Sénescence
En biologie, la sénescence (du latin : senex, « vieil homme » ou « grand âge ») est un processus physiologique qui entraîne une lente dégradation des fonctions de la cellule (notion spécifique de sénescence cellulaire) à l'origine du vieillissement des organismes. Par extension, les biologistes parlent de processus de sénescence des organes et des organismes[1]. La relation entre la sénescence cellulaire et le vieillissement de l'organisme est encore mal comprise[2]. Depuis les années 2000, on sait que la sénescence cellulaire intervient de façon positive dans l'embryogenèse et la réparation tissulaire. Ses effets sur l'oncogenèse sont antagonistes (c'est-à-dire qu'elle constitue un mécanisme naturel de limitation du cancer).
Cet article concerne le vieillissement pour son aspect biologique. Pour les aspects sociaux, culturels, et sociaux économiques, voir Vieillissement.
Le vieillissement existe pour la plupart des espèces animales et végétales, mais pas pour toutes. De très rares espèces ne manifestent qu'une sénescence négligeable, voire aucun vieillissement observable. Certaines sont même capables d'inverser leur processus de vieillissement et de retourner à l'état larvaire — réalisant ainsi une forme d'immortalité biologique. On peut citer notamment le krill et certains cnidaires tels que l'espèce de méduse Turritopsis nutricula et certaines hydres[3]. Chez les plantes ligneuses pérennes, les botanistes observent le maintien d'une ontogenèse permanente (c'est-à-dire que la croissance de la plante ne cesse jamais). Ainsi, si des arbres et arbrisseaux (tel que Lomatia tasmanica) sont caractérisés par un vieillissement physiologique qui peut conduire à la sénescence, ils gardent « leur originalité dans deux domaines : la totipotence de la cellule somatique et le secret, au travers du renouvellement des méristèmes, d'une éventuelle éternité »[4].
Le vieillissement d'un organisme débute après la phase de maturité, et progresse alors de façon irréversible jusqu'à la mort. Il se caractérise le plus souvent par une dégradation des capacités générales de l'organisme : psychomotrices, immunitaires ou reproductives.
La sénescence chez les plantes (en) peut ne toucher qu'une seule partie d'un organisme. C'est le cas de la sénescence des feuilles par exemple qui se caractérise par leur jaunissement puis leur chute en automne, ou encore des fruits lorsqu'ils tombent de la plante.
Causes
La question du vieillissement a longtemps intrigué les philosophes et les naturalistes, tant il est difficile de lui trouver une explication sur le plan biologique.
Il existe un point de vue assez répandu tendant à considérer le vieillissement comme un long processus d'usure physique des tissus qui serait de même nature que l'usure de la matière inerte par le simple effet du passage du temps, c'est-à-dire par application du second principe de la thermodynamique.
D'autres approches concernent la physiologie cellulaire, notamment par l'étude des phénomènes d'oxydation (théorie mitochondriale ou théorie des radicaux libres de Denham Harman) ou d'usure des télomères (théorie télomérique du vieillissement d'Alekseï Olovnikov).
Aucune de ces explications ne résiste à une analyse détaillée, ne rendant notamment pas compte des différences de longévité qui peuvent exister entre différentes espèces.
Théories évolutionnistes

Selon certaines théories évolutionnistes, le vieillissement serait une cause indirecte du fait que la longévité en elle-même n'est en général pas un critère de sélection naturelle. Un caractère procurant un avantage du point de vue de la survie ne peut pas être transmis à la descendance s'il se manifeste lorsque le cycle reproductif de l'organisme est terminé. Lorsqu'il existe en outre des facteurs pathogènes extérieurs (par exemple la prédation), un caractère favorable agissant tardivement n'a alors statistiquement que peu d'influence sur l'espérance reproductive de l'organisme. Par conséquent, les gènes agissant tard dans le développement de l'organisme ne subissent que peu de pression sélective, de telle sorte que les gènes dont l'expression est défectueuse ou délétère, sont plus nombreux lorsque cette expression intervient à un âge avancé que lorsque cette expression intervient à un jeune âge.
Cette idée est illustrée par l'observation du fait que lorsque les facteurs pathogènes extrinsèques sont absents ou inversement corrélés avec l'âge, l'espèce a d'autant plus de chances de ne pas manifester de vieillissement, comme c'est le cas pour plusieurs espèces de grandes tortues, d'esturgeons ou de grands arbres, pour lesquelles la résistance aux prédateurs augmente avec la taille et donc l'âge ; ces espèces présentent une sénescence négligeable. Une autre illustration, pour le cas inverse, est visible chez certaines espèces dont la mort survient de façon spectaculaire après l'accouplement chez les animaux (cf. saumon), ou après la floraison chez les végétaux (cf. bambou). Ce type de reproduction est appelé « sémelparité ». La sémelparité est rarement observée parmi les mammifères, mais elle existe bel et bien, notamment chez certains marsupiaux. L'explication évolutionniste du vieillissement consiste donc dans l'idée d'une continuité entre sémelparité et itéroparité, impliquant que l'itéroparité n'est généralement pas absolue.
Une autre manière d'expliquer cette idée est de l'énoncer selon le point de vue du gène égoïste, concept imaginé par Richard Dawkins et selon lequel « une poule, ça n'est jamais que le moyen qu'a trouvé un œuf pour faire un autre œuf. » Selon ce point de vue une poule n'est pas conçue pour survivre plus longtemps qu'il ne lui est nécessaire pour pondre. Dans cette optique, l'organisme n'est qu'un outil à usage unique, un soma jetable, pour reprendre l'expression de Thomas Kirkwood, dont la seule fonction est de permettre la reproduction et la diffusion des cellules germinales.
La théorie évolutionniste s'applique aussi au niveau cellulaire. Au cours de leur vie, les cellules subissent de nombreux stress et lésions qui peuvent endommager l'ADN. Pour survivre et maintenir leur patrimoine génétique intact, elles ont développé au cours de l'évolution des systèmes de réparation de l'ADN ou de mécanisme d'apoptose (mort cellulaire programmée) qui élimine des cellules dont le génome est lésé ou qui sont potentiellement cancéreuses. Avec l'âge, ces mécanismes de sauvegarde du génome deviennent moins efficaces, d'où le processus de sénescence réplicative qui bloque irréversiblement la prolifération cellulaire[5]. Cette idée de la « sénescence forcée » qui s'applique par exemple aux grains de beauté représente ainsi une voie qui a évolué pour restreindre le développement des cancers dans les organismes supérieurs[6]. « La sénescence cellulaire est un phénomène normal pour les cellules humaines et est caractérisée par un arrêt irréversible de la division cellulaire en phase G 1, qui est associée à des changements morphologiques et fonctionnels de la cellule. C'est un mécanisme anti-tumoral très puissant[7] sélectionné par l'évolution »[8].
Compétition intercellulaire
Selon deux biologistes évolutionnistes de l'Université d'Arizona, le vieillissement est inéluctable pour les organismes multicellulaires, et ce compte tenu de l'antagonisme entre la nécessité d'empêcher la prolifération des cellules cancéreuses, et celle de permettre la croissance et le maintien en vie des autres cellules[9].
Régulation génétique
Chez les végétaux, loin d'être une simple dégradation des conditions de vie de la cellule, voire une simple dégénérescence de celle-ci, la sénescence est un processus contrôlé génétiquement. En effet, certains gènes s'expriment uniquement au moment de la sénescence tandis que d'autres deviennent silencieux.
Sénescence cellulaire
La sénescence cellulaire a été décrite pour la première fois in vitro par Leonard Hayflick en 1961 en étudiant des fibroblastes primaires (WI-38). Il a observé qu'en culture, ces fibroblastes prolifèrent pendant un nombre limité de doublements de population avant de rester bloqués irréversiblement dans un plateau d’arrêt de croissance[10]. Ce plateau nommé limite de Hayflick ou plateau de sénescence réplicative est observé pour la plupart des cellules normales (ex. : fibroblastes, cellules endothéliales…) mais pas pour les cellules cancéreuses.
Phénotype des cellules sénescentes
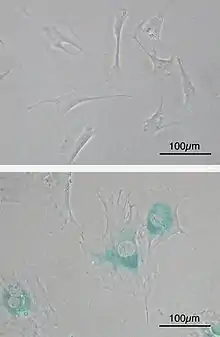
En culture in vitro, les cellules sénescentes développent certains phénotypes spécifiques permettant de les identifier. Parmi ces phénotypes associés à la sénescence on retrouve :
- une augmentation de la taille des cellules et étalement sur leur support ;
- augmentation de la taille des noyaux, parfois accompagnée d’une polynucléation et/ou la formation de micronoyaux ;
- changements dans la structure de la chromatine (foyers d’hétérochromatine, Senescence Associated Hetero-chromatine Foci (SAHF) ;
- augmentation de l’activité lysosomiale pouvant être associé à une augmentation des activités autophagiques ;
- accumulation d’agrégats lipido-protéiques (lipofuscine) ;
- une résistance accrue à l'apoptose ;
- Métabolisme très actif
- La sécrétion de nombreux facteurs pro-inflammatoires formant le SASP (Senescence-Associated Secretory phénotype)[11],[12].
Le marquage de l’activité de la Senescence-Associated β-galactosidase (SA-βgal) est le plus couramment utilisé pour identifier des cellules sénescentes[13]. La β-galactosidase normalement active à pH 4,5 dans les lysosomes est capable de dégrader le composé X-gal dans des conditions non optimales (pH 6) dans les seules cellules sénescentes du fait de leur forte activité lysosomiale [14]. Bien que certaines études montrent que l’on puisse obtenir un marquage positif avec des cellules en forte confluence (Severino et al. 2000)[source insuffisante], le SA-βgal reste le marqueur le plus souvent utilisé pour identifier des cellules sénescentes in vitro comme in vivo.
Mécanisme moléculaire de la sénescence cellulaire
La principale conséquence moléculaire de la sénescence est un blocage du cycle cellulaire à la transition entre les phases G1 et phases S (phase de réplication de l’ADN).
Il existe 2 principales catégories de sénescence cellulaire : la sénescence réplicative et la sénescence prématurée induite par des stress génotoxiques. Ces stress peuvent être de natures génétiques (activation d’oncogène), métaboliques (stress oxydant) ou être environnementaux (drogues cytotoxiques). Bien qu’il y ait des similarités phénotypiques et moléculaires, les sénescences réplicative et prématurée présentent des différences importantes.
Sénescence réplicative
Il s'agit de la sénescence cellulaire dépendant de la taille des Télomères. Ce sont les extrémités des chromosomes, composés de centaines d'éléments nucléotidiques répétitifs (TTAGGG)n, placés en tandem. Leur rôle est principalement protecteur. Leur taille est de 5 à 15 kb (kilobases) chez l'Homme. Cette séquence télomérique se raccourcit à chaque division cellulaire (environ 50 à 200 paires de bases par mitose). Leur longueur reflète le nombre de division d'une cellule, et plus une cellule a eu de divisions, plus la taille des télomères est courte. Ces extrémités sont non répliquées par l'ADN polymérase. La réduction télomérique est une caractéristique du vieillissement cellulaire et de la sénescence.
On observe une longueur limite chez l'être humain de 4,5 kilobases. Une fois cette limite atteinte, il n'y a plus de division cellulaire (activation de la p53 qui produit p21 et arrête le cycle).
La Télomérase est une transcriptase inverse qui allonge l'ADN télomérique. Quand une cellule est dite télomérase positive, elle échappe au phénomène de sénescence. C'est le cas des cellules qui se divisent beaucoup (couche basale de la peau, progéniteurs des lignées sanguines…). Une cellule n'exprimant plus la télomérase va réduire ses télomères et atteindre, au fil des mitoses, la limite de Hayflick[15].
Sénescence induite par des stress oncogéniques
Les cellules non réplicatives et les cellules ayant atteint la limite réplicative de Hayflick font l'objet d'une sénescence post-mitotique, qui peut être accélérée sous l'effet de différents facteurs (stress oxydatif, facteurs métaboliques, agents génotoxiques). Dans ce dernier cas, on parle de « sénescence prématurée induite par le stress » (SIPS)[16].
Elimination des cellules sénescentes par le système immunitaire
Lorsqu'une cellule est sénescente, elle exprime le SASP qui peut activer une réponse immunitaire de l'organisme contre ces cellules sénescentes. Mais ce n'est pas le cas de toutes les cellules sénescentes : certaines persistent dans les tissus, induisant une inflammation et une immunosuppression.
Les cellules dites natural killer (NK) et les macrophages sont les principales cellules responsables de l'élimination des cellules sénescentes. Les cellules NK jouent un rôle de sentinelle permettant la détection de cellules stressées, à l'image des cellules sénescentes. Ces cellules NK libèrent des cytokines qui permettent le recrutement des macrophages et neutrophiles. Ceux-ci peuvent éliminer les cellules sénescentes par phagocytose[17].
Lors de la vieillesse on observe une accumulation de cellules sénescentes qui pourrait être en partie expliquée par une diminution des cellules NK dans l'organisme dû à l'immunosénescence[18],[19],[20].
Sénescence et cancer
La sénescence cellulaire a des effets antagonistes dans l'oncogenèse, elle limite l'apparition de certains cancers en bloquant la division cellulaire de cellules précancéreuses ou en supprimant des tumeurs. Elle peut en outre avoir un effet activateur, la prolifération de cellules sénescentes, lorsqu'elles ne sont pas détruites par des cellules immunitaires, ouvrant la voie à l'apparition de cancers[21].
Sénescence et climat
Quelques espèces de milieux extrêmes entrent en dormance lors de stress thermiques intenses (température trop haute ou trop basse pour leur métabolisme), mais pour la plupart des autres espèces, ces stress sont des facteurs d'accélération de la sénescence ou de mortalité.
Chez les ectothermes, les variations thermiques influent sur le taux de sénescence des adultes en fonction des conditions climatiques[22]. Ceci a été par exemple été étudié chez des amphibiens par la technique de capture-recapture sur le long terme, au sein de plusieurs populations d'amphibiens de deux paires d'espèces d'anoures apparentées choisies dans deux familles ayant divergé il y a plus de 100 Mya : dans la famille des Ranidae (Rana luteiventris et Rana temporaria) et dans la famille des Bufonidae (Anaxyrus boreas et Bufo bufo) largement réparties dans l'hémisphère nord[22]. En outre, chez deux de ces espèces (R. luteiventris et A. boreas), ni les précipitations annuelles moyennes, ni l'empreinte environnementale humaine n'ont eu des effets significatifs sur les taux de sénescence ou la durée de vie, alors que les conditions thermiques de vie, en région tempérée, affectent le risque de mortalité par vieillissement précoce laissant craindre que le réchauffement climatique cause dans ces régions une accélération généralisée du vieillissement des amphibiens, déjà en fort déclin mondial[22].
Rôle de la sénescence dans l'embryogenèse et la réparation tissulaire
Les cellules sénescentes jouent un rôle actif dans l'embryogenèse, aussi bien chez la souris, le poulet, que chez l'être humain[23]. De la même façon, générées par la protéine CCN1, elles interviennent dans les phénomènes de cicatrisation, de régénérescence tissulaire, et de réduction des fibroses, par exemple hépatiques (cirrhose) ou cardiaques[23].
Notes et références
- Joël Aghion, « L’immortalité cellulaire existe-t-elle ? », Études sur la mort, no 124, , p. 60.
- Jacques Epelbaum, Le vieillissement : rythmes biologiques et hormonaux, Lavoisier, (lire en ligne), p. 13.
- (en) Absurd Creature of the Week: This Amazing Little Critter Just Might Be Immortal, Wired.
- Développement végétal, Encyclopaedia Universalis, , 60 p. (lire en ligne).
- Maurice Tubiana, La science au cœur de nos vies, Odile Jacob, (lire en ligne), p. 190.
- Gerald Karp, Biologie cellulaire et moléculaire, De Boeck Supérieur, (lire en ligne), p. 674.
- La sénescence n'est cependant pas un mécanisme parfait d'inhibition de la tumorigenèse car les cellules sénescentes sécrètent in vitro des substances carcinogènes qui peuvent favoriser à long terme le développement de certaines tumeurs.
- Georgia Barlovatz-Meimon, Xavier Ronot, Culture de cellules animales, Lavoisier, (lire en ligne), p. 328.
- (en)Intercellular competition and the inevitability of multicellular aging, Paul Nelson et Joanna Masel
- (en) Hayflick L, Moorhead PS., « The serial cultivation of human diploid cell strains », Exp Cell Res, no 25, , p. 585-621 (PMID 13905658, lire en ligne).
- Jean-Philippe Coppé, Christopher K. Patil, Francis Rodier et Yu Sun, « Senescence-associated secretory phenotypes reveal cell-nonautonomous functions of oncogenic RAS and the p53 tumor suppressor », PLoS biology, vol. 6, no 12, , p. 2853–2868 (ISSN 1545-7885, PMID 19053174, PMCID 2592359, DOI 10.1371/journal.pbio.0060301, lire en ligne, consulté le )
- Nicolas Malaquin, Aurélie Martinez et Francis Rodier, « Keeping the senescence secretome under control: Molecular reins on the senescence-associated secretory phenotype », Experimental Gerontology, vol. 82, , p. 39–49 (ISSN 1873-6815, PMID 27235851, DOI 10.1016/j.exger.2016.05.010, lire en ligne, consulté le )
- (en) Dimri GP, Lee X, Basile G, Acosta M, Scott G, Roskelley C, Medrano EE, Linskens M, Rubelj I & Pereira-Smith O (1995) « A biomarker that identifies senescenthuman cells in culture and in aging skin in vivo » Proc Natl Acad Sci.U.S.A. 92:9363-7. .
- (en) Bo Yun Lee, Jung A. Han, Jun Sub Im et Amelia Morrone, « Senescence-associated β-galactosidase is lysosomal β-galactosidase », Aging Cell, vol. 5, no 2, , p. 187–195 (ISSN 1474-9726, DOI 10.1111/j.1474-9726.2006.00199.x, lire en ligne, consulté le )
- Odélie Tacita, « Les causes biologiques du vieillissement et de la limitation de la durée de vie - work for human longevity », work for human longevity, (lire en ligne, consulté le ).
- Jeanne Mialet-Perez, Victorine Douin-Echinard, Daniel Cussac, Antoine Bril et Angelo Parini, « Vieillissement - Une question de cœur ? », sur inserm.fr, Med Sci, 31 novembre 2015 (DOI 10.1051/medsci/2015311101).
- Larissa G. P. Langhi Prata, Inna G. Ovsyannikova, Tamara Tchkonia et James L. Kirkland, « Senescent cell clearance by the immune system: Emerging therapeutic opportunities », Seminars in Immunology, vol. 40, , p. 101275 (ISSN 1096-3618, PMID 31088710, PMCID 7061456, DOI 10.1016/j.smim.2019.04.003, lire en ligne, consulté le )
- Fabrizio Antonangeli, Alessandra Zingoni, Alessandra Soriani et Angela Santoni, « Senescent cells: Living or dying is a matter of NK cells », Journal of Leukocyte Biology, vol. 105, no 6, , p. 1275–1283 (ISSN 1938-3673, PMID 30811627, DOI 10.1002/JLB.MR0718-299R, lire en ligne, consulté le )
- Eugenia Roupakia, Georgios S. Markopoulos et Evangelos Kolettas, « Genes and pathways involved in senescence bypass identified by functional genetic screens », Mechanisms of Ageing and Development, vol. 194, , p. 111432 (ISSN 1872-6216, PMID 33422562, DOI 10.1016/j.mad.2021.111432, lire en ligne, consulté le )
- Ping Song, Junqing An et Ming-Hui Zou, « Immune Clearance of Senescent Cells to Combat Ageing and Chronic Diseases », Cells, vol. 9, no 3, , E671 (ISSN 2073-4409, PMID 32164335, PMCID 7140645, DOI 10.3390/cells9030671, lire en ligne, consulté le )
- Pierre-François Roux et Oliver Bischof, « La sénescence, une destinée cellulaire aux multiples visages », sur ens.fr,
- (en) Hugo Cayuela, Jean-François Lemaître, Erin Muths et Rebecca M. McCaffery, « Thermal conditions predict intraspecific variation in senescence rate in frogs and toads », Proceedings of the National Academy of Sciences, vol. 118, no 49, , e2112235118 (ISSN 0027-8424 et 1091-6490, DOI 10.1073/pnas.2112235118, lire en ligne, consulté le )
- Pierre-François Roux et Oliver Bischof, « La sénescence, une destinée cellulaire aux multiples visages », sur ens.fr,
Voir aussi
Bibliographie
- Richard Dawkins, Le gène égoïste.
- Frederic Revah et Andre Klarsfeld, Biologie de la mort, Odile Jacob, , 289 p. (ISBN 978-2-7381-0757-2, lire en ligne).
Articles connexes
- Sénescence négligeable
- SENS Le projet initié par Aubrey de Grey qui a pour but l'extension radicale de l'espérance de vie humaine.
- Prix de la Souris Mathusalem (Methuselah Mouse Prize), une récompense pour les recherches visant à l'accroissement radical de l'espérance de vie humaine.
- Vieillesse
- Progéria, maladie présentant des signes de sénescence accélérée chez le jeune enfant
- le cas Brooke Greenberg, un défaut de vieillissement
- Cheveux blancs
Liens externes
(fr)
- Explications détaillées sur la sénescence cellulaire sur herman.denis.club.fr.
- Chronique du livre Serons-nous immortels ? de Ray Kurzweil et Terry Grossman sur schismatrice.net.
- Senescience.fr.
- « Le vieillissement des cellules est réversible » sur lemonde.fr, .
- Article sur jle.com.
(en)
- Mechanisms of Aging sur benbest.com.
- senescence.info Informational website on aging.
- AgeLab (MIT).
- Aging because body loses genetic info sur meucat.com.
- The Longevity Meme (Longevity Activism) sur longevitymeme.org.
- See the artproject Dialogue with the High Age sur willypuchner.com.
 Portail de la médecine
Portail de la médecine  Portail de la sociologie
Portail de la sociologie  Portail de la psychologie
Portail de la psychologie