Peptide antimicrobien
Les peptides antimicrobiens (PAM), (AMPs en anglais) également appelés peptides de défense de l'hôte (HDPs en anglais), font partie de la réponse immunitaire innée trouvée dans toutes les classes de la vie. Des différences fondamentales existent entre les cellules procaryotes et eucaryotes pouvant représenter des cibles pour les peptides antimicrobiens. Ces peptides sont de puissants antibiotiques à large spectre qui présentent un potentiel en tant que nouveaux agents thérapeutiques. Il a été démontré que les peptides antimicrobiens tuent les bactéries à Gram négatif et à Gram positif, les virus enveloppés, les champignons et même les cellules transformées ou cancéreuses[1]. Contrairement à la majorité des antibiotiques classiques, il apparaît que les peptides antimicrobiens déstabilisent fréquemment les membranes biologiques, peuvent former des canaux transmembranaires et peuvent également renforcer l’immunité (en) en jouant le rôle d’ immunomodulateurs.

Structure
Les peptides antimicrobiens constituent un groupe unique et diversifié de molécules, qui sont divisées en sous-groupes sur la base de leur composition et de leur structure en acides aminés[2]. Les peptides antimicrobiens contiennent généralement entre 12 et 50 acides aminés. Ces peptides incluent deux résidus chargés positivement ou plus fournis par l'arginine, la lysine ou, dans les environnements acides, l' histidine et une proportion importante (généralement > 50%) de résidus hydrophobes[3],[4],[5]. Les structures secondaires de ces molécules suivent 4 thèmes, dont i) les hélices α, ii) les brins β dus à la présence de 2 liaisons disulfure ou plus, iii) l’épingle ou la boucle β dues à la présence d’une simple liaison disulfure / ou cyclisation de la chaîne peptidique et iv) étendu[6]. Beaucoup de ces peptides ne sont pas structurés en solution libre et se replient dans leur configuration finale lors de la partition dans des membranes biologiques. Il contient des résidus d'acides aminés hydrophiles alignés le long d'un côté et des résidus d'acides aminés hydrophobes alignés le long du côté opposé d'une molécule hélicoïdale[2]. Cette amphipathicité des peptides antimicrobiens leur permet de se répartir dans la bicouche lipidique de la membrane. La capacité d'association avec des membranes est une caractéristique définitive des peptides antimicrobiens[7],[8] bien que la perméabilisation des membranes ne soit pas nécessaire. Ces peptides exercent diverses activités antimicrobiennes, allant de la perméabilisation membranaire à l’action sur une gamme de cibles cytoplasmiques.
| Type | caractéristiques | AMPs |
|---|---|---|
| Peptides anioniques | riche en acides glutamique et aspartique | Maximin H5 chez les amphibiens, Dermcidine chez l'homme |
| Peptides α-hélicoïdaux cationiques linéaires | manque de cystéine | Cétropines , andropine, moricine , cératotoxine et mélittine d'insectes, Magainin , dermaseptine , bombinine, brevinine-1, esculentines et buforine II d'amphibiens, CAP18 de lapins, LL37 d'humains |
| Peptide cationique enrichi en acide aminé spécifique | riche en proline, arginine, phénylalanine, glycine, tryptophane | abaecin, apidaecins d'abeilles domestiques, prophénine de porcs, indolicidine de bétail. |
| Peptides anioniques et cationiques contenant de la cysteine et formant des liaisons disulfure | contient 1 ~ 3 liaison disulfure | 1 liaison: brevinines, 2 liaisons: protégrine de porc, tachyplesines de limules, 3 liaisons: défenses de l'homme, plus de 3: drosomycine dans les mouches |
Activités
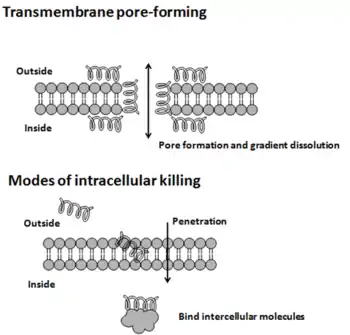
Les modes d'action par lesquels les peptides antimicrobiens tuent les microbes varient [9] et peuvent différer d'une espèce bactérienne à l'autre[10]. Certains peptides antimicrobiens tuent à la fois les bactéries et les champignons, par exemple le psoriasin tue E. coli et plusieurs champignons filamenteux[11]. La membrane cytoplasmique est une cible fréquente, mais les peptides peuvent également interférer avec la synthèse de l'ADN et des protéines, le repliement des protéines et la synthèse de la paroi cellulaire[9]. Le contact initial entre le peptide et l'organisme cible est électrostatique, car la plupart des surfaces bactériennes sont anioniques ou hydrophobes, comme dans le peptide antimicrobien Piscidin. Leur composition en acides aminés, leur caractère amphipathique, leur charge cationique et leur taille leur permettent de s’attacher aux bicouches membranaires et de s’insérer dans celles-ci pour former des pores par des mécanismes à barreaux, à tapis ou à pores toroïdaux. Alternativement, ils peuvent pénétrer dans la cellule pour lier des molécules intracellulaires qui sont essentielles à la vie cellulaire[12]. Les modèles de liaison intracellulaire comprennent l'inhibition de la synthèse de la paroi cellulaire, l'altération de la membrane cytoplasmique, l'activation de l'autolysine, l'inhibition de la synthèse de l'ADN, de l'ARN et des protéines et l'inhibition de certaines enzymes. Cependant, dans de nombreux cas, le mécanisme exact de la mise à mort n'est pas connu. Une technique émergente pour l'étude de tels mécanismes est l' interférométrie à double polarisation[13],[14]. Contrairement à de nombreux antibiotiques classiques, ces peptides semblent être bactéricides[1] au lieu de bactériostatiques. En général, l'activité antimicrobienne de ces peptides est déterminée en mesurant la concentration minimale inhibitrice (CMI), qui est la plus faible concentration de médicament inhibant la croissance bactérienne[15].
Les PAM peuvent posséder de multiples activités, notamment des activités bactériennes anti-gram-positives, anti-fongiques, anti-virales, anti-parasitaires et anti-cancéreuses. Une grande analyse fonctionnelle indique que parmi toutes les activités des PAM, l'amphipathie et la charge, deux propriétés principales des PAM, distinguent le mieux les PAM avec et sans activités bactériennes anti-gram négatif[16]. Cela implique qu'être un PAM ayant des activités bactériennes anti-gram-négatives peut nécessiter une forte amphipathie et une charge positive nette.
Immunomodulation
En plus de tuer directement les bactéries, il a été démontré qu'elles remplissaient un certain nombre de fonctions immunomodulatrices pouvant jouer un rôle dans l'élimination de l'infection, notamment la capacité de modifier l'expression du gène de l'hôte, d'agir comme chimiokines et/ou d'induire la production de chimiokines, en inhibant les protéines - production de cytokines inflammatoires, favorisant la cicatrisation des plaies et modulant les réponses des cellules dendritiques et des cellules de la réponse immunitaire adaptative. Les modèles animaux indiquent que les peptides de défense de l'hôte sont essentiels à la fois pour prévenir et éliminer l'infection. Il semble que de nombreux peptides initialement isolés sous le nom de "peptides antimicrobiens" se soient avérés avoir des fonctions alternatives plus importantes in vivo (par exemple, l'hepcidine[17]).
Mécanismes d'action
Les peptides antimicrobiens forment un groupe vaste et hétérogène de peptides ayant différents mécanismes d'actions, ils ont en commun de permettre la destruction de divers pathogènes (virus, bactéries, champignons). Les défensines s'assemblent en polymères dans la membrane des bactéries pour former un pore et lyser ainsi la cellule. Les histatines inhibent des enzymes indispensables au métabolisme de la cellule.
Les peptides antimicrobiens possédant une charge nette positive sont attirés et incorporés dans les membranes bactériennes chargées négativement. Une fois à l'intérieur de la membrane, ils sont supposés causer des perturbations par trois mécanismes possibles:
- formation de pores toroïdaux
- formation de tapis
- formation de douves de tonneau
Bien que les spécificités de chaque mécanisme diffèrent, toutes proposent une rupture de la membrane induite par un peptide, permettant ainsi une fuite cytoplasmique qui conduit finalement à la mort.
Des travaux récents ont brossé un tableau plus complexe de l'activité des peptides antimicrobiens. Les peptides antimicrobiens peuvent également agir en tant qu'inhibiteurs métaboliques, inhibiteurs de la synthèse de l'ADN, de l'ARN et des protéines et inhibiteurs de la synthèse de la paroi cellulaire ou de la formation de septum. Ils sont également connus pour causer l’agrégation des ribosomes et délocaliser les protéines membranaires.
Ajoutant une couche supplémentaire de complexité, de nombreux peptides antimicrobiens naturels possèdent une activité bactéricide faible. Plutôt que d'inhiber directement la croissance bactérienne, ils sont maintenant connus pour agir de concert avec le système immunitaire de l'hôte par le biais de mécanismes comprenant l'induction de chimiokines, la libération d'histamine et la modulation de l'angiogenèse. Ces effets immunomodulateurs ont récemment commencé à attirer l'attention.
Plusieurs méthodes ont été utilisées pour déterminer les mécanismes de l'activité des peptides antimicrobiens[10],[12]. En particulier, les études de RMN à l'état solide ont fourni une explication de résolution au niveau atomique de la rupture de membrane par des peptides antimicrobiens[18],[19]. Au cours des dernières années, la cristallographie aux rayons X a été utilisée pour décrire en détail la manière dont la famille des défensines végétales rompent les membranes en identifiant les phospholipides clés dans les membranes cellulaires du pathogène[20],[21]. On a pensé que les défensines humaines agissaient par un mécanisme similaire, ciblant les lipides des membranes cellulaires dans le cadre de leur fonction. En fait, il a été démontré que la bêta-défensine 2 humaine tue les champignons pathogènes Candida albicans par le biais d’interactions avec des phospholipides spécifiques[22].
| Les méthodes | Applications |
|---|---|
| Microscopie | visualiser les effets des peptides antimicrobiens sur les cellules microbiennes |
| Spectroscopie d'émission atomique | pour détecter une perte de potassium intracellulaire (une indication que l'intégrité de la membrane bactérienne a été compromise) |
| Colorants fluorescents | mesurer la capacité des peptides antimicrobiens à perméabiliser les vésicules membranaires |
| Formation de canaux ioniques | évaluer la formation et la stabilité d'un pore induit par un peptide antimicrobien |
| Dichroïsme circulaire et dichroïsme circulaire orienté | mesurer l'orientation et la structure secondaire d'un peptide antimicrobien lié à une bicouche lipidique |
| Interférométrie à double polarisation | mesurer les différents mécanismes des peptides antimicrobiens |
| Spectroscopie RMN à l'état solide | mesurer la structure secondaire, l'orientation et la pénétration de peptides antimicrobiens dans des bicouches lipidiques à l'état cristallin liquide biologiquement pertinent |
| Diffraction des neutrons et des rayons x | mesurer les diagrammes de diffraction des pores induits par les peptides dans les membranes de multicouches ou de liquides orientés |
Extraction et caractérisation des peptides antimicrobiens
Extraction par hydrolyse enzymatique de protéines
À partir des années 1990, plusieurs peptides possédant des activités antimicrobiennes vis-à-vis de bactéries à gram-positif et à gram-négatif, ont été mis en évidence et isolés au sein d’hydrolysats de protéines. La nature de ces protéines est très diverse, telle que les caséines, le lysozyme, l’ovotransferrine, la lactoferrine, la β-lactoglobuline.
Les protéines du lait représentent une source potentielle et intéressante de peptides antimicrobiens. Plusieurs protéines du lactosérum (lactoferrine, α-Lactalbumine et β-Lactoglobuline) ainsi que les caséines (caséine αS1, caséine αS2, caséine ҡ) ont permis l’extraction de peptides antimicrobiens lors de leur hydrolyse par des enzymes digestives.
Si la majorité des peptides antimicrobiens préparés par hydrolyse enzymatique de protéine a été isolée à partir de protéines du lait, plusieurs auteurs (Froidevaux et al., 2001, Mak et al., 2004, Daoud et al., 2005 et Nedjar-Arroume et al., 2006) ont rapporté l’obtention de peptides antimicrobiens par hydrolyse enzymatique de l’hémoglobine, protéine connue pour être une source de peptides à activité biologique. Ces peptides sont nommés « hémocidines », « peptides antimicrobiens dérivés de protéines contenant une molécule d’hème ».
Extraction des PAM par hydrolyse pepsique de l'hémoglobine bovine
Hémoglobine bovine :
L’hémoglobine bovine est une protéine de 64 500 Da et de 574 acides aminés**//, constituée d’une partie protéinique, la globine, et d’un hème(*). Quelle que soit l’espèce, l’hème représente la partie constante de l’hémoglobine et les différences se situent au niveau de la globine.*
Structure de l’hémoglobine :
Chez tous les vertébrés, la globine est un tétramère formé par l’association de quatre chaines polypeptidiques identiques deux à deux : deux chaines alpha (ou globines α) et deux chaines bêta (ou globines β). Chacune de ces chaines adopte une conformation spatiale qui lui donne une forme globulaire et ménage une « poche » superficielle dans laquelle se loge l’hème (une molécule de l’hémoglobine peut ainsi fixer quatre molécules d’oxygène).*
Chaine α
La chaine α de 15 053 Da est formée de 141 acides aminés :
V1LSAADKGNV10KAAWGKVGGH20AAEYGAEALE30RMFLSFPTTK40TYFPHFDLSH50GSAQVKGHGA60KVAAALTKAV70EHLDDLPGAL80SELSDLHAHK90LRVDPVNFKL100LSHSLLVTLA110SHLPSDFTPA120VHASLDKFLA130NVSTVLTSKY140R
Chaine β
La chaine β de 15 954 Da possède 146 acides aminés :
M1LTAEEKAAV10TAFWGKVKVD20EVGGEALGRL30LVVYPWTQRF4FESFGDLSTA50DAVMNNPKVK60AHGKKVLDSP70SNGMKGLDDL80KGTFAALSEL90HCDKLHVDPE100NFKLLGNVLV110VVLARNFGKF120FTPVLQADFQ130KVVAGVANAL140AHRYH
Structure de l’hème :
L’hème est constitué d’une protoporphyrine ayant en son centre un atome de fer (Fe2+). La protoporphyrine est formée de quatre cycles pyrroliques unis par l’intermédiaire de ponts méthylés (-CH=) et substitués par des groupes méthyle, propionate et vinyle.
Pepsine
La pepsine est une endo-protéase digestive, une enzyme du règne animal découverte par le docteur Beaumont en 1833.
C’est une enzyme qui subdivise les protéines en petits peptides (protéase). Elle est sécrétée dans l’estomac et c’est l’une des principales enzymes de digestion chez l’homme et plusieurs types d’animaux.
Structure de la pepsine :
La pepsine avec une masse moléculaire de 34 600 Da est constituée de 326 acides aminés répartis en deux domaines principaux : Nt (AA1-AA176) et le domaine Ct (AA177-AA326), formant une chaîne peptidique. La pepsine possède également trois ponts disulfure répartis selon Cys45-Cys50, Cys206-Cys210, Cys249-Cys282.
Mécanisme d’action de l’hydrolyse pepsique :
La pepsine appartient à la famille des protéases, enzymes spécialisées dans la coupure de la liaison peptidique. Son mécanisme d’action est basé sur la reconnaissance d’un motif structural du substrat à hydrolyser au niveau du site actif de l’enzyme. Schechter et Berger (1967, 1968) ont établi une nomenclature concernant la spécificité des protéases. L’enzyme va cliver la liaison peptidique située entre les résidus (acides aminés) P1 et P1’ localisés au niveau des sites d’interaction de l’enzyme nommés S1 et S1’.
La pepsine est une protéase fonctionnant avec l’acide aspartique (Asp) dans son site actif. Celui-ci comprend deux résidus d’acide aspartique proches géométriquement Asp32 et Asp215, ainsi que d’autres acides aminés: Ile30, Ser39, Ile120, Ala124, Ile213, Thr218, Ala301, Thr305, forment une crevasse où vient se loger le substrat. Lors de la coupure de la liaison peptidique, les deux acides aspartiques jouent simultanément le rôle de donneur et de receveur de protons.*
La pepsine coupe préférentiellement en Cterminal des acides aminés aromatiques (Phé et Tyr) et hydrophobes (Leu, Ala, Glu, et Gln), en position P1. Certains acides aminés peuvent avoir un effet négatif sur le clivage de la liaison peptidique par la pepsine selon leur position au sein de son site actif : l’arginine en position P1 et P3, la lysine et l’histidine en position P3. La proline a un effet positif lorsqu’elle se situe en position P3 ou P4, et un effet négatif quand elle est en position P2 ou P3’. La pepsine ne coupe pas en présence de la proline et la glycine en position P1.*
Usage thérapeutique
Les peptides antimicrobiens ont été utilisés comme agents thérapeutiques; leur utilisation est généralement limitée à une administration par voie intraveineuse ou à des applications topiques en raison de leur courte demi-vie. En , les peptides antimicrobiens suivants étaient utilisés en clinique[23]:
- Bacitracine pour la pneumonie, topique
- Bocéprévir , Hépatite C (oral, peptide cyclique)
- Dalbavancine , infections bactériennes, IV
- Daptomycine , infections bactériennes, IV
- Enfuvirtide , VIH, injection sous-cutanée
- Oritavancine , infections bactériennes, IV
- Teicoplanine , infections bactériennes, IV
- Télaprévir , Hépatite C, peptide cyclique oral
- Télavancine , infection bactérienne, IV
- Vancomycine , infection bactérienne, IV.
- Guavanine 2 , infection bactérienne à Gram positif et à Gram négatif également.
Recherche
Certaines des cécropines (par exemple la cécropine A et la cécropine B) ont des propriétés anticancéreuses et sont appelées peptides anticancéreux (ACPs en anglais)[24]. Les ACP hybrides basés sur Cecropin A ont été étudiés pour leurs propriétés anticancéreuses. Des recherches ont récemment été menées pour identifier des peptides antimicrobiens potentiels provenant de procaryotes [25] et d'organismes aquatiques tels que les poissons [26],[27] et les mollusques et crustacés [28].
Sélectivité
Dans la compétition des cellules bactériennes et des cellules hôtes avec les peptides antimicrobiens, les peptides antimicrobiens vont interagir préférentiellement avec la cellule bactérienne avec les cellules de mammifère, ce qui leur permet de tuer des micro-organismes sans être significativement toxiques pour les cellules de mammifère[29]. La sélectivité est une caractéristique très importante des peptides antimicrobiens et peut garantir leur fonction d'antibiotique dans les systèmes de défense de l'hôte.
Les facteurs
Certains facteurs sont étroitement liés à la propriété de sélectivité des peptides antimicrobiens, parmi lesquels la propriété cationique contribue le plus. Étant donné que la surface des membranes bactériennes est plus chargée négativement que les cellules de mammifère, les peptides antimicrobiens présenteront des affinités différentes pour les membranes bactériennes et les membranes cellulaires des mammifères[30].
En outre, il existe également d'autres facteurs qui affecteront la sélectivité. Il est bien connu que le cholestérol est normalement largement distribué dans les membranes cellulaires des mammifères en tant qu’agents stabilisants, mais absent dans les membranes des cellules bactériennes; et la présence de ces cholestérols réduira aussi généralement l'activité des peptides antimicrobiens, du fait soit de la stabilisation de la bicouche lipidique, soit des interactions entre le cholestérol et le peptide. Ainsi, le cholestérol contenu dans les cellules de mammifères protégera les cellules des attaques des peptides antimicrobiens[31].
En outre, il est bien connu que le potentiel trans-membranaire (ou potentiel électrochimique de membrane) affecte les interactions peptide-lipides[32]. Un potentiel trans-membranaire interne négatif existe de la feuille externe à la feuille interne des membranes cellulaires et ce potentiel trans-membranaire interne négatif facilitera la perméabilisation de la membrane, probablement en facilitant l'insertion de peptides chargés positivement dans les membranes. En comparaison, le potentiel trans-membranaire des cellules bactériennes est plus négatif que celui des cellules de mammifère normales; la membrane bactérienne risque donc d'être attaquée par les peptides antimicrobiens chargés positivement.
De même, on pense également que l'augmentation de la force ionique [31] qui réduit généralement l'activité de la plupart des peptides antimicrobiens, contribue en partie à la sélectivité des peptides antimicrobiens en affaiblissant les interactions électrostatiques requises pour l'interaction initiale.
Mécanisme
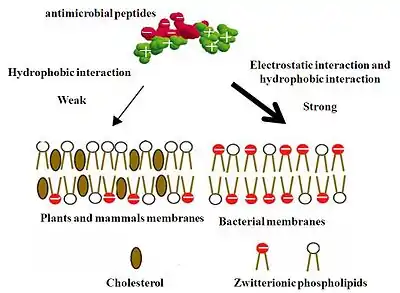
Les membranes cellulaires des bactéries sont riches en phospholipides acides, tels que le phosphatidylglycérol et la cardiolipine[29],[33]. Ces groupes de tête phospholipidiques sont fortement chargés négativement. Par conséquent, les feuillets les plus à l'extérieur de la bicouche qui sont exposés à l'extérieur des membranes bactériennes sont plus attrayants pour l'attaque des peptides antimicrobiens chargés positivement. Ainsi, l'interaction entre les charges positives des peptides antimicrobiens et les membranes bactériennes chargées négativement concerne principalement les interactions électrostatiques, qui constituent le principal moteur de l'association cellulaire. De plus, comme les peptides antimicrobiens forment des structures avec une face chargée positivement et une face hydrophobe, il existe également des interactions hydrophobes entre les régions hydrophobes des peptides antimicrobiens et la surface des phospholipides zwitterioniques (électriquement neutres) seulement comme un effet mineur dans ce cas.
En revanche, la partie externe des membranes des plantes et des mammifères est principalement composée de lipides sans charge nette, car la plupart des lipides avec des groupes de tête chargés négativement sont principalement séquestrés dans la foliole interne des membranes plasmiques[30]. Ainsi, dans le cas des cellules de mammifères, les surfaces extérieures des membranes sont généralement constituées de phosphatidylcholine et de sphingomyéline zwitterioniques, même si une petite partie des surfaces extérieures de la membrane contient des gangliosides chargés négativement. Par conséquent, l'interaction hydrophobe entre la face hydrophobe des peptides antimicrobiens amphipathiques et les phospholipides zwitterioniques à la surface des membranes cellulaires des mammifères joue un rôle majeur dans la formation de la liaison peptide-cellule[34]. Cependant, l'interaction hydrophobe est relativement faible comparée à l'interaction électrostatique, ainsi, les peptides antimicrobiens vont interagir préférentiellement avec les membranes bactériennes. [réf. nécessaire] L'interférométrie à double polarisation a été utilisée in vitro pour étudier et quantifier l'association avec le groupe de tête, l'insertion dans la bicouche, la formation de pores et la rupture éventuelle de la membrane[35],[36].
Contrôle
De nombreux efforts ont été déployés pour contrôler la sélectivité des cellules. Par exemple, des tentatives ont été faites pour modifier et optimiser les paramètres physico-chimiques des peptides afin de contrôler les sélectivités, y compris la charge nette, l'hélicité, l'hydrophobie par résidu (H), le moment hydrophobe (μ) et l'angle sous-tendu par la face de l'hélice polaire chargée positivement (Φ)[32]. On pense que d'autres mécanismes, tels que l'introduction de D-aminoacides et d'aminoacides fluorés dans la phase hydrophobe, brisent la structure secondaire et réduisent ainsi les interactions hydrophobes avec les cellules de mammifère. Il a également été découvert que la substitution Pro → Nlys dans les peptides antimicrobiens β-turn contenant du Pro était une stratégie prometteuse pour la conception de nouveaux peptides antimicrobiens sélectifs pour les cellules bactériennes dotés de mécanismes d'action intracellulaires[37]. Il a été suggéré que la fixation directe de la magainine à la surface du substrat diminuait la liaison des cellules non spécifiques et conduisait à une limite de détection améliorée pour les cellules bactériennes telles que Salmonella et E. coli[38].
Résistance bactérienne
Les bactéries utilisent diverses stratégies de résistance pour éviter la destruction des peptides antimicrobiens[12]. Certains micro-organismes modifient les charges nettes en surface. Staphylococcus aureus transporte la D-alanine du cytoplasme à l'acide teichoïque de surface, ce qui réduit la charge négative nette en introduisant des groupes amino basiques[39]. S. aureus modifie également ses membranes anioniques via MprF avec L-lysine, augmentant la charge nette positive[39]. L'interaction des peptides antimicrobiens avec les cibles membranaires peut être limitée par le polysaccharide en capsule de Klebsiella pneumoniae[40]. Des altérations se produisent dans le lipide A. Les espèces de Salmonella réduisent la fluidité de leur membrane externe en augmentant les interactions hydrophobes entre un nombre accru de queues acyles du lipide A en ajoutant du myristate au lipide A avec du 2-hydroxymyristate et en formant du lipide hepta-acylé en ajoutant du palmitate. On pense que le moment hydrophobe accru retarde ou supprime l’insertion de peptides antimicrobiens et la formation de pores. Les résidus subissent une altération dans les protéines membranaires. Chez certaines bactéries à Gram négatif, l’altération de la production de protéines de la membrane externe est corrélée à la résistance à la destruction par des peptides antimicrobiens[41]. L'Hemophilus influenzae non typable transporte les PAM à l'intérieur de la cellule, où ils sont dégradés. En outre, H. influenzae remodèle ses membranes pour lui donner l’impression que la bactérie a déjà été attaquée avec succès par les Φ, ce qui l’empêche d’être attaquée par davantage de PAM[42]. Les transporteurs de cassette de liaison à l'ATP importent des peptides antimicrobiens et la pompe d'efflux à division de cellules à résistance-nodulation exporte des peptides antimicrobiens[43]. Les deux transporteurs ont été associés à une résistance aux peptides antimicrobiens. Les bactéries produisent des enzymes protéolytiques susceptibles de dégrader les peptides antimicrobiens et donc de les rendre résistantes[44]. Les vésicules de la membrane externe produites par les bactéries à Gram négatif lient les peptides antimicrobiens et les séquestrent loin des cellules, protégeant ainsi les cellules[45]. Les vésicules de la membrane externe sont également connues pour contenir diverses protéases, peptidases et autres enzymes lytiques, qui peuvent jouer un rôle dans la dégradation des molécules de peptide extracellulaire et d’acide nucléique, qui, si on les laisse atteindre les cellules bactériennes, peuvent être dangereuses pour les cellules. La signalisation cyclique-di-GMP avait également été impliquée dans la régulation de la résistance aux peptides antimicrobiens chez Pseudomonas aeruginosa[46].
Bien que ces exemples montrent que la résistance peut évoluer naturellement, il est de plus en plus préoccupant que l’utilisation de copies pharmaceutiques de peptides antimicrobiens puisse rendre la résistance plus fréquente et plus rapide. Dans certains cas, la résistance à ces peptides utilisés en tant que produit pharmaceutique pour traiter des problèmes médicaux peut entraîner une résistance, non seulement à l'application médicale des peptides, mais à la fonction physiologique de ces peptides[47],[48].
Exemples
Les peptides antimicrobiens sont produits par toutes les espèces connues, y compris les peptides de bactéries, de champignons, d’hydres, d’insectes ( mastoparan, ponératoxine, cécropine, moricine, mélittine et autres)[49], de grenouilles (magainine, dermaseptine et autres)[50], aviaire (par exemple, la cathélicidine et les défensines)[51], de mammifères (par exemple, la cathélicidine, les défensines et les protégrines ). Les recherches se sont multipliées ces dernières années pour mettre au point des mimétiques artificielles de peptides antimicrobiens tels que les SNAPP , en raison notamment du coût prohibitif de la production de AMP d'origine naturelle[52]. Un exemple de ceci est le peptide C18G facialement cationique , qui a été conçu à partir du domaine C-terminal du facteur IV plaquettaire humain[53]. Actuellement, le peptide antimicrobien le plus largement utilisé est la nisine. Étant le seul peptide antimicrobien approuvé par la FDA, il est couramment utilisé comme conservateur artificiel[54].
Bioinformatique
Plusieurs bases de données bio-informatiques existent pour cataloguer les peptides antimicrobiens, telles que ADAM (base de données de peptides antimicrobiens) [55], APD (base de données de peptides antimicrobiens), BioPD (base de données de peptides biologiquement actifs), CAMP (collecte de séquences et de structures de peptides antimicrobiens)[56], DBAASP (base de données sur l'activité antimicrobienne et la structure des peptides) et LAMP (PAM de liaison). Les bases de données de peptides antimicrobiens peuvent être divisées en deux catégories sur la base de la source de peptides qu'elles contiennent, en tant que bases de données spécifiques et bases de données générales. Ces bases de données contiennent divers outils d’analyse et de prévision des peptides antimicrobiens. Par exemple, CAMP contient des prévisions AMP, un calculateur de caractéristiques, une recherche BLAST, ClustalW, VAST, PRATT, roue hélicoïdale, etc. En outre, ADAM permet aux utilisateurs de rechercher ou de parcourir des relations séquence-structure AMP. Les peptides antimicrobiens englobent souvent un large éventail de catégories telles que les peptides antifongiques, antibactériens et antituberculeux. Des outils tels que PeptideRanker[57] et PeptideLocator[58] permettent de prédire les peptides antimicrobiens tandis que d'autres ont été développés pour prédire les activités antifongiques et antituberculeuses[59],[60]. AntiMPmod a été développé pour prédire le potentiel antimicrobien de peptides modifiés chimiquement[61] (voir liens externes).
Notes et références
- « Antimicrobial peptides: premises and promises », International Journal of Antimicrobial Agents, vol. 24, no 6, , p. 536–47 (PMID 15555874, DOI 10.1016/j.ijantimicag.2004.09.005)
- « Mechanisms of antimicrobial peptide action and resistance », Pharmacological Reviews, vol. 55, no 1, , p. 27–55 (PMID 12615953, DOI 10.1124/pr.55.1.2)
- « Ribosomally synthesized peptides with antimicrobial properties: biosynthesis, structure, function, and applications », Biotechnology Advances, vol. 21, no 6, , p. 465–99 (PMID 14499150, DOI 10.1016/S0734-9750(03)00077-6)
- « Host-defense antimicrobial peptides: importance of structure for activity », Current Pharmaceutical Design, vol. 8, no 9, , p. 727–42 (PMID 11945168, DOI 10.2174/1381612023395358)
- « LL-37, the only human member of the cathelicidin family of antimicrobial peptides », Biochimica et Biophysica Acta, vol. 1758, no 9, , p. 1408–25 (PMID 16716248, DOI 10.1016/j.bbamem.2006.03.030)
- « The human beta-defensin-3, an antibacterial peptide with multiple biological functions », Biochimica et Biophysica Acta, vol. 1758, no 9, , p. 1499–512 (PMID 16978580, DOI 10.1016/j.bbamem.2006.07.007)
- « Role of membranes in the activities of antimicrobial cationic peptides », FEMS Microbiology Letters, vol. 206, no 2, , p. 143–9 (PMID 11814654, DOI 10.1111/j.1574-6968.2002.tb11000.x)
- « Antibacterial activity of linear peptides spanning the carboxy-terminal beta-sheet domain of arthropod defensins », Peptides, vol. 27, no 11, , p. 2614–23 (PMID 16914230, DOI 10.1016/j.peptides.2006.06.010)
- « The expanding scope of antimicrobial peptide structures and their modes of action », Trends in Biotechnology, vol. 29, no 9, , p. 464–72 (PMID 21680034, DOI 10.1016/j.tibtech.2011.05.001)
- « Production and evaluation of an antimicrobial peptide-containing wafer formulation for topical application », Current Microbiology, vol. 66, no 3, , p. 271–8 (PMID 23183933, DOI 10.1007/s00284-012-0268-3)
- « Disulphide-reduced psoriasin is a human apoptosis-inducing broad-spectrum fungicide », Proceedings of the National Academy of Sciences of the United States of America, vol. 112, no 42, , p. 13039–44 (PMID 26438863, PMCID 4620902, DOI 10.1073/pnas.1511197112, Bibcode 2015PNAS..11213039H)
- « Antimicrobial peptides: pore formers or metabolic inhibitors in bacteria? », Nature Reviews. Microbiology, vol. 3, no 3, , p. 238–50 (PMID 15703760, DOI 10.1038/nrmicro1098)
- « Effect of acyl chain structure and bilayer phase state on binding and penetration of a supported lipid bilayer by HPA3 », European Biophysics Journal : EBJ, vol. 40, no 4, , p. 503–14 (PMID 21222117, DOI 10.1007/s00249-010-0664-1)
- « Real-time quantitative analysis of lipid disordering by aurein 1.2 during membrane adsorption, destabilisation and lysis », Biochimica et Biophysica Acta, vol. 1798, no 10, , p. 1977–86 (PMID 20599687, DOI 10.1016/j.bbamem.2010.06.023)
- John D. Turnidge et Jan M. Bell, Antibiotics in Laboratory Medicine, Baltimore, Md., , 4e éd., 52–111 p. (ISBN 978-0-683-05169-8), « Susceptibility testing of Antimicrobials in liquid media. National Committee of Laboratory Safety and Standards (NCLSS) »
- « Large-Scale Analysis of Antimicrobial Activities in Relation to Amphipathicity and Charge Reveals Novel Characterization of Antimicrobial Peptides », Molecules (Basel, Switzerland), vol. 22, no 11, (PMID 29165350, DOI 10.3390/molecules22112037)
- « The solution structure of human hepcidin, a peptide hormone with antimicrobial activity that is involved in iron uptake and hereditary hemochromatosis », The Journal of Biological Chemistry, vol. 277, no 40, , p. 37597–603 (PMID 12138110, DOI 10.1074/jbc.M205305200)
- « MSI-78, an analogue of the magainin antimicrobial peptides, disrupts lipid bilayer structure via positive curvature strain », Biophysical Journal, vol. 84, no 5, , p. 3052–60 (PMID 12719236, PMCID 1302867, DOI 10.1016/S0006-3495(03)70031-9, Bibcode 2003BpJ....84.3052H)
- « Mechanism of lipid bilayer disruption by the human antimicrobial peptide, LL-37 », Biochemistry, vol. 42, no 21, , p. 6545–58 (PMID 12767238, DOI 10.1021/bi0273563)
- (en) « X-ray structure of a carpet-like antimicrobial defensin-phospholipid membrane disruption complex », Nature Communications, vol. 9, no 1, , p. 1962 (PMID 29773800, DOI 10.1038/s41467-018-04434-y)
- « Phosphoinositide-mediated oligomerization of a defensin induces cell lysis », eLife, vol. 3, , e01808 (PMID 24692446, PMCID 3968744, DOI 10.7554/elife.01808)
- (en) Järvå, Phan, Lay et Caria, « Human β-defensin 2 kills Candida albicans through phosphatidylinositol 4,5-bisphosphate–mediated membrane permeabilization », Science Advances, vol. 4, no 7, , eaat0979 (ISSN 2375-2548, DOI 10.1126/sciadv.aat0979, lire en ligne)
- « Designing improved active peptides for therapeutic approaches against infectious diseases », Biotechnology Advances, vol. 36, no 2, , p. 415–429 (PMID 29330093, DOI 10.1016/j.biotechadv.2018.01.004)
- « Studies on anticancer activities of antimicrobial peptides », Biochimica et Biophysica Acta, vol. 1778, no 2, , p. 357–75 (PMID 18078805, PMCID 2238813, DOI 10.1016/j.bbamem.2007.11.008)
- Hassan M, Kjos M, IF, Diep DB, Lotfipour F. Peptides antimicrobiens naturels issus de bactéries: caractéristiques et applications potentielles pour lutter contre la résistance aux antibiotiques. J Appl Microbiol. 2012; 113 (4): 723-36. https://doi.org/10.1111/j.1365-2672.2012.05338.x
- Kumaresan V, P Bhatt, MR Ganesh, R Harikrishnan, M Arasu, NA Al-Dhabi, M Pasupuleti, K Marimuthu, J. Arockiaraj. Mol Immunol. 2015; 68 (2 Pt B): 421-33. https://doi.org/10.1016/j.molimm.2015.10.001
- Arasu A, Kumaresan V, MR Ganesh, Pasupuleti M, Arasu MV, Al-Dhabi NA, Arockiaraj J. Activité bactéricide du peptide de liaison à la membrane dérivé de la galectine de poisson 4 étiqueté avec de l'oligotryptophane. Dev Comp Immunol. 2017; 71: 37-48. https://doi.org/10.1016/j.dci.2017.01.019
- Arockiaraj J, MK Chaurasia, Kumaresan V, R Palanisamy, R Harikrishnan, M Pasupuleti, Kasi M. Macrobrachium rosenbergii liant la lectine: synthèse des peptides MrMBL-N20 et MrMBL-C16 et de leurs gènes transcriptionnels et génétiques. Poissons Crustacés Immunol. 2015; 43 (2): 364-74. https://doi.org/10.1016/j.fsi.2014.12.036
- « Control of cell selectivity of antimicrobial peptides », Biochimica et Biophysica Acta, vol. 1788, no 8, , p. 1687–92 (PMID 18952049, DOI 10.1016/j.bbamem.2008.09.013)
- « Antimicrobial and host-defense peptides as new anti-infective therapeutic strategies », Nature Biotechnology, vol. 24, no 12, , p. 1551–7 (PMID 17160061, DOI 10.1038/nbt1267)
- « Antimicrobial peptides of multicellular organisms », Nature, vol. 415, no 6870, , p. 389–95 (PMID 11807545, DOI 10.1038/415389a, Bibcode 2002Natur.415..389Z)
- « Molecular basis for membrane selectivity of an antimicrobial peptide, magainin 2 », Biochemistry, vol. 34, no 10, , p. 3423–9 (PMID 7533538, DOI 10.1021/bi00010a034)
- « Design and synthesis of cationic antimicrobial peptides with improved activity and selectivity against Vibrio spp », International Journal of Antimicrobial Agents, vol. 32, no 2, , p. 130–8 (PMID 18586467, DOI 10.1016/j.ijantimicag.2008.04.003)
- « Molecular evolution of animal antimicrobial peptides: widespread moderate positive selection », Journal of Evolutionary Biology, vol. 18, no 6, , p. 1387–94 (PMID 16313451, DOI 10.1111/j.1420-9101.2005.00925.x)
- « Interaction of an artificial antimicrobial peptide with lipid membranes », Biochimica et Biophysica Acta, vol. 1788, no 2, , p. 333–44 (PMID 19013127, DOI 10.1016/j.bbamem.2008.10.005)
- « Molecular imaging and orientational changes of antimicrobial peptides in membranes », Advances in Experimental Medicine and Biology, vol. 611, , p. 313–5 (PMID 19400207, DOI 10.1007/978-0-387-73657-0_140)
- « Effects of Pro --> peptoid residue substitution on cell selectivity and mechanism of antibacterial action of tritrpticin-amide antimicrobial peptide », Biochemistry, vol. 45, no 43, , p. 13007–17 (PMID 17059217, DOI 10.1021/bi060487)
- « Antimicrobial peptides for detection of bacteria in biosensor assays », Analytical Chemistry, vol. 77, no 19, , p. 6504–8 (PMID 16194120, DOI 10.1021/ac050639r)
- « Inactivation of the dlt operon in Staphylococcus aureus confers sensitivity to defensins, protegrins, and other antimicrobial peptides », The Journal of Biological Chemistry, vol. 274, no 13, , p. 8405–10 (PMID 10085071, DOI 10.1074/jbc.274.13.8405)
- « Capsule polysaccharide mediates bacterial resistance to antimicrobial peptides », Infection and Immunity, vol. 72, no 12, , p. 7107–14 (PMID 15557634, PMCID 529140, DOI 10.1128/IAI.72.12.7107-7114.2004)
- « Role of YadA in resistance of Yersinia enterocolitica to phagocytosis by human polymorphonuclear leukocytes », Infection and Immunity, vol. 62, no 4, , p. 1275–81 (PMID 8132334, PMCID 186269)
- « Sap transporter mediated import and subsequent degradation of antimicrobial peptides in Haemophilus », PLoS Pathogens, vol. 7, no 11, , e1002360 (PMID 22072973, PMCID 3207918, DOI 10.1371/journal.ppat.1002360)
- « Multidrug efflux pumps of gram-negative bacteria », Journal of Bacteriology, vol. 178, no 20, , p. 5853–9 (PMID 8830678, PMCID 178438)
- « The degradation of human endothelial cell-derived perlecan and release of bound basic fibroblast growth factor by stromelysin, collagenase, plasmin, and heparanases », The Journal of Biological Chemistry, vol. 271, no 17, , p. 10079–86 (PMID 8626565, DOI 10.1074/jbc.271.17.10079)
- « Molecular characterization and functional analysis of outer membrane vesicles from the antarctic bacterium Pseudomonas syringae suggest a possible response to environmental conditions », Journal of Proteome Research, vol. 13, no 3, , p. 1345–58 (PMID 24437924, DOI 10.1021/pr4009223)
- « Bis-(3'-5')-cyclic dimeric GMP regulates antimicrobial peptide resistance in Pseudomonas aeruginosa », Antimicrobial Agents and Chemotherapy, vol. 57, no 5, , p. 2066–75 (PMID 23403434, PMCID 3632963, DOI 10.1128/AAC.02499-12)
- « Therapeutic antimicrobial peptides may compromise natural immunity », Biology Letters, vol. 8, no 3, , p. 416–8 (PMID 22279153, PMCID 3367763, DOI 10.1098/rsbl.2011.1203)
- « Antimicrobial peptides », Pharmaceuticals, vol. 6, no 12, , p. 1543–75 (PMID 24287494, DOI 10.3390/ph6121543)
- « Insect antimicrobial peptides OPM », Orientations of Proteins in Membranes database (OPM)
- « Amphibian antimicrobial peptides », Orientations of Proteins in Membranes database (OPM)
- Yang M, Zhang C, et al., « Structure-function analysis of Avian β-defensin-6 and β-defensin-12: role of charge and disulfide bridges », BMC Microbiology, vol. 16, (DOI 10.1186/s12866-016-0828-y)
- Niedermaier, « Synthetic Mimics of Antimicrobial Peptides: A New Wave of Antibiotics » [archive du ], Department of Chemistry, University of Georgia, (consulté le )
- « Role of Cationic Side Chains in the Antimicrobial Activity of C18G », Molecules, vol. 23, no 2, , p. 329 (PMID 29401708, DOI 10.3390/molecules23020329)
- « Biomedical applications of nisin », Journal of Applied Microbiology, vol. 120, no 6, , p. 1449–65 (PMID 26678028, PMCID 4866897, DOI 10.1111/jam.13033)
- « A large-scale structural classification of antimicrobial peptides », BioMed Research International, vol. 2015, , p. 475062 (PMID 26000295, PMCID 4426897, DOI 10.1155/2015/475062)
- « CAMP: Collection of sequences and structures of antimicrobial peptides », Nucleic Acids Research, vol. 42, no Database issue, , D1154-8 (PMID 24265220, PMCID 3964954, DOI 10.1093/nar/gkt1157)
- « Towards the improved discovery and design of functional peptides: common features of diverse classes permit generalized prediction of bioactivity », PLOS One, vol. 7, no 10, , e45012 (PMID 23056189, DOI 10.1371/journal.pone.0045012, Bibcode 2012PLoSO...745012M)
- « PeptideLocator: prediction of bioactive peptides in protein sequences », Bioinformatics, vol. 29, no 9, , p. 1120–6 (PMID 23505299, DOI 10.1093/bioinformatics/btt103)
- « In Silico Approach for Prediction of Antifungal and Peptides », Frontiers in Microbiology, vol. 9, , p. 323 (PMID 29535692, DOI 10.3389/fmicb.2018.00323)
- « Prediction of Antitubercular Peptides From Sequence Information Using Ensemble Classifier and Hybrid Features », Frontiers in Pharmacology, vol. 9, , p. 954 (DOI 10.3389/fphar.2018.00954)
- « Prediction of antimicrobial potential of a chemically modified peptide from its tertiary structure », Frontiers in Microbiology, vol. 9, , p. 2551 (DOI 10.3389/fmicb.2018.02551)
Voir aussi
- Cathélicidine
- Aurein
- Copsin
- Protéines membranaires périphériques
- Nombre de colonies virtuelles
Articles connexes
- Protéine membranaire
- Cathélicidine
- Défensine
Liens externes
- (en) Sara Wilcox, Cationic peptides: a new hope, The Science Creative Quaterly, Université de la Colombie-Britannique.
- Anne-Sophie Michel, À la découverte des peptides antimicrobiens, université Henri Poincaré - Nancy-I (thèse).
Lectures complémentaires
- Giuliani A, Pirri G, Nicoletto S, « Antimicrobial peptides: an overview of a promising class of therapeutics », Cent. Eur. J. Biol., vol. 2, no 1, , p. 1–33 (DOI 10.2478/s11535-007-0010-5)
- Langham AA, Ahmad AS, Kaznessis YN, « On the nature of antimicrobial activity: a model for protegrin-1 pores », Journal of the American Chemical Society, vol. 130, no 13, , p. 4338–46 (PMID 18335931, PMCID 2474802, DOI 10.1021/ja0780380)
- Mátyus E, Kandt C, Tieleman DP, « Computer simulation of antimicrobial peptides », Current Medicinal Chemistry, vol. 14, no 26, , p. 2789–98 (PMID 18045125, DOI 10.2174/092986707782360105)
- Moore AJ, Devine DA, Bibby MC, « Preliminary experimental anticancer activity of cecropins », Peptide Research, vol. 7, no 5, , p. 265–9 (PMID 7849420)
- Thomas S, Karnik S, Barai RS, Jayaraman VK, Idicula-Thomas S, « CAMP: a useful resource for research on antimicrobial peptides », Nucleic Acids Research, vol. 38, no Database issue, , D774-80 (PMID 19923233, PMCID 2808926, DOI 10.1093/nar/gkp1021)
Liens externes
- ADAM (Une base de données de peptides antimicrobiens) sur ntou.edu.tw
- AntiFP Prédiction de peptides antifongiques
- AntiMPmod Prédiction du potentiel antimicrobien des peptides modifiés
- (en) MeSH Antimicrobial+Cationic+Peptides
- AntiTbPred Prédiction de peptides antituberculeux
- Base de données sur les peptides antimicrobiens du centre médical de l'Université du Nebraska
- Serveur de prévision AMP basé sur Deep Learning Scanner Antimicrobial Scptide Scanner
- AntiTbPdb Base de données peptidique antituberculeuse
- BioPD au Centre des sciences de la santé de l'Université de Pékin
- CAMP: Collection de peptides antimicrobiens à l'Institut national de recherche sur la santé de la reproduction (NIRRH)
- DBAASP - Base de données sur l'activité antimicrobienne et la structure des peptides]
- LAMP à l'Université de Fudan
- PeptideLocator Prédiction de peptides fonctionnels, y compris de peptides antimicrobiens, dans une séquence protéique
- PeptideRanker Peptide bioactif, y compris un peptide antimicrobien, prédiction
 Portail de la biologie
Portail de la biologie