Phalloïdine
La phalloïdine fait partie des toxines extraites des champignons comme l'amanite phalloïde, Amanita phalloides, mais aussi d'Amanita verna, d'Amanita virosa et Galerina marginata connues sous le nom de phallotoxines. Elle se lie à l'actine, empêchant sa dépolymérisation et empoisonnant la cellule. Cette propriété de liaison avec l'actine lui permet d'être utilisée en imagerie cellulaire afin de les visualiser.
| Phalloïdine | |
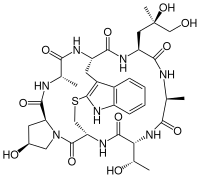
| |
| Structure de la phalloïdine | |
| Identification | |
|---|---|
| No CAS | |
| No ECHA | 100.037.697 |
| Propriétés chimiques | |
| Formule | C35H48N8O11S [Isomères] |
| Masse molaire[1] | 788,868 ± 0,041 g/mol C 53,29 %, H 6,13 %, N 14,2 %, O 22,31 %, S 4,06 %, |
| Unités du SI et CNTP, sauf indication contraire. | |
Découverte
La phalloïdine est l'une des toxines actives contenues dans plusieurs des champignons les plus toxiques au monde : la célèbre amanite phalloïde (Amanita phalloides) à laquelle elle doit son nom, mais aussi l'Amanite printanière, l'Amanite vireuse et Galerina marginata. Ces champignons contiennent cependant d'autres phallotoxines et amatoxines. Bien que les phallotoxines soient extrêmement toxiques pour les cellules du foie et du rein, où elles perturbent la dynamique du cytosquelette d'actine en empêchant la dépolymérisation des filaments[2], elles n'ont qu’un impact léger sur la toxicité générale de l’amanite phalloïde, étant très peu absorbées au niveau intestinal[3]. Par exemple, la phalloïdine est retrouvée dans une autre espèce, l’amanite rougissante (Amanita rubescens) parfaitement comestible si elle est bien cuite[4].
On doit les premiers travaux sur la phalloïdine au prix Nobel Heinrich Wieland dans les années 1930. Cette toxine fut finalement purifiée et cristallisée en 1937 par le fils et étudiant d'Heinrich, Feodor Lynen (qui obtiendra le prix Nobel en 1964 pour ses travaux sur le métabolisme du cholestérol), et par son neveu Ulrich Weiland[5].
Effets sur la cellule

La phalloïdine se lie spécifiquement à l'interface entre les sous-unités d'actine F, bloquant les sous-unités adjacentes ensemble. Cet heptapeptide bicyclique qui se lie plus facilement aux filaments qu'aux monomères d'actine, conduit à la diminution de la constante de vitesse de dissociation des sous-unités d'actine à la fin du filament, ce qui le stabilise en prévenant sa dépolymérisation[6]. De plus, la phalloïdine inhibe l'activité hydrolysase d'ATP de l'actine F[7]. Ainsi, la phalloïdine piège les monomères d'actine dans une conformation distincte de l'actine G et elle stabilise la structure de l'actine F en réduisant de façon importante la constante de vitesse pour la dissociation des monomères, un phénomène associé au piégeage de l'ADP[7]. Globalement, la phalloïdine se trouve réagir stœchiométriquement avec l'actine, promouvant sa polymérisation et stabilisant les polymères [8].
Cette toxine a des effets différents selon sa concentration dans la cellule. Lorsqu'elle est introduite dans le cytoplasme à de faibles concentrations, la phalloïdine recrute les formes les moins polymérisées d'actine cytoplasmique, ainsi que les filaments, dans des « îlots » stables d'agrégats de polymères, mais elle n'interfère pas avec les câbles de tension, c'est-à-dire les épais faisceaux de microfilaments[8]. Wehland et al. notèrent également qu'à de plus fortes concentrations, la phalloïdine induit une contraction cellulaire.
Utilisation en imagerie

Les propriétés de la phalloïdine font d'elle un outil intéressant dans l'étude de la distribution d'actine F dans les cellules en la couplant avec des analogues fluorescents et en les utilisant pour marquer les filaments d'actine en microscopie optique. Les dérivés fluorescents de la phalloïdine se sont avérés extrêmement utiles dans la localisation des filaments d'actine dans les cellules vivantes ou fixées, ainsi que dans la visualisation d'un filament d'actine individuel in vitro[6].

Une technique a été développée pour détecter l'actine F à la fois par microscopie optique (ou photonique) et par microscopie électronique en utilisant la phalloïdine conjuguée à l'éosine qui agit comme un marqueur fluorescent[9]. Dans cette méthode connue sous le nom de fluorescence photo-oxydative, des molécules fluorescentes peuvent être utilisées pour conduire à une oxydation de la diaminobenzidine (DAB) afin de créer un produit de réaction qui est dense aux électrons et détectable par un microscope électronique[9]. La quantité de fluorescence observée peut être considérée comme une mesure quantitative de la quantité de filaments d'actine présents dans la cellule si l'on utilise une quantité saturante de phalloïdine fluorescente[6].
Par conséquent, la microscopie immunofluorescente couplée avec des microinjections de phalloïdine peut être utilisée pour évaluer les fonctions directes ou indirectes de l'actine cytoplasmique à différents stades de la formation du polymère (Wehland et al., 1977). C'est pourquoi la phalloïdine fluorescente peut être utilisée comme un outil important dans l'étude des réseaux d'actine à haute résolution.
Limites des phalloïdines
Les phalloïdines passent difficilement à travers les membranes cellulaires, ce qui les rend moins efficaces dans les expériences utilisant des cellules vivantes. De plus, les cellules traitées à la phalloïdine présentent un nombre important d'effets toxiques et meurent fréquemment[6]. Finalement, les cellules exposées à la phalloïdine présentent un niveau plus important d'actine associée à leurs membranes plasmatiques, et les microinjections de phalloïdine dans une cellule vivante changent la distribution de l'actine ainsi que la mobilité cellulaire[6].
Voir aussi
Notes et références
- Masse molaire calculée d’après « Atomic weights of the elements 2007 », sur www.chem.qmul.ac.uk.
- (en) T. Wieland; V.M. Govindan (1974) Phallotoxins bind to actins. FEBS Lett. 46 (1): 351-3. DOI:10.1016/0014-5793(74)80404-7.
- (en) C. Karlson-Stiber, H. Persson (2003) "Cytotoxic fungi - an overview". Toxicon 42 (4): 339-49. DOI:10.1016/S0041-0101(03)00238-1.
- (en) W. Litten (mars 1975). The most poisonous mushrooms. Scientific American 232 (3): 90–101.
- (en) Feodor Lynen, Ulrich Wieland (1938) Über die Giftstoffe des Knollenblätterpilzes ; Justus Liebig's Annalen der Chemie 533 (1): 93-117. doi:
- (en) JA. Cooper JA (1987) Effects of Cytochalasin and Phalloidin on Actin. J. Cell Biol. 105 (4): 1473-1478.
- (en)JA. Barden, M. Miki, BD. Hambly et CG. Dosremedios (1987) Localization of the phalloidin and nucleotide-binding sites on actin. Eur. J. Biochem. 162 (3): 583-588.
- (en) J. Wehland, M. Osborn et K. Weber (1977) Phalloidin-induced actin polymerization in the cytoplasm of cultured cells interferes with cell locomotion and growth. Proc. Natl. Acad. Sci. 74(12): 5613–5617.
- (en) F. Capani, TJ. Deerinck, MH. Ellisman, E. Bushong, M. Bobik et ME. Martone (2001) Phalloidin–Eosin Followed by Photo-oxidation: A Novel Method for Localizing F-Actin at the Light and Electron Microscopic Levels. J Histochem Cytochem. 49(11): 1351-1362.
 Portail de la mycologie
Portail de la mycologie  Portail de la chimie
Portail de la chimie  Portail de la biochimie
Portail de la biochimie