Thalassodromeus
Thalassodromeus sethi

dont toute la partie post-crânienne est hypothétique
au Musée national de la nature et des sciences de Tokyo.
| Règne | Animalia |
|---|---|
| Classe | Archosauria |
| Ordre | † Pterosauria |
| Sous-ordre | † Pterodactyloidea |
| Super-famille | † Azhdarchoidea |
| Famille | † Thalassodromidae |
Thalassodromeus est un genre éteint de ptérosaures qui vivait dans ce qui est aujourd'hui le Brésil au cours du Crétacé inférieur, à l'Albien inférieur, il y a environ 110 Ma (millions d'années).
Thalassodromeus avait l’un des plus grands crânes connus parmi les ptérosaures, environ 1,42 m de long, avec l’une des plus grandes crêtes crâniennes de tous les vertébrés. Bien que seul le crâne soit connu, on estime que l’animal avait une envergure de 4,20 à 4,50 mètres. La crête était légère. Elle s'étend de la pointe de la mâchoire supérieure à l'arrière du crâne, se terminant par une entaille unique en forme de « V ». Les mâchoires étaient dépourvues de dents mais possédaient des bords supérieur et inférieur tranchants. Son crâne avait une grande fenêtre nasoantorbita (ouverture qui combine la fenestra antorbital devant l'œil avec la narine osseuse), et une partie de son palais était concave. La mâchoire inférieure ressemblait à une lame et parait légèrement tournée vers le haut.
Le crâne original, découvert en 1983 dans le bassin d'Araripe, au nord-est du Brésil, a été recueilli en plusieurs morceaux. En 2002, les paléontologues Alexander Kellner et Diógenes de Almeida Campos (d) ont nommé le spécimen d'holotype de Thalassodromeus sethi, seule espèce connue du genre.
Étymologie
Le nom générique signifie « coureur de la mer » (en référence à son mode d'alimentation supposé), et le nom spécifique fait référence au dieu égyptien Seth, en raison de sa crête rappelant soi-disant la couronne de Seth. D'autres érudits ont fait remarquer que la crête ressemblait plutôt à la couronne d'Amon et que Seth ne portait pas une telle couronne.
Un bout de mâchoire a été attribué à T. sethi en 2005, transféré au nouveau genre Banguela en 2014, puis attribué à Thalassodromeus en tant qu'espèce T. oberlii en 2018. Une autre espèce (T. sebesensis) a été nommée en 2015 sur la base d'un fragment de crête supposé, mais il a été démontré par la suite que ces os faisaient partie d'une carapace de tortue. Le plus proche parent de Thalassodromeus était Tupuxuara ; les deux sont regroupés dans un clade qui a été placé dans les Tapejaridae (sous-famille des Thalassodrominae) ou dans Neoazhdarchia (comme famille des Thalassodromidae).
Plusieurs théories ont été suggérées pour expliquer la fonction de la crête de Thalassodromeus, y compris la thermorégulation et l'affichage, mais celle-ci avait probablement plus d'une fonction. Les crêtes des Thalassodromidae semblent s'être développées tardivement (probablement en corrélation avec la maturité sexuelle) et pourraient avoir été sexuellement dimorphes (différentes selon le sexe). Comme le nom du genre l'indique, il a été proposé à l'origine que Thalassodromeus se soit nourri comme un oiseau écumeur moderne, en rasant la surface de l'eau et en plongeant sa mâchoire inférieure pour attraper une proie. Cette idée a ensuite été critiquée pour manque de preuves ; Thalassodromeus s'est depuis avéré avoir une forte musculature de la mâchoire et aurait pu tuer et manger des proies relativement grandes sur le sol. Les proportions d'espèces apparentées dans les membres indiquent qu'il s'est peut-être adapté pour voler dans les environnements intérieurs et qu'il aurait été efficace pour se déplacer sur le sol. Thalassodromeus est connu depuis la Formation de Romualdo, où il a coexisté avec de nombreux autres types de ptérosaures, dinosaures et autres animaux.
Description
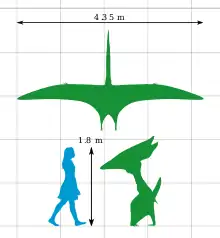
L'holotype (et le seul crâne connu) de Thalassodromeus sethi est constituée d'un des plus grands crânes de ptérosaure jamais découvert. On estime le crâne entier à 1,42 mètre de long ; les os ont été fusionnés, indiquant l'âge adulte. Sur la base de ptérosaures apparentés, l'envergure de ses ailes était de 4,2 à 4,5 m, faisant de Thalassodromeus le plus grand membre connu de son clade, Thalassodromidae. De proportions similaires, son crâne était plus lourd que celui de son parent Tupuxuara[1],[2],[3] Bien que le squelette post-crânien de Thalassodromeus soit inconnu, les proches avaient des vertèbres cervicales exceptionnellement courtes et bloceuses, avec des membres antérieurs et postérieurs bien développés, de longueur presque égale (à l'exclusion du long doigt d'aile). Les membres postérieurs représentaient 80% de la longueur de la patte antérieure, un ratio unique parmi les ptérodactyloïdes (ptérosaures à queue courte). [3] En tant que ptérosaure (ordre éteint des reptiles volants), Thalassodromeus était recouvert de pycnofibres ressemblant à des cheveux et avait de grandes membranes d'aile (prolongées par le doigt de l'aile). [3]
Le crâne de T. Sethi avait un profil fuselé, en particulier à partir de l'extrémité du museau à l'arête avant de la nasoantorbital fenestra (ouverture qui combine le fenestra antorbital en face de l'oeil avec la narine osseuse). La caractéristique la plus remarquable du crâne était la grande crête, qui longeait l'extrémité supérieure du museau et dépassait l'occiput à l'arrière du crâne, doublant presque la longueur et la hauteur du crâne. À l'exception du ptérosaure Tupandactylus imperator (dont la crête était principalement constituée de tissus mous), T. sethi présentait la crête crânienne proportionnellement la plus grande de tous les vertébrés connus (75% de la surface latérale du crâne). La crête était principalement formée par les prémaxillées (os du museau le plus en avant), les os frontaux, les os pariétaux et une partie de l'os supra-occipital. Les prémaxillaires formaient la plus grande partie de la crête, s'étendant jusqu'à son dos, et touchaient la partie frontopariétale de la crête par une suture droite (caractéristique distincte de cette espèce). La crête variait entre 1 et 10,5 mm d'épaisseur ; il s'épaississait au contact des prémaxillaires et de la partie frontopariétale et devenait progressivement plus mince vers le haut et vers l'arrière (sauf pour la partie inférieure derrière l'occiput, où il avait une base épaisse)[1],[2]. [3]

Malgré sa taille, la crête était légère et essentiellement creuse ; certaines zones indiquent des signes de pneumatisation du squelette et un système trabéculaire bien développé unissant les os. La surface de la crête avait un système de canaux de taille et d'épaisseur variables, probablement les empreintes de vaisseaux sanguins étendus[1]. [3] Une petite ouverture de 46 mm était présente au-dessus de l’ orbite (orbite), perçant la partie basale de la crête; une telle caractéristique est inconnue chez les autres ptérosaures et ne semble pas être due à des dommages. Les marges de l'ouverture sont lisses et la bordure intérieure est fenêtrée le reliant à la structure interne de la crête[2],[4]. Le dos de la crête se terminait par une encoche en forme de V, caractéristique unique de cette espèce. [3][1] Bien que d'autres parties de la crête présentent des ruptures en forme de V, la forme en V à l'extrémité ne semble pas être due à une rupture. on peut y voir les marges de l'os, toujours encapsulées dans une matrice. La crête avait probablement une couverture kératinique (cornée) et aurait pu être étendue par des tissus mous dans certaines zones, mais l'étendue de celle-ci est inconnue[1],[5].
La mâchoire supérieure de T. sethi était principalement composée de prémaxillées et de maxillaires ; la suture qui formait la limite entre ces os n'est pas visible. Comme dans tous les membres de son clade, les mâchoires étaient édentées. Le rostrum (museau) était de 650 millimètres de longueur, de la pointe du prémaxillaire à l'articulation où l'os quadratique du crâne est relié à l'os articulaire de la mâchoire inférieure. L'avant des prémaxillaires avait des bords supérieurs et inférieurs nets, uniques à cette espèce. Comme dans les genres apparentés, la fenêtre nasoantorbitale était relativement grande;elle faisait 650 mm de long et 200 mm de haut, ce qui correspond à 71% de la longueur du crâne (sans la crête). L'os lacrymal, qui séparait l'orbite de la nasoantorbital fenestra, était allongé verticalement et plus haut que la surface supérieure de l'orbite (contrairement à la condition observée chez les ptérodactyloïdes avec des fénestras nasoantorbital plus petits). L'orbite était mince et comprimée d'avant en arrière par rapport à Tupuxuara et aux tapejaridés, mais ressemblait à certaines d'entre elles en ce qu'elle faisait plus de la moitié de la hauteur de la fente nasoantorbitale. L'orbite était positionnée plus bas que le bord supérieur de la fente nasoantorbitale et donc très basse sur le crâne. Bien que les os bordant la fenêtre temporale inférieure (ouverture derrière l’orbite) soient incomplets, il semble qu’ils aient été allongés et fendus (comme dans Tupuxuara et Tapejara)[2]. [3]

La région palatine à la pointe du museau de T. sethi était une arête tranchante, semblable à la quille observée à la surface supérieure de la symphyse mandibulaire où les deux moitiés de la mâchoire inférieure étaient reliées. De petits foramina (ouvertures) en forme de fente sur les bords latéraux inférieurs de la crête indiquent que celle-ci avait dans la vie une couverture cornée semblable à celle de Tupandactylus. Le bord inférieur de la zone était légèrement incurvé, ce qui a probablement créé un petit espace lorsque les mâchoires ont été fermées. Plus en arrière, immédiatement en face de la fente nasoantorbitale, la crête palatine est devenue une quille forte, arrondie et convexe. Cette convexité s’insère parfaitement dans l’étagère symphysaire située à l’extrémité avant de la mâchoire inférieure et se serait étroitement liée lorsque les mâchoires étaient fermées. La crête palatine aboutissait à une zone fortement concave unique à cette espèce. Les fenêtres postpalatins (ouvertures derrière l'os palatin) étaient ovales et très petites, différentes de celles d'espèces apparentées. L'ectoptérygoïde (os sur le côté du palais) avait de grands côtés en forme de plaque et était bien développé par rapport aux espèces apparentées. L'os supra-occipital, qui constituait la base la plus reculée de la crête crânienne, portait des cicatrices musculaires à son extrémité supérieure (correspondant probablement à la fixation des muscles du cou)[2].
Bien que la mâchoire inférieure de T. sethi soit incomplète, sa longueur totale est estimée à 670 mm, dont 47% étaient occupés par la symphyse mandibulaire. La pointe de la mandibule est absente, mais sa face antérieure indique qu'elle pourrait avoir été légèrement tournée vers le haut, comme dans T. oberlii (la seconde espèce possible de Thalassodromeus, ou peut-être d'un autre genre - Banguela - qui n'est connu que du bout des mâchoires). La tablette symphysaire, la face supérieure de la symphyse, prolongée de 170 mm et avait une surface plane. Vus de haut, les bords latéraux de cette zone étaient hauts et formaient une marge nette. Près de l'extrémité avant de la symphyse, les bords qui formaient les marges se sont élargis vers l'avant du plateau jusqu'à ce qu'ils se rencontrent et se fondent. Les surfaces supérieure et inférieure de la mâchoire à l'avant de l'étagère étaient carénées (la quille supérieure était plus robuste et commençant avant la partie inférieure), ce qui donnait à la symphyse une forme en forme de lame. La quille inférieure est devenue plus profonde vers l'avant de la mâchoire, donnant l'impression que la mâchoire a dévié vers le bas; c'était en fait directement, à l'exception du (peut-être) bout renversé. Les fosses mandibulaires (dépressions) à l'arrière de la mâchoire supérieure étaient plus profondes et plus larges que d'habitude chez les ptérodactyloïdes, créant ainsi de grandes surfaces pour l'articulation de la mâchoire inférieure. L’espèce possible T. oberlii se distinguait de T. sethi et d’autres espèces apparentées par la surface supérieure de sa symphyse mandibulaire, étant légèrement plus courte que la surface inférieure, et se distinguait davantage de T. sethi par le bord supérieur de la symphyse beaucoup plus pointu que le inférieur. Les deux espèces partageaient des caractéristiques telles que la compression de la symphyse latéralement et de haut en bas, la quille tranchante sur le devant supérieur de la symphyse et le petit sillon longeant la surface supérieure du plateau[2].
Histoire de découverte
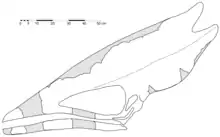
Le tout premier spécimen de Thalassodromeus connu a été trouvé en 1983 près de la ville de Santana do Cariri dans le bassin d'Araripe, dans le nord-est du Brésil. Trouvé dans les affleurements de la formation Romualdo, il a été collecté pendant plusieurs années sur une longue période. L'échantillon (catalogué comme DGM 1476-R au Museu de Ciências da Terra) a été conservé dans un calcaire nodule, et consiste en un (sont souvent aplatis os de pterosaur presque-complet, le crâne en trois dimensions fossiles de compression), manquant deux segments de le bas du crâne et de la mandibule et le devant de la mâchoire inférieure. La région jugale gauche et le ramus mandibulaire droit (la moitié de la mandibule) sont légèrement poussés vers l'intérieur. Le crâne a été signalé pour la première fois dans un livre italien de 1984, puis décrit et figuré de manière préliminaire en 1990 par les paléontologues Alexander WA Kellner et Diogenes de Almeida Campos. Bien que les morceaux de crâne aient été divisés entre des musées d'Amérique du Sud et d'Amérique du Nord, ils avaient été assemblés avant 2002[1],[6]. [3][7],[8]

En 2002, Kellner et Campos ont décrit et nommé le nouveau genre et espèce Thalassodromeus sethi, le crâne DGM 1476-R étant le spécimen holotype. Le nom générique est dérivé des mots grecs antiques θάλασσα (thálassa, "mer") et δρομεύς (dromeús, "coureur"), qui signifie "coureur de la mer" en référence au comportement supposé d'alimentation de l'animal. Le nom spécifique fait référence à l'ancien dieu égyptien Seth[1]. [3] Le spécimen n'était pas complètement préparé au moment de cette description préliminaire[2]. Les descripteurs d'origine ont choisi le nom de sethi parce que la crête du ptérosaure était supposée rappeler la couronne portée par Seth; cependant, les paléontologues André Jacques Veldmeijer, Marco Signore et Hanneke JM Meijer ont souligné en 2005 que la couronne (avec ses deux hauts plumes) était généralement portée par le dieu Amon (ou Amon-Ra), et non par Seth[9],[2].
En 2006, les paléontologues David M. Martill et Darren Naish ont suggéré que Thalassodromeus était un synonyme junior du genre apparenté Tupuxuara, nommé par Kellner et Campos en 1988 à partir de fossiles de la même formation. Selon Martill et Naish, les différences entre ces genres (y compris entre les trois espèces de Tupuxuara, T. longicristatus et T. leonardii) étaient dues à l'ontogenèse (changements durant la croissance) et à la compression des fossiles; Thalassodromeus était simplement un individu plus âgé, plus grand et mieux préservé[10]. Mais Kellner et Campos ont rejeté cette idée en 2007, en soulignant que ces espèces présentaient des caractéristiques différentes de celles de leurs crêtes. Ils ont également noté qu'un échantillon de Tupuxuara avait un crâne plus grand que Thalassodromeus, malgré l'affirmation de Martill et Naish selon laquelle ce dernier était un individu plus âgé[4]. Le point de vue de Kellner et Campos a depuis été accepté par d'autres chercheurs, notamment Martill et Naish[2],[11].

Veldmeijer et ses collègues ont attribué à T. sethi la partie antérieure d'une mandibule prélevée dans la même formation en 2005. Ils ont conclu que, bien que les deux spécimens différaient sur plusieurs points de détail, ces différences n'étaient pas suffisamment significatives pour fonder une nouvelle espèce sur la mandibule et que le nouveau spécimen comblait l'espace entre la reconstruction du crâne de T. sethi de Kellner et Campos[9]. Les paléontologues Jaime A. Headden et Herbert BN Campos ont inventé le nouveau genre Banguela oberlii, basé sur leur réinterprétation de la pointe de la mâchoire comme appartenant à un membre édenté de la famille des Dsungaripteridae en 2014. Headden et Campos interpréter la pointe de la mâchoire inférieure de T. Sethi comme tournée vers le bas; cette particularité et d'autres le distinguaient de Banguela[12]. En 208, dans leur re-description de l'holotype de T. Sethi, les paléontologues Rodrigo V. pegas, Fabiana R. Costa, et Kellner ont assigné B. oberlii à nouveau comme une seconde espèce de Thalassodromeus tout en le reconnaissant comme une espèce distincte, et ont ainsi créer T. oberlii. Pêgas et ses collègues ont également rejeté la théorie selon laquelle la mâchoire inférieure de T. sethi avait été renversée et ont réinterprété la partie la plus antérieure de la mâchoire inférieure pour s'être connectée directement à la pièce suivante (sans espace)[2].
En 2015, les paléontologues Gerald Grellet Tinner et Vlad A. Codrea ont nommé une nouvelle espèce, T. sebesensis, avec ce qu'ils ont interprété comme faisant partie d'un crête crânienne dans une concrétion trouvée près du fleuve Sebeș en Roumanie. Les auteurs ont déclaré que cela élargirait considérablement la portée temporelle et spatiale du genre Thalassodromeus, ce qui créerait un écart de 42 millions d'années entre les espèces plus anciennes d'Amérique du Sud et les plus jeunes espèces européennes[13]. Le paléontologue Gareth J. Dyke et une large équipe de collègues ont immédiatement rejeté l'identification par le ptérosaure du fossile de T. sebesensis, affirmant qu'il s'agissait d'une partie mal identifiée d'un plastron (coquille inférieure) de la tortue préhistorique Kallokibotion bajazidi (nommée en 1923). L'idée que le fragment appartenait à une tortue avait été examinée et rejetée par Grellet-Tinnera et Codrea dans leur description originale[14]. Grellet-Tinnera et Codrea ont nié l'identité de la tortue suggérée par Dyke et ses collègues, notant que ces chercheurs n'avaient pas directement examiné le fossile[15].
Classification
La classification de Thalassodromeus et de ses parents les plus proches est l’une des questions les plus controversées concernant leur groupe. [3] Kellner et Campos ont à l'origine attribué Thalassodromeus à la famille Tapejaridae, avec Tapejara, en raison de sa grande crête et de sa grande fenestra nasoantorbitale. Au sein de ce clade, ils ont constaté qu'il était différent du genre à visage court Tapejara, mais partageaient une quille en bouche avec Tupuxuara[1]. Kellner a détaillé les relations au sein des Tapejaridae en 2004 et a souligné que Thalassodromeus et Tupuxuara partageaient également une crête composée principalement d'os; la crête avait une grande partie des tissus mous chez les autres membres du groupe[5].

Martill et Naish ont considéré les Tapejaridae comme un groupe paraphylétique (non naturel) en 2006 et ont conclu que Tupuxuara (qui inclut Thalassodromeus dans leur analyse) était le taxon frère de la famille Azhdarchidae. Ce clade (Tupuxuara et Azhdarchidae) avait été baptisé Neoazhdarchia par le paléontologue David Unwin en 2003, selon un accord approuvé par Martill et Naish. Selon Martill, les caractéristiques de l'union de Neoazhdarchia comprenaient la présence d'un notarium (vertèbre soudée dans la région de l'épaule), la perte de contact entre les premier et troisième métacarpiens (os à la main) et de très longs museaux (plus de 88% de la longueur du crâne)[10],[16]. Kellner et Campos ont défendu la validité des Tapejaridae en 2007, en le divisant en deux clades: Tapejarinae et Thalassodrominae, ce dernier contenant Thalassodromeus (le genre type) et Tupuxuara. Ils ont distingué les thalassodromines par leur haute fenestre nasoantorbitale et la partie osseuse de leur crête commençant à l'avant du crâne et se prolongeant plus en arrière que chez les autres ptérosaures[4].
Les relations entre ces clades au sein du plus grand clade Azhdarchoidea demeuraient controversées et le clade contenant Thalassodromeus et Tupuxuara avait reçu différents noms de chercheurs (Thalassodrominae et Tupuxuaridae). Le paléontologue Mark P. Witton a tenté de résoudre le problème de la dénomination en 2009, soulignant que le nom "Tupuxuaridae" (utilisé pour la première fois dans la forme vernaculaire "tupuxuarids" par le paléontologue Lü Junchang et ses collègues en 2006) n'avait nom propre (bien qu'il ait été accordé un an plus tard). Witton convertit ensuite le nom de sous - famille Thalassodrominae en nom de famille Thalassodromidae et considère que le clade fait partie de Neoazhdarchia[11].
Une analyse effectuée en 2011 par le paléontologue Felipe Pinheiro et ses collègues a confirmé le regroupement des clades Tapejarinae et Thalassodrominae dans la famille Tapejaridae, rejoints par les Chaoyangopterinae[17]. Une étude réalisée en 2014 par le paléontologue Brian Andres et ses collègues a plutôt révélé que les thalassodromides se regroupaient avec les dsungaripérides, formant le clade Dsungaripteromorpha dans Neoazhdarchia (défini comme le clade le plus inclusif contenant Dsungaripterus weii mais pas Quetzalcoatlus northropi)[18].
Cladogramme basé sur l'analyse de Pinheiro et ses collègues en 2011[17] :
|
Cladogramme basé sur l'analyse de Andres et ses collègues en 2014[18] :
|
Pêgas et ses collègues ont placé les Tapejarinae et les Thalassodrominae au sein des Tapejaridae en 2018, mais ont reconnu que le sujet était toujours controversé[2].
Paléobiologie
Fonction de crête
Les fonctions possibles de la crête crânienne de Thalassodromeus ont été imaginées par Kellner et Campos en 2002. Ils ont suggéré que le réseau de vaisseaux sanguins sur sa grande surface était compatible avec l'utilisation pour la thermorégulation, ce qui avait également été suggéré pour les crêtes de certains dinosaures. Kellner et Campos pensaient que la crête était utilisée pour le refroidissement (permettant à l'animal de dissiper l'excès de chaleur métabolique par convection), tandis que le transfert de chaleur était contrôlé par le réseau de vaisseaux sanguins et en dépendait. La capacité de contrôler sa température corporelle aurait aidé Thalassodromeus pendant une activité intense (comme la chasse), et ils ont suggéré que, en vol, la chaleur aurait été dissipée plus efficacement si la crête était alignée avec le vent, alors que la tête était intentionnellement. déplacé sur les côtés. Kellner et Campos ont avancé que le blason aurait pu avoir des fonctions supplémentaires, telles que l' affichage ; aidé par la couleur, il aurait pu être utilisé dans la reconnaissance des individus et aurait également pu être une caractéristique sexuellement dimorphique (différente selon le sexe), comme cela a été proposé pour Pteranodon[1].
En 2006, Martill et Naish ont découvert que les crêtes de Tupuxuara et de leurs parents s'étaient développées avec la partie prémaxillaire des crêtes qui repoussaient sur le toit du crâne (comme l'indique la suture bien définie entre le prémaxilla et les os sous-jacents). La marge postérieure de la partie prémaxillaire de la crête de ce spécimen n'avait atteint que la marge postérieure de la fenêtre du nasoantorbital, indiquant que ce n'était pas un adulte au moment du décès. Ceci suggère que le développement de la crête a eu lieu tard dans la croissance d'un individu, était probablement lié à l'affichage sexuel et que la maturité sexuelle d'un spécimen donné pouvait être évaluée par la taille et la disposition de la crête. L'holotype T. sethi, avec sa crête prémaxillaire hypertrophiée (élargie), représenterait ainsi un individu adulte âgé (et le stade mature de Tupuxuara, selon leur interprétation)[10]. Kellner et Campos ont trouvé la discussion de Martill et Naish sur le développement de la crête crânienne intéressante, bien qu'ils aient trouvé le modèle proposé spéculatif.[4]
Les paléontologues David WE Hone, Naish et Innes C. Cuthill ont réitéré l'hypothèse de croissance de Martill et Naish en 2012; les ptérosaures étant probablement précoces et capables de voler peu après l'éclosion, le rôle de la crête n'était pertinent qu'après la maturité (lorsque la structure était complètement développée). Ils ont estimé que l'hypothèse de la thermorégulation était une explication improbable des canaux des vaisseaux sanguins situés sur la crête, ce qu'ils ont trouvé être compatible avec l'alimentation des tissus en croissance (tels que la kératine dans les becs d'oiseaux). Hone, Naish et Cuthill ont laissé entendre que les membranes des ailes et le système de sac gonflable auraient été plus efficaces pour contrôler la chaleur qu’une crête, et que le vent et l’eau auraient également aidé à refroidir les ptérosaures dans des environnements maritimes à haute température[19]. En 2013, Witton a convenu que les crêtes de thalassodromides adultes beaucoup plus grandes indiquaient qu'elles étaient plus importantes pour les activités comportementales que pour la physiologie. Il a trouvé l'idée que l'utilisation des crêtes pour la thermorégulation était problématique, car elles ne poussaient pas régulièrement avec la taille du corps. ils ont connu une croissance rapide chez les quasi-adultes, plus rapidement que prévu pour la croissance d'une structure thermorégulatrice. Selon Witton, les grandes membranes d'aile hautement vasculaires des ptérosaures fourniraient la surface nécessaire à la thermorégulation, ce qui signifie que les crêtes n'étaient pas nécessaires à cette fonction. Il a conclu que les modèles de vaisseaux sanguins de la crête ne différaient pas beaucoup de ceux observés sur les os sous le bec des oiseaux, qui sont utilisés pour transporter les nutriments vers les os et les tissus mous plutôt que pour la thermorégulation. Witton a noté que, bien que les becs d'oiseaux perdent rapidement de la chaleur, ce n'est pas pour cela qu'ils ont été développés; les crêtes de ptérosaures pourraient également avoir eu un effet sur la thermorégulation, sans que cela soit leur fonction première. [3]

Pêgas et ses collègues ont noté que le dimorphisme sexuel de la taille et de la forme de la crête avait été proposé pour certains ptérosaures; la forme de crête observée dans l'holotype T. sethi peut être en corrélation avec un sexe et peut avoir été le résultat d'une sélection sexuelle. Ils ont suggéré que les deux sexes auraient pu avoir des crêtes similaires en raison de la sélection sexuelle mutuelle, mais l'interprétation des caractéristiques exagérées était difficile en raison de la petite taille de l'échantillon ; il faudrait trouver plus de spécimens de T. sethi pour évaluer ces théories. Ils ne pensaient pas que la thermorégulation était en corrélation avec la croissance de la crête par rapport à la taille du corps, car le bec des toucans (le plus grand de tous les oiseaux modernes) devenait de plus en plus démesuré par rapport à la taille du corps et fonctionnait comme une structure de thermorégulation, tout en facilitant le comportement. Pêgas et ses collègues ont trouvé que la structure vasculaire du bec toucan était comparable à celle de la crête de T. sethi, concluant que la crête avait également de multiples fonctions[2].
Régime alimentaire

Kellner et Campos ont trouvé à l'origine des mâchoires de Thalassodromeus similaires à celles des écumeurs modernes - trois espèces d'oiseaux du genre Rhynchops - avec leurs mâchoires comprimées latéralement, leur bec en forme de lame et leur mâchoire inférieure saillante (ressemblant à des ciseaux vu de côté). Ils ont fait valoir que Thalassodromeus se serait nourri de la même manière, comme l'indique le nom du genre; Les écumeurs parcourent la surface de l'eau et plongent leur mâchoire inférieure pour attraper des poissons et des crustacés. Kellner et Campos ont répertorié les caractéristiques supplémentaires du crâne des écumeurs, qui sont des adaptations pour l'alimentation réduite, notamment des os palatins élargis, une caractéristique également partagée avec Thalassodromeus. Contrairement aux écumeurs moderrnes et aux autres ptérosaures, les os palatins de Thalassodromeus étaient concaves, ce qui, selon les auteurs, aurait pu l’aider à stocker momentanément les aliments. Comme les skimmers, Thalassodromeus semble également avoir des muscles puissants au niveau du cou, des muscles de la mâchoire larges et une extrémité de la mâchoire supérieure bien irriguée par le sang (caractéristiques que Kellner et Campos interprétaient comme des adaptations permettant d'écrémer). Ils ont conclu que le bec en forme de ciseau et la fine crête rendaient presque impossible d'autres modes de capture de proies, tels que plonger dans l'eau et s'y plonger, impossible. En admettant la difficulté de reconstruire la méthode de pêche ' ils l' ont envisagé avec un cou moins mobiles que les skimmers; avec la crête empêchant sa tête de la submerger, il glissait et battait seulement des ailes de temps en temps. Ils ont découvert que le ptérosaure ayant la mâchoire la plus semblable à celle de Thalassodromeus était le petit Rhamphorhynchus, bien qu'ils pensaient qu'il aurait eu une capacité d'écrémage limitée[1],[20].
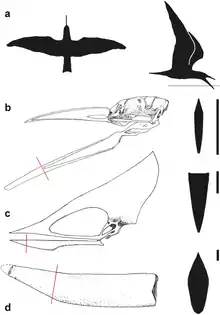
En 2004, le paléontologue Sankar Chatterjee et l’ingénieur R. Jack Templin ont déclaré que des ptérosaures plus petits auraient peut-être pu survivre. Ils doutaient que cela soit possible pour les plus grands, en raison de leur moindre manœuvrabilité et de leur capacité de vol tout en résistant à l'eau. Chatterjee et Templin ont noté que les skimmers avaient un bec plus haut que les ptérosaures comme Thalassodromeus, pour diriger l'eau de la mâchoire tout en rasant[21]. En 2007, le biophysicien Stuart Humphries et ses collègues se demandaient si des ptérosaures auraient normalement été nourris par écrémage et ont déclaré que ces conclusions reposaient sur des comparaisons anatomiques plutôt que sur des données biomécaniques. La traînée ressentie par les becs d'oiseaux et les mâchoires de ptérosaures a été testée hydrodynamiquement et aérodynamiquement en créant des modèles de becs du skimmer noir Thalassodromeus et du Tupuxuara (probablement) non rasant et en les tractant le long d'un bac rempli d'eau à différentes vitesses. Les chercheurs ont découvert que l'écumage coûtait plus cher à l'énergie pour les écumeurs que ce que l'on pensait auparavant et qu'il aurait été impossible pour un ptérosaure pesant plus de 1 kilogramme raison de la puissance métabolique requise. Ils ont découvert que même les ptérosaures plus petits, comme Rhamphorhynchus, n'étaient pas adaptés à l'écumage[22],[23]. Le gréement en aluminium du modèle Thalassodromeus a été détruit au cours de l'expérience en raison des forces élevées et instables qu'il exerce lors de l'écumage à grande vitesse, ce qui jette un doute supplémentaire sur cette méthode d'alimentation. [3] Les auteurs ont utilisé la pointe de la mâchoire de T. oberlii pour modéliser les performances de Thalassodromeus, puisqu'elles avaient été attribuées à T. sethi à l'époque[2],[12].
Unwin et Martill ont suggéré en 2007 que les thalassodromides auraient pu se nourrir de la même manière que les cigognes, comme cela avait été suggéré pour les azhdarchidés[24]. Witton a déclaré en 2013 que, bien que l'alimentation allégée ait été suggérée pour de nombreux groupes de ptérosaures, l'idée a été critiquée ces dernières années. les ptérosaures manquaient pratiquement de toutes les adaptations pour une alimentation réduite, ce qui rendait improbable leur alimentation de cette façon. Thalassodromeus (contrairement aux skimmers) ne possédait pas de crâne particulièrement large ou robuste, ni de sites de fixation des muscles de la mâchoire particulièrement grands, et sa mandibule était comparativement courte et bombée. Witton a convenu avec Unwin et Martill que les thalassodromides, avec leurs proportions égales de membres et leurs mâchoires allongées, étaient adaptés à l'itinérance terrestre et à une alimentation opportuniste; leurs cous plus courts et plus flexibles indiquaient une manière de se nourrir différente de celle des azhdarchids, qui avaient des cous plus longs et plus raides. Il a suggéré que les thalassodromides avaient peut-être des habitudes alimentaires plus généralisées et que les azhdarchids étaient peut-être plus restreints; Thalassodromeus était peut-être plus doué pour le traitement de proies relativement grandes et en difficulté que son parent, Tupuxuara, dont le crâne était plus léger. Witton a souligné que plus d' études de morphologie fonctionnelle devraient être fait pour éclairer le sujet et spéculé que Thalassodromeus aurait pu être un prédateur raptorial, en utilisant ses mâchoires pour maîtriser leurs proies avec de fortes morsures; sa bouche concave pourrait l'aider à avaler de grandes proies. [3]
Pêgas et Kellner ont présenté une reconstruction des muscles mandibulaires de T. sethi lors d'une conférence en 2015. Ils ont constaté que ses muscles de la mâchoire bien développés étaient différents de ceux du possible Anhanguera, alimenteur par immersion, et des azhdarchidés traqueurs terrestres, indiquant que T. sethi exerçait une force de morsure importante[25]. En 2018, Pêgas et ses collègues sont convenus que ' robustes mâchoires en forme de lame de Thalassodromeus indiquaient qu'il aurait pu les utiliser pour frapper et tuer des proies, mais estimaient qu'un travail biomécanique était nécessaire pour étayer cette idée. Ils ont découvert (contrairement à Witton) que Thalassodromeus avait une articulation de la mâchoire renforcée et des muscles de la mâchoire robustes, mais que des efforts supplémentaires étaient nécessaires pour déterminer ses habitudes alimentaires. Selon Pêgas et ses collègues, l'articulation entre T. articulaire de Sethi et les os quadrate (où la mâchoire inférieure reliée au crâne) indique un bâillement maximum de 50 degrés - semblable aux 52 degrés inférés pour Quetzalcoatlus.[2]
Locomotion

Dans un commentaire de 2002 sur la description originale de T. sethi, l’ingénieur John Michael Williams a noté que, bien que Kellner et Campos aient mentionné que la grande crête aurait pu interférer de manière aérodynamique pendant le vol, ils n’avaient pas précisé ce point et avaient comparé le ptérosaure à un oiseau un cinquième de sa taille. Il a suggéré que Thalassodromeus utilisait sa crête pour équilibrer ses mâchoires, l'attitude changeant de la tête dépendant du mode de locomotion. Williams a supposé que la crête serait gonflable avec du sang et présentait une résistance à l'air variable, ce qu'il comparait à un ventilateur portatif; cela aurait aidé l'animal à modifier l'attitude de la tête pendant le vol (et lors du contact avec de l'eau), l'empêchant de tourner sans des muscles puissants du cou. La crête aurait rendu possible de longs vols plutôt que d'interférer; Williams l'a comparée au spermaceti dans la tête du cachalot, affirmant qu'il est censé être utilisé pour modifier la flottabilité en ajustant la température. Kellner et Campos ont rejeté l'idée d'une crête gonflable, car ses os compressés ne le permettraient pas. ils n'ont pas trouvé l'analogie cachalot-baleine convaincante en ce qui concerne les animaux volants, notant que le spermaceti est plus susceptible d'être utilisé pendant une agression ou pour le sonar. Ils sont convenus que l'idée que la crête avait une fonction en vol était tentante et qu'un mouvement latéral de la tête l'aurait aidé à changer de direction, mais des études biomécaniques et mécaniques en vol de la crête devraient être menées pour déterminer l'aérodynamique de l'animal[26].
Witton a également exprimé l'espoir d'une analyse plus approfondie de la locomotion du thalassodromide. Il a noté que puisque les proportions de leurs membres étaient similaires à celles des azhdarchidés mieux étudiés, la forme de leurs ailes et leur style de vol auraient pu être similaires. Les thalassoïdes pourraient également avoir été adaptés au vol intérieur; leurs ailes étaient courtes et larges (contrairement aux ailes longues et étroites des soulèvements marins), plus maniables et moins susceptibles de s’accrocher aux obstacles. Leurs muscles de l'épaule inférieure semblent avoir été dilatés, ce qui aurait pu contribuer à des mouvements puissants (ou fréquents) de la chute des ailes et à une capacité de décollage. Bien qu’il ait peut-être dû compenser sa crête importante en vol, son développement tardif indique qu’il ne s’est pas développé principalement pour l’aérodynamique. Witton a suggéré que la similarité proportionnelle entre les membres des thalassodromides et des azhdarchides indique également que leurs capacités terrestres auraient été comparables. Leurs membres auraient été capables de longues foulées et leurs pieds courts et compacts auraient rendu ces mécanismes efficaces. Les muscles de l'épaule élargis ont peut-être permis une accélération rapide de la course et ont été adaptés au mouvement sur le sol, comme cela a été suggéré pour les azhdarchidés; Witton a averti qu'une analyse plus poussée des thalassodromides était nécessaire pour déterminer cela. [3]
Paléoécologie

Thalassodromeus est connu de la formation Romualdo, qui date de l'Albien stade du Crétacé inférieur période (il y a environ 110 millions d' années). La formation fait partie du groupe Santana et, à l'époque de la description de Thalassodromeus, était considérée comme un membre de ce qui était alors considéré comme la formation Santana. La formation de Romualdo est un Lagerstätte (dépôt sédimentaire qui conserve des fossiles en excellent état), constituée de concrétions calcaires de lagons enchâssées dans des schistes argileux et recouvrant la formation de Crato. Il est bien connu pour la conservation tridimensionnelle de fossiles dans des concrétions calcaires, notamment de nombreux fossiles de ptérosaures. On y a trouvé des fibres musculaires de ptérosaures et de dinosaures, des branchies préservant le poisson, des voies digestives et des cœurs[1],[2]. Le climat tropical de la formation correspond en grande partie au climat brésilien actuel[27]. La plupart de sa flore était xérophyte (adaptée aux environnements secs). Les plantes les plus répandues étaient Cycadales et le conifère Brachyphyllum[28].
D'autres ptérosaures de la Formation de Romualdo comprennent Anhanguera, Araripedactylus, Araripesaurus, Brasileodactylus, cearadactylus, Coloborhynchus, Santanadactylus, Tapejara, tupuxuara,[29] Barbosania, [30] Maaradactylus[31], Tropeognathus[32], et Unwindia[33]. Les thalassodromes ne sont connus que dans cette formation et, bien que des restes postcraniens bien préservés aient été attribués au groupe, ils ne peuvent pas être affectés au genre en raison de leur manque de crânes[34],[35]. La faune de dinosaures comprend théropodes comme Irritator, santanaraptor, mirischia, et pour une période indéterminée unenlagiine dromaeosaur[28]. Les crocodyliformes Araripesuchus et Caririsuchus[36], ainsi que les tortues Brasilemys[37], Cearachelys,[38] Araripemys, Euraxemys[39] et Santanachelys, sont connus à partir des gisements[40]. Il y avait aussi des crevettes de palourde, des oursins, des ostracodes et des mollusques[27]. Les fossiles de poissons bien préservés enregistrent la présence de requins hybodont, guitarfish, orphies, amiids, ophiopsids, oshuniids, pycnodontids, aspidorhynchids, cladocyclids, bananes de mer, chanids, mawsoniids et certaines formes incertaines[41]. Pêgas et ses collègues ont noté que les taxons de ptérosaures de la Formation de Romualdo avaient plusieurs espèces: deux de Thalassodromeus, deux de Tupuxuara et jusqu'à six espèces d’ Anhanguera. Il est possible que toutes les espèces de chaque taxon n'aient pas coexisté dans le temps (comme cela a été proposé pour les ptéranodontidés de la formation de Niobrara), mais il n’existe pas suffisamment de données stratigraphiques pour que la formation de Romualdo puisse le vérifier[2].
Bibliographie
![]() : document utilisé comme source pour la rédaction de cet article.
: document utilisé comme source pour la rédaction de cet article.
- (en) Kellner et Campos, « The function of the cranial crest and jaws of a unique pterosaur from the early Cretaceous of Brazil », Science, vol. 297, no 5580, , p. 389–392 (PMID 12130783, DOI 10.1126/science.1073186).

- (en) Pêgas, Costa et Kellner, « New Information on the osteology and a taxonomic revision of The genus Thalassodromeus (Pterodactyloidea, Tapejaridae, Thalassodrominae) », Journal of Vertebrate Paleontology, vol. 38, no 2, , e1443273 (DOI 10.1080/02724634.2018.1443273).
Liens externes
- Ressources relatives au vivant :
Références
- Kellner et Campos 2002, p. 389–392
- Pêgas, Costa et Kellner 2018, p. e1443273
- Witton 2013.
- (en) Kellner et Campos, « Short note on the ingroup relationships of the Tapejaridae (Pterosauria, Pterodactyloidea) », Boletim do Museu Nacional: Geologia, vol. 75, , p. 1–14
- (en) Kellner, « New information on the Tapejaridae (Pterosauria, Pterodactyloidea) and discussion of the relationships of this clade », Ameghiniana, vol. 41, no 4, , p. 521–534 (ISSN 1851-8044, lire en ligne)
- (en) André Jacques Veldmeijer, « Toothed pterosaurs from the Santana Formation (Cretaceous; Aptian–Albian) of northeastern Brazil », Université d'Utrecht (thèse), Proefschrift Universiteit Utrecht, , p. 11
- (it) G. Leonardi, Sulle Orme Dei Dinosauri, Venise, Erizzo Editrice., , 187–194 p. (ISBN 978-88-7077-018-6), « I rettili volanti »
- (en) Kellner et Campos, « Preliminary description of an unusual pterosaur skull of the Lower Cretaceous from the Araripe Basin », Atas I Simposio Sobre a Bacia do Araripe e Bacias Interiores do Nordeste, , p. 401–405
- (en) Veldmeijer, Signore et Meijer, « Description of two pterosaur (Pterodactyloidea) mandibles from the upper Cretaceous Santana Formation, Brazil », Deinsea, vol. 11, , p. 67–86
- (en) Martill et Naish, « Cranial crest development in the Azhdarchoid pterosaur Tupuxuara, with a review of the genus and tapejarid monophyly », Palaeontology, vol. 49, no 4, , p. 925–941 (DOI 10.1111/j.1475-4983.2006.00575.x)
- (en) Witton, « A new species of Tupuxuara (Thalassodromidae, Azhdarchoidea) from the Lower Cretaceous Santana Formation of Brazil, with a note on the nomenclature of Thalassodromidae », Cretaceous Research, vol. 30, no 5, , p. 1293–1300 (DOI 10.1016/j.cretres.2009.07.006)
- (en) Headden et Campos, « An unusual edentulous pterosaur from the Early Cretaceous Romualdo Formation of Brazil », Historical Biology, vol. 27, no 7, , p. 815–826 (DOI 10.1080/08912963.2014.904302)
- (en) Grellet Tinner et Codrea, « Thalassodromeus sebesensis, an out of place and out of time Gondwanan tapejarid pterosaur », Gondwana Research, vol. 27, no 4, , p. 1673–1679 (DOI 10.1016/j.gr.2014.06.002)
- (en) Dyke, Vremir, Brusatte et Bever, « Thalassodromeus sebesensis — A new name for an old turtle. Comment on "Thalassodromeus sebesensis, an out of place and out of time Gondwanan tapejarid pterosaur", Grellet-Tinner and Codrea (online July 2014 DOI 10.1016/j.gr.2014.06.002) », Gondwana Research, vol. 27, no 4, , p. 1680–1682 (DOI 10.1016/j.gr.2014.08.004, lire en ligne)
- (en) Codrea et Grellet-Tinner, « Reply to comment by Dyke et al. on "Thalassodromeus sebesensis, an out of place and out of time Gondwanan tapejarid pterosaur" by Grellet-Tinner and Codrea (July 2014), DOI 10.1016/j.gr.2014.06.002 », Gondwana Research, vol. 27, no 4, , p. 1683–1685 (DOI 10.1016/j.gr.2014.08.003)
- (en) Unwin, « On the phylogeny and evolutionary history of pterosaurs », Geological Society, London, Special Publications, vol. 217, no 1, , p. 139–190 (DOI 10.1144/GSL.SP.2003.217.01.11)
- (en) Pinheiro, Fortier, Schultz et De Andrade, « New information on Tupandactylus imperator, with comments on the relationships of Tapejaridae (Pterosauria) », Acta Palaeontologica Polonica, vol. 56, no 3, , p. 567–580 (DOI 10.4202/app.2010.0057, lire en ligne)
- (en) Andres, Clark et Xu, « The Earliest Pterodactyloid and the Origin of the Group », Current Biology, vol. 24, no 9, , p. 1011–1016 (PMID 24768054, DOI 10.1016/j.cub.2014.03.030)
- (en) Hone, Naish et Cuthill, « Does mutual sexual selection explain the evolution of head crests in pterosaurs and dinosaurs? », Lethaia, vol. 45, no 2, , p. 139–156 (DOI 10.1111/j.1502-3931.2011.00300.x)
- (en) Bogle, « New find: pterosaur had strange crest, fishing style », National Geographic, (lire en ligne [archive du ])
- (en) S. Chatterjee et R. J. Templin, Posture, locomotion, and paleoecology of pterosaurs, vol. 376, , 64 p. (ISBN 978-0-8137-2376-1, DOI 10.1130/0-8137-2376-0.1, lire en ligne), p. 56
- (en) Humphries, Bonser, Witton et Martill, « Did pterosaurs feed by skimming? Physical modelling and anatomical evaluation of an unusual feeding method », PLOS Biology, vol. 5, no 8, , e204 (PMID 17676976, PMCID 1925135, DOI 10.1371/journal.pbio.0050204)
- (en) Gross, « A new view of pterosaur feeding habits », PLoS Biology, vol. 5, no 8, , e217 (PMID 20076682, PMCID 1925131, DOI 10.1371/journal.pbio.0050217)
- (en) D. Unwin et D. Martill, The Crato Fossil Beds of Brazil : Window into an Ancient World, Cambridge, Cambridge University Press, , 475–524 p. (ISBN 978-0-521-85867-0), « Pterosaurs of the Crato Formation »
- (en) Pêgas et Kellner, « Preliminary mandibular myological reconstruction of Thalassodromeus sethi (Pterodactyloidea: Tapejaridae) », Flugsaurier 2015 Portsmouth, Abstracts, , p. 47–48
- (en) Williams, Kellner et Campos, « Form, function, and the flight of the pterosaur », Science, vol. 297, no 5590, , p. 2207–2208 (JSTOR 3832354)
- (en) Mabesoone et Tinoco, « Palaeoecology of the Aptian Santana Formation (Northeastern Brazil) », Palaeogeography, Palaeoclimatology, Palaeoecology, vol. 14, no 2, , p. 97–118 (ISSN 0031-0182, DOI 10.1016/0031-0182(73)90006-0, lire en ligne)
- (en) Martill, Frey, Sues et R. I. Cruickshank, « Skeletal remains of a small theropod dinosaur with associated soft structures from the Lower Cretaceous Santana Formation of NE Brazil », Canadian Journal of Earth Sciences, vol. 37, no 6, , p. 891–900 (DOI 10.1139/cjes-37-6-891, lire en ligne)
- (en) Barrett, Butler, Edwards et R. Milner, « Pterosaur distribution in time and space: An atlas », Zitteliana Reihe B: Abhandlungen der Bayerischen Staatssammlung für Palaontologie und Geologie, vol. 28, , p. 61–107 (lire en ligne)
- (en) Elgin, A. et Frey, E., « A new ornithocheirid, Barbosania gracilirostris gen. et sp. nov. (Pterosauria, Pterodactyloidea) from the Santana Formation (Cretaceous) of NE Brazil », Swiss Journal of Palaeontology, vol. 130, no 2, , p. 259–275 (DOI 10.1007/s13358-011-0017-4)
- (en) Bantim, R. A. M., Saraiva, A. A. F., Oliveira, G. R. et Sayão, J. M., « A new toothed pterosaur (Pterodactyloidea: Anhangueridae) from the Early Cretaceous Romualdo Formation, NE Brazil », Zootaxa, vol. 3869, no 3, , p. 201–223 (PMID 25283914, DOI 10.11646/zootaxa.3869.3.1)
- (en) Rodrigues et Kellner, « Taxonomic review of the Ornithocheirus complex (Pterosauria) from the Cretaceous of England », ZooKeys, no 308, , p. 1–112 (PMID 23794925, PMCID 3689139, DOI 10.3897/zookeys.308.5559)
- (en) Martill, D. M., « A new pterodactyloid pterosaur from the Santana Formation (Cretaceous) of Brazil », Cretaceous Research, vol. 32, no 2, , p. 236–243 (DOI 10.1016/j.cretres.2010.12.008)
- (en) Aires, Kellner, Müller et Da Silva, « New postcranial elements of the Thalassodrominae (Pterodactyloidea, Tapejaridae) from the Romualdo Formation (Aptian-Albian), Santana Group, Araripe Basin, Brazil », Palaeontology, vol. 57, no 2, , p. 343–355 (DOI 10.1111/pala.12069)
- (en) Buchmann, Rodrigues, Polegario et Kellner, « New information on the postcranial skeleton of the Thalassodrominae (Pterosauria, Pterodactyloidea, Tapejaridae) », Historical Biology, vol. 30, no 8, , p. 1139–1149 (DOI 10.1080/08912963.2017.1343314)
- (en) Figueiredo et Kellner, « A new crocodylomorph specimen from the Araripe Basin (Crato Member, Santana Formation), northeastern Brazil », Paläontologische Zeitschrift, vol. 83, no 2, , p. 323–331 (DOI 10.1007/s12542-009-0016-6)
- (en) de Lapparent de Broin, « The oldest pre-podocnemidid turtle (Chelonii, Pleurodira), from the early Cretaceous, Ceará state, Brasil, and its environment », Treballs del Museu de Geologia de Barcelona, vol. 9, , p. 43–95 (ISSN 2385-4499, lire en ligne)
- (en) Gaffney, D. A. Campos et Hirayama, « Cearachelys, a new side-necked turtle (Pelomedusoides: Bothremydidae) from the Early Cretaceous of Brazil », American Museum Novitates, vol. 3319, , p. 1–20 (DOI 10.1206/0003-0082(2001)319<0001:CANSNT>2.0.CO;2)
- (en) Gaffney, Tong et Meylan, « Evolution of the side-necked turtles: The families Bothremydidae, Euraxemydidae, and Araripemydidae », Bulletin of the American Museum of Natural History, vol. 300, , p. 1–698 (DOI 10.1206/0003-0090(2006)300[1:EOTSTT]2.0.CO;2, lire en ligne)
- (en) Hirayama, « Oldest known sea turtle », Nature, vol. 392, no 6677, , p. 705–708 (ISSN 0028-0836, DOI 10.1038/33669, lire en ligne)
- (en) Brito et Yabumoto, « An updated review of the fish faunas from the Crato and Santana formations in Brazil, a close relationship to the Tethys fauna », Bulletin of Kitakyushu Museum of Natural History and Human History, Series A, vol. 9, (lire en ligne)
 Portail de la paléontologie
Portail de la paléontologie  Portail du Brésil
Portail du Brésil