Supercolonie de fourmis
Une supercolonie de fourmis est une colonie de fourmis, composée de multiples reines fécondes et constituée d'un maillage de zones de nidification interconnectées. Sa structure sociale est flexible, étendue et complexe. Ce type de colonie peut s'étendre sur des centaines de kilomètres[1].

Définition

Une supercolonie est définie, en premier lieu par une forte polygynie, c'est-à-dire par la présence de plusieurs dizaines voire centaines de reines fécondes dans une même fourmilière. En deuxième lieu, elle est polydôme ou polycalique[2], c'est-à-dire que sa structure sociale est caractérisée par une certaine flexibilité et l'interconnexion de plusieurs zones de nidification de différents types et de zones de chasse ou de récolte communes. Par opposition, une colonie monogyne est constituée par une seule reine féconde, et une colonie monodôme, par un système se traduisant par l'établissement d'un territoire indépendant[3].
Les termes « pluricolonie » (ou polycolonie et multicolonie) et « unicolonie » définissent des gradients d'agressivité intraspécifiques, des espèces les plus agressives aux moins agressives. Le terme « supercolonie » étant compris dans cet ensemble[4].
Plus précisément, le terme « supercolonie » est généralement utilisé pour définir de grands rassemblements de fourmilières interconnectées et non agressives entre elles, mais pouvant l'être contre d'autres fourmilières de leur propre espèce (par exemple, Formica paralugubris et Formica yessensis). En revanche, le terme « unicolonie » se limite aux espèces dont tous les individus sont considérés comme étant amicaux (par exemple la Fourmi argentine), tandis que le terme « pluricolonie » définit une espèce dont toutes les colonies sont agressives les unes par rapport aux autres[4].
Types biologiques
Formica yessensis est une espèce du sous-genre Formica (formica), moyennement polygyne et polydôme, réputée pour sa supercolonie située dans la sous-préfecture de Ishikari sur la côte ouest d'Hokkaidō au Japon. Cet ensemble a été estimé à 306 millions d'ouvrières et 1 million de reines vivant sur une zone de 2,7 km² et comprenant 45 000 fourmilières[5].
Formica paralugubris est également une espèce du sous-genre Formica (formica), fortement polygyne et polydôme, connue pour sa supercolonie établie au sein de la station du Chalet à Roch dans le massif du Jura en Suisse. Elle est constituée de 1 200 fourmilières reliées entre elles par 100 km de pistes sur une surface de 70 hectares[3]
Ces supercolonies sont étudiées depuis les années 1970, respectivement par le département zoologie de l'université de Hokkaidō (représenté par Seigo Higashi, Masao Ito et Shinji Imamura)[6] et par le secteur écologie et évolution de l'université de Lausanne (sous la houlette de Daniel Cherix[7]). Elles représentent toutes les deux le type biologique de la supercolonie[4],[8].
Structure sociale
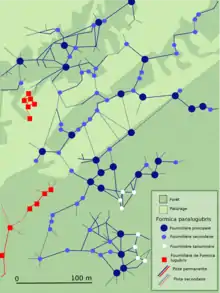
Une supercolonie est constituée de fourmilières principales, possédant chacune une importante population et plusieurs pistes de liaisons qui mènent à des fourmilières secondaires ou saisonnières. Les fourmilières secondaires sont les filles des principales alors que les saisonnières sont utilisées momentanément pour désengorger la principale au moment où les nymphes sont produites en nombre[3]. les fourmilières voisines échangent beaucoup d'ouvrières transportant de la nourriture, des matériaux de construction ou des nymphes. La dérive est importante[9]. D'autres pistes, dites de « fourragement », mènent aux zones de chasse et de récolte[3].
Une supercolonie est constituée de différents secteurs, comprenant chacun quelques fourmilières principales, reliées par des pistes de liaison qui permettent des échanges de nourriture, en cas de disette ou de surplus localisés[3]. Ils sont plutôt génétiquement similaires en leur sein[3] et peuvent présenter de grandes différences génétiques entre eux[10]. Il y a peu d'échanges d'individus de l'un à l'autre ; cependant, un individu d'un secteur donné sera toujours accepté au sein d'un autre secteur[3].
Limites territoriales
La cuticule des fourmis est imprégnée de lipides, essentiellement des hydrocarbures, qui leur confère une identité chimique. Cette reconnaissance identitaire permet aux individus de défendre leur territoire et de prodiguer les soins uniquement aux membres de leur propre colonie. Les phéromones cuticulaires d'une colonie monogyne et monodôme sont spécifiques. À l'inverse, le profil chimique d'une supercolonie est composé d'un mélange des phéromones de tous les individus et est très varié[3].
Afin de connaître les limites d'une supercolonie, le modus operandi des chercheurs est simple. Ils réalisent des « tests d'agressivité » en présentant des ouvrières de la colonie évaluée dans des fourmilières voisines. Tant que les fourmis sont acceptées, elles partagent leur profil cuticulaire et la fourmilière fait ainsi partie de la supercolonie. Si elles sont attaquées, elles ne partagent plus la même identité chimique, la limite de la supercolonie est alors atteinte. De proche en proche, il est alors possible de dresser des cartes territoriales[3].
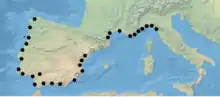
Ainsi, une carte territoriale de la supercolonie de fourmis argentines a été dressée le long de la côte méditerranéenne entre l'Italie, la France, l'Espagne et le Portugal, tout en discriminant une partie des fourmilières catalanes, alors considérées comme une supercolonie distincte. La supercolonie principale mesure plus de 6000 km avec des millions de fourmilières[4]. Cette espèce forme également des supercolonies sur la côte de l'État de Victoria, en Australie (100 km de long)[11] et sur les littoraux californiens[12] et japonais[13]. Elle est directement liée au climat méditerranéen[14] et son extension rapide est directement liée aux activités humaines[13]. Sous la direction de E. Sunamura, des chercheurs ont déplacé des individus européens et californiens afin de tester leur agressivité au Japon. Les ouvrières n'ont montré aucun signe d'agressivité. Les trois supercolonies forment ainsi la plus grande unité non agressive créés par une espèce d’insecte social[13]. Dès lors, certains médias parlent de « mégacolonie » colonisant notre planète[15]. À la stratégie polygyne-polydôme peut donc s'ajouter la stratégie unicoloniale où la solidarité n'est pas uniquement localisée, mais généralisée à l'ensemble de l'espèce. La fourmi argentine est généralement utilisée comme exemple type parfait d'unicolonialité, mais la découverte des deux supercolonies différentes au sein des populations européennes remet en question ce modèle[4].
Stratégie écologique
Une supercolonie, en tant que polygyne (jusqu'à 2 000 reines fécondes par fourmilière), augmente significativement la quantité d'œufs pondus par rapport à une colonie monogyne et peut facilement pallier la mort d'une ou plusieurs reines. Il existe des fourmilières produisant uniquement des sexués femelles et des fourmilières produisant également des sexués mâles. Les fourmilières mixtes permettent un accouplement à l'intérieur ou à la surface du nid. Une femelle peut s'accoupler directement dans sa fourmilière. Certaines vierges découpent même leurs ailes avant fécondation, empêchant tout vol nuptial, coûteux et dangereux. Au moment où les nids sont surpeuplés, des ouvrières et des reines peuvent alors quitter leur fourmilière pour aller en constituer une nouvelle à proximité. Ce phénomène est nommé « bourgeonnement » (phénomène similaire à l'essaimage chez les abeilles domestiques). Les supercolonies sont donc le résultat de bourgeonnements successifs où chaque fourmilière reste connectée par une piste de liaison permanente à sa fourmilière d'origine. La stratégie de colonisation consiste ainsi à investir localement et permet de dominer grâce à un très fort pouvoir de compétition, tout en évitant les dangers des vols nuptiaux. Ainsi aucune autre espèce de fourmi n'a la possibilité de s'établir sur le territoire de la supercolonie. Ce système est adapté à des ressources alimentaires denses. Cette stratégie est toutefois risquée en cas d’événement catastrophique tel qu'un feu de forêt ou une maladie contagieuse. En ce cas, c'est l'ensemble de la colonie qui pourrait disparaître et avec elle une grande partie de l'espèce. C'est pourquoi il y a toujours une faible dispersion des sexués afin de coloniser de nouveaux territoires[3].
Dans une colonie monogyne, la mort de la reine engendre presque inévitablement la disparition de la colonie. D'autre part, le nombre d'œufs pondu est limité et la colonie est composée uniquement de ses propres descendants. D'un point de vue général, une colonie monodôme accepte les coûts d'un comportement agressif. Il en résulte, entre autres, la perte d'ouvrières et de soldates lors des luttes de territoire. Mais la mort d'une colonie ne met pas en danger l'ensemble de l'espèce. Sa stratégie est d'investir dans la dispersion. Pour ce faire, le nombre de sexués femelles est plus important et elles augmentent leur capacité à survivre sans l'aide d'ouvrières grâce à d'importantes réserves de graisse. Certaines espèces pratiquent le parasitisme social temporaire. Une princesse Formica lugubris pourra par exemple parasiter une colonie de Formica lemani en tuant sa reine et en utilisant ses ouvrières pour s'occuper de sa propre descendance. Les monogynes-monodômes peuvent ainsi coloniser plus efficacement de nouveaux sites plus éloignés que les supercolonies. Ce système est adapté à des ressources alimentaires dispersées[3].
Une présence localisée et dense pour les polygynes polydômes (caractéristiques des supercolonies), une présence largement dispersée et en faible densité pour les monogynes monodômes. Ces deux stratégies peuvent cohabiter localement comme le montre la station suisse du Chalet à Roche où la supercolonie de Formica paralugubris cohabite sur ses limites avec Formica lugubris, espèce monogyne à faiblement polygyne et monodôme à faiblement polydôme[3].
Des ouvrières coopératives et des reines antagonistes

Iridomyrmex purpureus est une espèce de fourmi faiblement polygyne, polydôme et fortement pluricoloniale, endémique des déserts arides australiens. Elle forme parfois des supercolonies souterraines dont la plus grande répertoriée à ce jour occupe plus de 10 hectares avec 85 fourmilières et 1 500 trous d’entrée[16]. Elle est constituée de plusieurs fourmilières principales reliées par des pistes de liaison aux fourmilières secondaires. Une fourmilière est composée de plusieurs trous d'entrée amenant à un réseau de galeries indépendant[17]. Les ouvrières coopèrent pour la recherche de nourriture et la récolte de miellat sur les eucalyptus en livrant la majeure partie de leur collecte à la fourmilière la plus proche. Cependant, elles restent fidèles à leur fourmilière d'origine et entretiennent uniquement des larves de leur propre reine. Elles sont particulièrement agressives envers les fourmis de leur propre espèce provenant de colonies éloignées, au point qu'elles peuvent se nourrir des sexués lors de leur vol nuptial[1].
Les modes de fondation des colonies de I. purpureus sont remarquablement diversifiés. Les nouvelles colonies peuvent provenir de fondatrices uniques, d'associations de fondatrices ou de bourgeonnement de colonies. Dans ce cas, des reines nouvellement accouplées creusent des chambres de fondation à côté de fourmilières matures, les ouvrières les protégeant et les aidant à creuser. La coexistence de plusieurs reines dans une même fourmilière est antagoniste. Les reines dominantes inhibent la ponte des subordonnées alors que les ouvrières ignorent ces interactions et traitent les reines de manière égale. À mesure que la colonie se développe, les reines deviennent intolérantes face à la présence des autres et se séparent dans des réseaux de galeries adjacents. Une fois séparées, les reines semblent avoir le même statut, pondant à peu près autant d'œufs. Ce comportement dit d'« oligogynie » est défini par la tolérance des ouvrières envers une multitude de reines et l’antagonisme de ces dernières[18].
Une stratégie précurseure
Lasius austriacus, une espèce du genre Lasius d'Europe centrale, est une monogyne-monodôme. Cependant, elle n'est pas agressive envers les autres colonies de son espèce et intègre occasionnellement des ouvrières étrangères en son sein, tout en conservant son fonctionnement indépendant. L'abandon de l'agression tout en maintenant la distinction entre les colonies pourrait être une première étape dans la formation des supercolonies[19].
Changement de stratégie des fourmis argentines
Les populations natives de fourmi argentine sont polygynes-polydômes-multicoloniales, et les populations invasives unicoloniales. Ce changement de stratégie est au cœur de nombreuses études scientifiques. L'hypothèse première met en avant l'appauvrissement génétique des souches importées, entraînant une réduction de la diversité génétique du locus de reconnaissance phéromonale de la colonie et une perte d'agressivité entre les colonies ayant le même allèle de reconnaissance. Mais le goulot d'étranglement génétique attendu n'est pas si important et ne permet pas d'expliquer l'unicolonialité. De plus, l'agressivité de la supercolonie principale avec la supercolonie catalane est importante, que ce soit in situ ou en laboratoire, même après plusieurs mois. Une autre hypothèse met en avant le « nettoyage génétique ». À cause de l'augmentation des ressources alimentaires et de l'assouplissement des contraintes écologiques, les populations invasives ont une densité de colonies bien plus importante que les populations indigènes. Les colonies présentant les indices de reconnaissance les plus courants bénéficient alors d'un avantage sélectif, car elles combattent moins souvent et sont donc plus productives. Une sélection des allèles de reconnaissance se met en place. Peuvent alors émerger de grandes supercolonies composées de colonies non agressives[4],[10].
Exemples d'espèces de fourmis formant des supercolonies
Notes et références
- (en) Wilson et Hölldobler, The Ants, Belknap Press, 1990, (ISBN 9780674040755)
- Claude Lebas, « polycalique », sur glossaire.fourmis.free.fr
- (fr) Daniel Cherix, Anne Freitag, Arnaud Maeder, Fourmis des bois du Parc jurassien vaudois, 2006, Parc Jurassien vaudois et Musée de zoologie de Lausanne en partenariat avec les Editions Rossolis, (ISBN 9782940365128)
- (en) Giraud, Tatiana, Jes S. Pedersen, and Laurent Kelle, « Evolution of supercolonies: The Argentine ants of southern Europe », National Academy of Sciences, vol. 99, no 9, (résumé)
- (en) Higashi, S. et K. Yamauchi, « Influence of a Supercolonial Ant Formica (Formica) yessensis Forel on the Distribution of Other Ants in Ishikari Coast », Japanese Journal of Ecology, vol. 29, , p. 257-264 (résumé)
- (en) Thèses portant sur Formica yessensis et sa supercolonie de Hokkaidō sur Hokkaido University Collection of Scholarly and Academic Papers
- Daniel Cherix sur Uniscience de l'Université de Lausanne
- Pierre Jaisson, La fourmi et le sociobiologiste, Editions Odile Jacob, 1993, (ISBN 978-2738102072)
- (en) HIGASHI, Seigo, « Analysis of Internest Drifting in a Supercolonial Ant Formica (Formica) yessensis by Individually Marked Workers », Journal of the faculty of science Hokkaido University Series VI. zoologie, vol. 46, no 2, , p. 176-191 (résumé)
- (en) Krista K. Ingram, Deborah M. Gordon, « Genetic analysis of dispersal dynamics in an invading population of argentine ants », Ecology, vol. 84, no 11, (résumé)
- ,(en) Alexei D. Rowles et Dennis J. O'Dowd, « Impacts of the invasive Argentine ant on native ants and other invertebrates in coastal scrub in south‐eastern Australia », Austral Ecology, vol. 34, , p. 239-248 (résumé)
- (en) Nathan J. SandersKasey E. BartonDeborah M. Gordon, « Long-term dynamics of the distribution of the invasive Argentine ant, Linepithema humile, and native ant taxa in northern California », Oecologia, vol. 127, , p. 123-130 (résumé)
- (en) E. Sunamura, X. Espadaler, H. Sakamoto, S. Suzuki, M. Terayama, S. Tatsuki, « Intercontinental union of Argentine ants: behavioral relationships among introduced populations in Europe, North America, and Asia », Insectes Sociaux, vol. 56, no 2, (résumé)
- (en) Ellen Van Wilgenburg, Candice W Torres, and Neil D Tsutsui, « The global expansion of a single ant supercolony », Evolutionnary applications, vol. 3(2), , p. 136–143 (résumé)
- Matt Walker, « Ant mega-colony takes over world », Earth News, BBC News, (lire en ligne [archive du ], consulté le )
- (en) Greenslade, P. J. M.; Halliday, R. B., « Colony dispersion and relationships of meat ants Iridomyrmex purpureus and allies in an arid locality in south Australia », Insectes Sociaux, vol. 30, , p. 82-99 (résumé)
- (en) Ettershank G., « The three dimensional gallery structure of the nest of the meat ant Iridomyrmex purpureus (SM.) (Hymenoptera : Formicidae) », Australian Journal of Zoology, vol. 16, , p. 715-723 (résumé)
- (en) Bert Hölldobler, Norman F. Carlin, « Colony founding, queen dominance and oligogyny in the Australian meat ant Iridomyrmex purpureus », Behavioral Ecology and Sociobiology, vol. 18, , p. 45-58 (résumé)
- (en) Florian M. Steiner, Birgit C. Schlick-Steiner, Karl Moder, Christian Stauffer, Wolfgang Arthofer, Alfred Buschinger, Xavier Espadaler, Erhard Christian, Katrin Einfinger, Eberhard Lorbeer, Christa Schafellner, Manfred Ayasse, Ross H. Crozier, « Abandoning aggression while maintaining self-nonself discrimination as a first stage in ant supercolongy formation. », Current biology, vol. 21, (résumé)
Articles connexes
 Portail de l’entomologie
Portail de l’entomologie