Sélection clonale
La sélection clonale est une théorie scientifique en immunologie qui explique les fonctions des cellules du système immunitaire (lymphocytes) en réponse à des antigènes envahissant le corps.
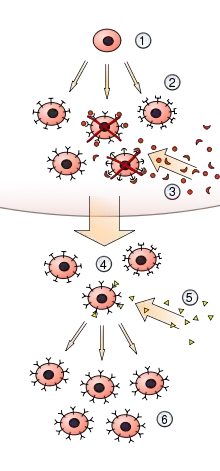
Elle rend compte du mécanisme de sélection des acteurs capables de s'attaquer spécifiquement à l'antigène correspondant à un épitope particulier. Par exemple, les lymphocytes T.
Les acteurs ainsi sélectionnés subissent ensuite une phase de prolifération durant laquelle ils se répliquent à l'identique, menant à la formation d'une population cellulaire dirigée contre l'antigène initialement repéré[1].
Premières approches
En 1900, Paul Ehrlich proposa la "théorie des chaînes latérales" de la production d'anticorps. Selon cette théorie, certaines cellules présentent à leur surface différents "chaîne latérales" (c'est-à-dire des anticorps liés à la membrane) capables de réagir avec différents antigènes. Lorsqu'un antigène est présent, il se lie à une chaîne latérale correspondante. Ensuite, la cellule arrête de produire toutes les autres chaînes latérales et commence une synthèse et une sécrétion intensives de la chaîne latérale de liaison à l'antigène en tant qu'anticorps soluble. Bien que distincte de la sélection clonale, l'idée d'Ehrlich était une théorie de la sélection beaucoup plus précise que les théories instructives qui ont dominé l'immunologie au cours des décennies suivantes.
En 1955, l'immunologiste danois Niels Jerne émis l'hypothèse qu'il existe déjà une vaste gamme d'anticorps solubles dans le sérum avant toute infection. L'entrée d'un antigène dans le corps entraîne la sélection d'un seul type d'anticorps correspondant. Cela se serait produit par certaines cellules phagocytose du complexe immunitaire, répliquant d'une manière ou d'une autre la structure de l'anticorps pour en produire davantage[2].
En 1957, David W. Talmage émis l'hypothèse que les antigènes se lient aux anticorps à la surface des cellules productrices d'anticorps et « seules les cellules sont sélectionnées pour la multiplication dont le produit synthétisé a une affinité pour l'antigène ». La principale différence avec la théorie d'Ehrlich était que chaque cellule était supposée synthétiser une seule sorte d'anticorps. Après la liaison à l'antigène, la cellule prolifère, formant des clones avec des anticorps identiques.
Formulation de la théorie
Le concept de sélection clonale a été introduit par le médecin australien Frank Macfarlane Burnet en 1957, pour tenter d'expliquer la grande diversité des anticorps formés lors de l'initiation de la réponse immunitaire[3],[4]. Dans son article, Burnet développa les idées de Talmage, nommant la théorie résultante « théorie de la sélection clonale ». Il formalisa ensuite cette théorie dans son livre de 1959, « The Clonal Selection Theory of Acquired Immunity ».
La première preuve expérimentale en a été donnée en 1958, lorsque Gustav Nossal et Joshua Lederberg ont montré qu'une cellule B ne produit qu'un anticorps unique[5].
Cette théorie est à présent largement acceptée comme modèle de la façon dont le système immunitaire humain répond à infection et comment certains types de B et lymphocytes T sont sélectionnés pour la destruction de antigènes[6] .
Mécanisme de sélection clonale
Burnet proposa d'expliquer la mémoire immunologique comme étant la conséquence du clonage de deux types de lymphocytes. L'un des clone agit immédiatement pour combattre l'infection, tandis que l'autre dure plus longtemps, restant longtemps dans le système immunitaire et provoquant une immunité contre cet antigène.
Selon l'hypothèse de Burnet, parmi les différents anticorps susceptibles d'être produits se trouvent des molécules qui peuvent probablement correspondre avec des degrés de précision variables à tous, ou pratiquement tous, les déterminants antigéniques qui se produisent dans le matériel biologique autre que ceux caractéristiques du corps lui-même. Chaque type de motif est un produit spécifique d'un clone de lymphocytes. Au cœur de la théorie est l'hypothèse que chaque cellule dispose automatiquement à sa surface de sites réactifs représentatifs équivalents à ceux de la globuline qu'elle produit. Lorsqu'un antigène pénètre dans le sang ou les fluides tissulaires, on suppose qu'il se fixe à la surface de tout lymphocyte portant des sites réactifs qui correspondent à l'un de ses déterminants antigéniques.
La théorie proposée est que dans un groupe préexistant de lymphocytes (en particulier les cellules B), un antigène spécifique active (c'est-à-dire sélectionne) uniquement sa cellule contre-spécifique, qui induit alors cette cellule particulière à se multiplier, produisant un clone pour la production d'anticorps. Cette activation se produit dans les organes lymphoïdes secondaires tels que la rate et les ganglion lymphatiques[7].
De cette manière est initiée une prolifération préférentielle de tous ces clones dont les sites réactifs correspondent aux déterminants antigéniques sur les antigènes présents dans le corps. Les descendants sont capables de libérer activement des anticorps solubles et des lymphocytes, les mêmes fonctions que les formes parentales[8],[9].
En bref, la théorie est une explication du mécanisme de génération de la diversité de la spécificité des anticorps[8].
La théorie de la sélection clonale peut être résumée par les quatre principes suivants :
- Chaque lymphocyte porte un seul type de récepteur avec une spécificité unique (générée par Recombinaison V(D)J).
- L'occupation du récepteur est requise pour l'activation de la cellule.
- Les cellules effectrices différenciées dérivées d'un lymphocyte activé portent des récepteurs de spécificité identique à celle de la cellule mère.
- Les lymphocytes porteurs de récepteurs pour les molécules du soi (c'est-à-dire les antigènes endogènes produits dans le corps) sont détruits à un stade précoce.
Théories supportées par la sélection clonale
Burnet et Peter Medawar ont travaillé ensemble pour comprendre la tolérance immunologique, un phénomène également expliqué par la sélection clonale. Il s'agit de la capacité de l'organisme à tolérer l'introduction de cellules avant le développement d'une réponse immunitaire tant qu'elle se produit tôt dans le développement de l'organisme. Il existe un grand nombre de lymphocytes présents dans le système immunitaire, allant des cellules qui tolèrent les tissus du soi aux cellules qui ne le font pas. Cependant, seules les cellules tolérantes au tissu propre survivent au stade embryonnaire. Si du tissu non-soi est introduit, les lymphocytes qui se développent sont ceux qui incluent les tissus non-soi en tant que tissu propre.
En 1959, Burnet a proposé que dans certaines circonstances, des tissus puissent être transplantés avec succès chez des receveurs étrangers. Ces travaux ont conduit à une bien meilleure compréhension du système immunitaire et également à de grandes avancées dans la transplantation de tissus. Burnet et Medawar ont partagé le Prix Nobel de physiologie ou médecine en 1960.
En 1974, Niels Kaj Jerne a proposé que le système immunitaire fonctionne comme un réseau régulé via des interactions entre les parties variables des lymphocytes et leurs molécules sécrétées. La théorie des réseaux immunitaires est fermement basée sur le concept de sélection clonale. Jerne a remporté le prix Nobel de physiologie ou médecine en 1984, en grande partie pour ses contributions à la théorie des réseaux immunitaires.
L'idée s'est avérée être le fondement de l'immunologie moléculaire, en particulier dans l'immunité adaptative[10].
Notes et références
- F. M. Burnet, « A modification of Jerne's theory of antibody production using the concept of clonal selection », CA: a cancer journal for clinicians, vol. 26, no 2, , p. 119–121 (ISSN 0007-9235, PMID 816431, lire en ligne, consulté le )
- F. M. Burnet, « A Modification of Jerne's Theory of Antibody Production using the Concept of Clonal Selection », CA: A Cancer Journal for Clinicians, vol. 26, no 2, , p. 119–121 (ISSN 0007-9235, PMID 816431, DOI 10.3322/canjclin.26.2.119
 , S2CID 40609269)
, S2CID 40609269) - FM Burnet, « A modification of Jerne's theory of antibody production using the concept of clonal selection. », CA: A Cancer Journal for Clinicians, vol. 26, no 2, , p. 119–21 (PMID 816431, DOI 10.3322/canjclin.26.2.119 , S2CID 40609269)
- Melvin Cohn, Av Mitchison, N., Paul, William E., Silverstein, Arthur M., Talmage, David W. et Weigert, Martin, « Reflections on the clonal-selection theory », Nature Reviews Immunology, vol. 7, no 10, , p. 823–830 (PMID 17893695, DOI 10.1038/nri2177, S2CID 24741671)
- G. J. V. Nossal et Lederberg, Joshua, « Antibody Production by Single Cells », Nature, vol. 181, no 4620, , p. 1419–1420 (PMID 13552693, PMCID 2082245, DOI 10.1038/1811419a0)
- Klaus Rajewsky, « Sélection clonale et apprentissage dans le système d'anticorps », Nature, vol. 381, no 6585, , p. 751–758 (ISSN 0028-0836, PMID 8657279, DOI 10.1038/381751a0, S2CID 4279640)
- Kenneth Murphy, Janeway's Immunobiology 8th Edition, New York, NY, Garland Science, (ISBN 9780815342434)
- Margaret A Jordan et Baxter, Alan G, « Quantitative and qualitative approaches to GOD: the first 10 years of the clonal selection theory », Immunology and Cell Biology, vol. 86, no 1, , p. 72–79 (PMID 18040281, DOI 10.1038/sj.icb.7100140, S2CID 19122290)
- Philip D Hodgkin, Heath, William R et Baxter, Alan G, « The clonal selection theory: 50 years since the revolution », Nature Immunology, vol. 8, no 10, , p. 1019–1026 (PMID 17878907, DOI 10.1038/ni1007-1019, S2CID 29935594)
- R. Medzhitov, « Pattern Recognition Theory and the Launch of Modern Innate Immunity », The Journal of Immunology, vol. 191, no 9, , p. 4473–4474 (PMID 24141853, DOI 10.4049/jimmunol.1302427 )
 Portail de la biologie
Portail de la biologie  Portail de la médecine
Portail de la médecine