Surenroulement de l'ADN
Le surenroulement de l'ADN résulte de la présence de supertours positifs (dans le sens de la double hélice) ou négatifs (dans le sens inverse) sur une double hélice d'ADN qui imposent une contrainte sur cette molécule. Le surenroulement, qui correspond mathématiquement à la torsade d'un ruban, joue un rôle important dans un grand nombre de processus biologiques, notamment la condensation de l'ADN. Il module en effet l'entortillement de l'ADN, la somme de la torsade et de l'entortillement correspondant à l'enlacement de la molécule, lequel a un effet sur le volume occupé par cette dernière. Par ailleurs, certaines enzymes telles que les ADN topoisomérases sont capables de modifier la topologie de l'ADN pour faciliter certains processus tels que la réplication de l'ADN ou sa transcription en ARN. D'une manière générale, l'ADN de la plupart des êtres vivants est surenroulé négativement[1].
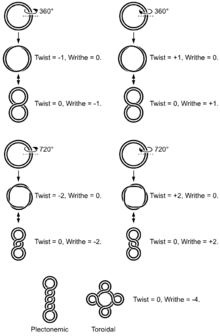
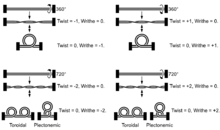
Conformations
Dans un segment d'ADN B relâché, les deux brins de la double hélice s'enroulent l'un autour de l'autre avec un pas constitué d'environ 10,4 à 10,5 paires de bases. Ajouter ou retirer des tours à ce segment d'ADN, comme certaines enzymes sont capables de le faire, lui impose une contrainte mécanique. Dans le cas d'un ADN circulaire, l'introduction de supertours successifs conduit à différentes conformations de la molécule. Chacune de ces conformations est caractéristique d'un surenroulement de l'ADN.
Le surenroulement le plus simple de l'ADN circulaire est en forme de 8. Il résulte de l'introduction d'un seul supertour, positif ou négatif. Les deux lobes du 8 sont tordus dans le sens des aiguilles d'une montre sous l'effet d'un supertour positif, et dans le sens inverse de celui des aiguilles d'une montre sous l'effet d'un supertour négatif. L'introduction de nouveaux supertours, positifs ou négatifs, conduit à une torsion supplémentaire entre les deux lobes de l'ADN circulaire.

L'ADN linéaire peut également être surenroulé, en raison notamment de la très grande taille des chromosomes, qui empêche de relâchement complet des contraintes localisées sur un segment d'ADN donné.
L'ADN surenroulé est susceptible de former une combinaison de deux types de structures : des toroïdes et des plectonèmes.
- Un toroïde est une succession de boucles distinctes le long d'un segment d'ADN surenroulé.
- Un plectonème est une accumulation de boucles localisées en un point du segment d'ADN surenroulé conduisant à une structure torsadée. Dans le cas d'un ADN linéaire, le plectonème se développe dans une direction perpendiculaire au segment d'ADN surenroulé ; dans le cas d'un ADN circulaire, il forme une torsade scindant la boucle en deux lobes. C'est la conformation la plus fréquente in vivo.
Occurrence

Le surenroulement de l'ADN est important pour son conditionnement au sein des cellules. Une molécule d'ADN pouvant être mille fois plus longue que le diamètre d'une cellule, sa condensation pour qu'elle occupe un espace à la mesure de celui d'une cellule est une opération critique. C'est à ce niveau qu'intervient le surenroulement de l'ADN, dans la mesure où il en réduit sensiblement l'encombrement. Chez les procaryotes, les plectonèmes sont prédominants en raison de la géométrie généralement circulaire des chromosomes et de la quantité plutôt limitée du matériel génétique dans ces cellules. Chez les eucaryotes, on trouve une combinaison de structures à la fois plectonémiques et solénoïdales, ces dernières offrant la méthode la plus efficace pour rendre l'ADN plus compact. Cette conformation est réalisée au sein des chromosomes d'eucaryotes à l'aide d'histones, protéines basiques qui forment, avec l'ADN, une fibre d'environ 10 nm de diamètre.
Les protéines SMC interviennent notamment lors de la division cellulaire, qu'il s'agisse de mitose ou de méiose, sous la forme de condensines et de cohésines qui favorisent la condensation des chromatides sœurs et la liaison du centromère en introduisant des supertours positifs.
Le surenroulement de l'ADN intervient également lors de la réplication de l'ADN et de sa transcription en ARN. Il résulte du déroulement de la double hélice par l'ADN polymérase et l'ARN polymérase respectivement, déroulement qui introduit des supertours positifs en amont et négatifs en aval. Des ADN topoisomérases telles que l'ADN gyrase interviennent pour relâcher en partie ce surenroulement[2].
Notes et références
- (en) James J. Champoux, « DNA TOPOISOMERASES: Structure, Function, and Mechanism », Annual Review of Biochemistry, vol. 70, , p. 369-413 (PMID 11395412, DOI 10.1146/annurev.biochem.70.1.369, lire en ligne)
- (en) Annie-Claude Albert, Flavia Spirito, Nara Figueroa-Bossi, Lionello Bossi et A. Rachid Rahmouni, « Hyper-Negative Template DNA Supercoiling During Transcription of the Tetracycline-Resistance Gene in topA Mutants is Largely Constrained In Vivo », Nucleic Acids Research, vol. 24, no 15, , p. 3093-3099 (PMID 8760899, PMCID 146055, DOI 10.1093/nar/24.15.3093, lire en ligne)
 Portail de la biologie cellulaire et moléculaire
Portail de la biologie cellulaire et moléculaire