Granulocyte éosinophile
Les granulocytes éosinophiles ou polynucléaires éosinophiles (ou plus simplement « éosinophiles ») sont des cellules sanguines de la lignée blanche (ou leucocytes). Présentes chez les chordés, elles sont impliquées dans le système immunitaire inné, notamment dans les phénomènes d'allergie et de parasitisme.

Ils sont apparentés aux neutrophiles. Les éosinophiles sécrètent de nombreux composés toxiques et des radicaux libres permettant également l'élimination des bactéries et des parasites, mais qui peuvent occasionner des lésions tissulaires.
Histoire
Les éosinophiles sont découverts chez l'humain par Paul Ehrlich en 1879, qui les mentionne pour la première fois avec leur affinité tinctoriale[1]. Dès la fin du XIXe siècle, des auteurs notent un afflux de ces cellules dans les tissus présentant un processus inflammatoire[2].
Après les observations d'Elie Metchnikoff sur des processus inflammatoires comme la phagocytose et le rôle des globules blancs, l'éosinophile est apparu comme relativement mineur (activité phagocytaire faible), le plus souvent réduit à un rôle de simple témoin des réactions de défense. On lui attribuait tout au plus un rôle régulateur dans certaines réactions allergiques[2].
En 1974, Anthony Butterworth (en) montre qu'un éosinophile, d'une dizaine de μm de diamètre, pouvait détruire in vitro une cible multicellulaire de 150 à 200 μm, à savoir la larve de Schistosoma mansoni, l'agent de la bilharziose[2].
L'éosinophile est alors devenu une cellule efficace, une « cellule tueuse » impliquée dans la protection immunitaire anti-parasitaire, surtout les helminthiases à pénétration tissulaire. Dans les années qui suivent, les publications sur ce sujet se multiplient[3].
À la fin du XXe siècle, les propriétés bénéfiques de l'éosinophile sont étudiées (activité cytotoxique, relations avec d'autres facteurs de l'immunité cellulaire ou de l'immunité humorale). Mais l'éosinophile a aussi des propriétés néfastes, car il peut dépasser son but ou se retourner contre des cellules de l'hôte et participer activement à certaines maladies, dites « pathologies à éosinophiles »[3].
Terminologie
Les granulocytes ont été appelés polynucléaires en raison d'une erreur historique : du fait du caractère lobé de leur noyau (deux lobes en général), on a longtemps cru que ces cellules possédaient plusieurs noyaux.
Le qualificatif de « granulocyte » provient de la présence, dans le cytoplasme, de granules spécifiques et celui d'« éosinophile » du fait, qu'en microscopie optique, après ajout des colorants vitaux usuels, ces inclusions cytoplasmiques fixent l'éosine (elles sont acidophiles) colorant la cellule en rouge.
Les autres granulocytes sont les granulocytes neutrophiles et basophiles.
Origine
Comme les autres granulocytes, les éosinophiles sont formés à partir de cellules souches hématopoiétiques présentes dans la moelle osseuse. La maturation des cellules myéloïdes éosinophiles est contrôlée par divers facteurs tels que GATA-1, CEBP... La cytokine la plus spécifique de la lignée éosinophile est l'interleukine 5[4].
Les granules cytoplasmiques apparaissent au stade du promyélocyte, et ils deviennent spécifiques (éosinophiles) au stade du myélocyte[5].
Morphologie et répartition
.png.webp)
Leur taille varie de 12 à 14 µm. Ils présentent un noyau bilobé ainsi que de nombreuses granules qui se colorent avec de l'éosine (d'où leur nom).
À leur sortie de la moelle osseuse, les éosinophiles matures sont destinés à se répartir dans tous les tissus. Leur fraction circulante dans le sang est une fraction en transit qui ne représente que 1 % de leur nombre total. Leur durée de passage (demi-vie sanguine) est courte, de 3 à 8 heures, ou 18 heures au maximum[4].
L'éosinophilie circulante sanguine, dans des conditions normales, est de moins de 500 éléments par mm3 en chiffres absolus ; et en pourcentage de 1 à 4 % (par rapport à l'ensemble des globules blancs).
99 % des éosinophiles se situent dans les tissus, pour une durée d'une dizaine de jours. Ils se retrouvent plus particulièrement au niveau des muqueuses de surface, et de zones en contact avec l'environnement (peau, tube digestif, système bronchopulmonaire, urogénital…), mais potentiellement au niveau de tout organe selon l'existence d'un foyer inflammatoire[4],[5].
La membrane des éosinophiles exprime divers récepteurs, spécifiques et non spécifiques, pour diverses chimiokines. La migration vers et la localisation dans les tissus sont ainsi contrôlées par des facteurs chimiotactiques comme l'éotaxine (une chimiokine spécifique), ainsi que par l'expression de molécules d'adhésion cellulaire sur l'endothélium.
Fonctions
Après stimulation antigénique (microorganismes, allergènes, médiateurs de l'inflammation, immunoglobulines…), l'éosinophile est activé en libérant, selon les cas, diverses substances[4].
Rôle antiparasitaire et cytotoxique
L'éosinophile joue un rôle important en s'attaquant aux parasites de l'organisme, sans les phagocyter : ils se fixent dessus, déversent leurs granules qui contiennent des enzymes destinées à les détruire. Les vacuoles contenues dans leur cytoplasme contiennent des substances toxiques pour les parasites, mais parfois aussi contre l'organisme hôte.
Ce sont des protéines cationiques stockées dans les granules cytoplasmiques, et douées d'une activité anti-helminthique. La principale (50 % de ces protéines), responsable du caractère éosinophilique, est la protéine basique majeure MBP Major Basic Protein.
Les autres sont la peroxydase éosinophilique EPO Eosinophil Peroxydase, qui se lie aux microorganismes facilitant leur phagocytose par les macrophages ; la protéine cationique éosinophilique ECP Eosinophil Cationic Protein, neutralise l'action de l'héparine et exerce une cytotoxicité sur les parasites en les fragmentant ; la neurotoxine dérivée de l'éosinophile EDN Eosinophil Derivated Neurotoxin, neutralise le SN des parasites ; l'histaminase, neutralise l'activité de l'histamine ; l'arylsulfatase, neutralise les leucotriènes libérés par les granulocytes basophile.
Ces protéines ont des propriétés cytotoxiques directes en adhérant aux groupes acides des membranes cellulaires, elles ont aussi la capacité d'activer ou de contrôler d'autres populations cellulaires comme les basophiles et mastocytes, les lymphocytes, les plaquettes[4],[5].
Rôle inflammatoire
L'éosinophile peut générer des médiateurs lipidiques, dérivés du métabolisme de l'acide arachidonique.
Ces substances semblent avoir un rôle d'amplification de la réaction allergique avec libération d'histamine par les mastocytes et basophiles, notamment au niveau bronchique (hyperréactivité dans l'asthme) ou cutané (angiœdème)[3]. La persistance d'une inflammation à éosinophiles au sein de la lésion d'allergie indique une pérennité de l'inflammation allergique et de nouvelles sensibilisations.
Les éosinophiles jouent un rôle dans l'allergie, et dans l'inflammation allergique, dont ils sont des cellules clés de régulation. En effet, ils possèdent dans leur granules de l'histaminase, une enzyme qui agit sur l'histamine et la neutralise. De plus, les polynucléaires éosinophiles ont la capacité de phagocyter les complexes Antigène-IgE, responsables de la libération de l'histamine par les polynucléaires basophiles et les mastocytes par fixation sur des récepteurs à immunoglobuline E.
Il existe ainsi une dualité de l'action des éosinophiles : réaction adaptée, physiologique (bénéfique) et réaction inadaptée, pathologique (néfaste)[2].
Éosinophilie
Une augmentation de la population d'éosinophiles est appelée « éosinophilie », que l'on rencontre le plus souvent, de façon modérée, chez les personnes présentant une atopie et des manifestations d'allergie (rhinite allergique, asthme, eczéma, urticaire, allergie médicamenteuse). Une hyperéosinophilie importante, mais transitoire, doit faire envisager une maladie parasitaire, comme certaines helminthiases (celles à pénétration tissulaire).
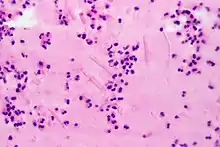
Les autres causes sont les leucémies, cancers, ainsi que des maladies inflammatoires chroniques de la peau et d'autres organes : pemphigus, polymyosite, lupus, périartérite noueuse, sarcoïdose, syndrome de Churg-Strauss...
Dans des situations pathologiques, l'infiltration d'éosinophiles dans les tissus peut se faire au niveau de tout organe, mais préférentiellement dans les tissus cutanés, pulmonaires et myocardiques[4].
La destruction finale des d'amas d'éosinophiles, dans ces conditions pathologiques, se traduit par la formation de cristaux hexagonaux (cristaux de Charcot-Leyden) caractéristiques au microscope optique. Ils ont d'abord été retrouvés dans le sang de leucémiques (Jean Martin Charcot, 1853) et le crachat de certains asthmatiques (Ernst von Leyden, 1872). Dans les années 1980, on les considérait comme formés de lysophospholipase, mais depuis les outils de génétique moléculaire, ce sont des cristaux formés de galectine-10[6] (les galectines appartenant à la famille des lectines animales)[7].
Éosinopénie
L'éosinopénie est une diminution du nombre d'éosinophile dans le sang, en général au dessous de 100 éléments par mm3. Une éosinopénie isolée n'a pas de signification particulière[8].
Selon le contexte clinique et biologique, une éosinopénie pourrait être un marqueur diagnostique et pronostique dans le sepsis et d'autres états pathologiques, mais ces données restent en discussion[9],[10],[11].
Notes et références
- (en) A. B. Kay, « The early history of the eosinophil », Clin. Exp. Allergy, vol. 45, no 3, , p. 575–582 (PMID 25544991, DOI 10.1111/cea.12480)
- Lionel Prin, « Avant-propos », La Revue du Praticien, vol. 50, no 6 « Hyperéosinophilies », , p. 595-596.
- L. Prin, « L'éosinophile aujourd'hui », La Revue du Praticien, vol. 40, no 20, , p. 1866-1872.
- Achille Aouba, « Eosinophilie », La Revue du Praticien, vol. 64, , p. 847-853.
- Sylvain Dubucquoi, « Structure et fonctions des polynucléaires éosinophiles », La Revue du Praticien, vol. 50, no 6, , p. 597-601.
- (en) Jiyong Su, « A Brief History of Charcot-Leyden Crystal Protein Galectin-10 Research », Molecules, (lire en ligne)
- Mireille Viguier, Cyrille Grandjean, Françoise Poirier et Frédérique Deshayes, « Les galectines - Des lectines pas comme les autres », médecine/sciences, vol. 31, no 5, , p. 499–505 (ISSN 0767-0974 et 1958-5381, DOI 10.1051/medsci/20153105011, lire en ligne, consulté le )
- « Laboratoire d'Hématologie Cellulaire du CHU d'Angers », sur www.hematocell.fr (consulté le )
- (en) Alex Smithson, « Is eosinopenia a reliable marker of sepsis? », Critical Care, vol. 13, no 3, (lire en ligne)
- H. Gil, B. Bouldoires, B. Bailly et N. Meaux Ruault, « L’éosinopénie en 2018 », La Revue de Médecine Interne, vol. 40, no 3, , p. 173–177 (ISSN 0248-8663, DOI 10.1016/j.revmed.2018.11.008, lire en ligne, consulté le )
- A. Moussiegt et B. Davido, « Impact diagnostique de l’éosinopénie aux urgences dans le sepsis : les données sont controversées », La Revue de Médecine Interne, vol. 39, no 3, , p. 208–209 (ISSN 0248-8663, DOI 10.1016/j.revmed.2017.12.004, lire en ligne, consulté le )
 Portail de l’hématologie
Portail de l’hématologie