, Xin Cheng1, Qiang Wang et al., « Egg accumulation with 3D embryos provides insight into the life history of a pterosaur », Science, vol. 358, no 6367, , p. 1197-1201 (DOI 10.1126/science.aan2329, présentation en ligne).
- Guide critique de l'évolution, Guillaume Lecointre, 2009.
- (en) S. Christopher Bennett, « The phylogenetic position of the Pterosauria within the Archosauromorpha », Zoological Journal of the Linnean Society, vol. 118, no 3, , p. 261–308
- (en) David M., The Pterosaurs: From Deep Time, New York, Pi Press, , 246 p. (ISBN 978-0-13-146308-0)
- (en) Kevin Padian, « Closest relatives found for pterosaurs, the first flying vertebrates », Nature, vol. 588, , p. 445-449 (DOI 10.1038/d41586-020-03420-z).
- (en) Martín D. Ezcurra, Sterling J. Nesbitt, Mario Bronzati, Fabio Marco Dalla Vecchia, Federico L. Agnolin et al., « Enigmatic dinosaur precursors bridge the gap to the origin of Pterosauria », Nature, (DOI 10.1038/s41586-020-3011-4).
- Slack, K. E., Jones, C. M., Ando, T., Harrison, G. L., Fordyce, R. E., Arnason, U., et Penny, D., 2006 : Early Penguin Fossils, Plus Mitochondrial Genomes, Calibrate Avian Evolution. Molecular Biology and Evolution 23, 1144-1155 ; .
- (en) Elizabeth Martin-Silverstone, Mark P. Witton, Victoria M. Arbour et Philip J. Currie, « A small azhdarchoid pterosaur from the latest Cretaceous, the age of flying giants », Royal Society Open Science, (DOI 10.1098/rsos.160333, lire en ligne, consulté le ).
- (en) Unwin, David M. (2006). The Pterosaurs: From Deep Time. New York, Pi Press. p. 246. (ISBN 0-13-146308-X).
- (de) Plieninger, F.,1901, Beiträge zur Kenntnis der Flugsaurier, Palaeontographica 48, p. 65-90
- BioLib, consulté le 6 juin 2016
- Bulletin d'informations de 21 h, diffusé le 16 juillet 2013, en radio sur la Première
Voir aussi
Références taxonomiques
- (en) Référence Paleobiology Database : Pterosauria
- (en) Référence BioLib : Pterosauria Kaup, 1834 †
- (en) Référence Tree of Life Web Project : Pterosauria
- Kellner, A. W. A. (2003) : Pterosaur phylogeny and comments on the evolutionary history of the group. Pp. 105-137. — In Buffetaut, E., & Mazin, J.-M. (eds.) (2003) : Evolution and Palaeobiology of Pterosaurs. Geological Society of London, Special Publications 217, London, 1-347.
- Unwin, D. M., 2003 : On the phylogeny and evolutionary history of pterosaurs. Pp. 139-190. — In Buffetaut, E., & Mazin, J.-M. (eds.) (2003) : Evolution and Palaeobiology of Pterosaurs. Geological Society of London, Special Publications 217, London, 1-347.
- Wellnhofer, P. (1991) : The Illustrated Encyclopedia of Pterosaurs. Salamander Books, London, p. 192.
Bibliographie
- Michaël Habib, « Les maîtres du ciel mésozoïque », Pour la science, no 507, , p. 44-52
Filmographie
- Ptérosaures, les monstres du ciel • Documentaire de David Attenborough et Matthew Dyas.
Articles connexes
- Vol du ptérosaure
- Phylogénie des Pterosauria
- Vectidraco, espèce de ptérosaures découverte en 2008 sur l'île de Wight
- Wukongopteridae
- Plage aux ptérosaures
Liens externes
- Jean Etienne, « Le grand ptérosaure bondissait pour s'envoler sur place », sur Futura-Sciences,
- « Deux nouvelles espèces de ptérosaures découvertes en Chine », sur Futura-Sciences, , source ADIT [Quoi ?] Chine
- Laurent Sacco, « Les ptérosaures, ces géants des airs, semblaient atterrir en douceur »,
- Ptérosauria
 Portail de la paléontologie
Portail de la paléontologie  Portail de l’herpétologie
Portail de l’herpétologie