Vacuole
La vacuole est un organite présent dans la cellule végétale, les pollens, les cellules fongiques et plus généralement toutes les cellules des différentes lignées d’eucaryotes photosynthétiques[1] et dans certaines cellules animales. Les vacuoles sont des compartiments délimités par une membrane (tonoplaste), remplis d'eau et contenant diverses molécules inorganiques et organiques telles que des enzymes. La vacuole n'a pas de forme ou de taille particulière, sa structure variant en fonction des besoins de la cellule. La vacuole se différencie de la vésicule par sa taille et sa fonction, malgré des similitudes au niveau des fonctions.
Cet article concerne les vacuoles en biologie. Pour les vacuoles en géologie, voir Vacuole (géologie).
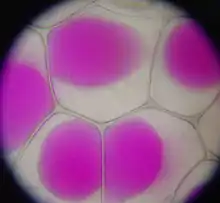
La fonction et l'importance des vacuoles varient selon le type de cellule dans lesquelles elles sont présentes. En général, ces fonctions sont liées à des mécanismes de défense (détoxication, protection) qui comprennent :
- L'isolement de composant potentiellement nocif pour la cellule
- La gestion des déchets à l'aide d'enzyme de digestion ou l'endocytose des organites vieillis
- Le maintien de l'équilibre hydrique
- Le stockage de l'eau et de molécules tel que certains pigments, molécules de défense, stockage transitoire des glucides, protéines et lipides.
- Rôle dans la pression et la turgescence cellulaire permettant la rigidité de certaines structures telles que les fleurs, les tiges ou les feuilles.
Origine
Les ancêtres des différentes lignées d’eucaryotes photosynthétiques se nourrissaient probablement par phagocytose, processus cellulaire qui leur a permis, par le phénomène d'endosymbiose, d'internaliser une cellule photosynthétique et d'en faire un plaste. Ils n'avaient pas de paroi cellulaire qui aurait entravé la phagocytose. Une fois le plaste acquis, cet organite nourrit la cellule et rend la phagocytose inutile. La paroi peut alors être sélectionnée, car elle protège la cellule contre les stress biotiques et abiotiques. L'acquisition de cette paroi a pour conséquence que la cellule ne peut plus expulser ses déchets. D’où l’apparition d’une vacuole qui stocke ces déchets et en corollaire participe au port dressé des plantes terrestres en formant un squelette hydrostatique (analogue à l'hydrosquelette animal) grâce aux échanges ioniques et hydriques responsables de la turgescence. Ces phénomènes (endosymbiose, acquisition du plaste qui permet en cascade, celle de la paroi et de la vacuole) illustrent un mécanisme de l’évolution, la convergence[2].
Cellule végétale
Présentation

La vacuole centrale des cellules végétales et des champignons a une fonction homéostatique, assurant la turgescence cellulaire, et sa membrane, le tonoplaste, possède des pompes ioniques. Elle contient de l'eau, des glucides, des ions organiques et inorganiques et des pigments. Certaines vacuoles centrales sont également des sites d'accumulation de réserves ou de substances particulières, parfois toxiques (latex, opium...) voire de déchets, ce qui leur confère un rôle de détoxification de la cellule.
La vacuole représente entre 80 et 90 % du volume cellulaire de la cellule végétale adulte[3]. Son contenu est essentiellement constitué de solutions, parfois cristallisées. C'est ainsi que les réserves de saccharose de la betterave et la canne à sucre ou d'inuline chez les Astéracées (Composées) sont accumulées en solution dans les vacuoles de leurs cellules. Des réserves de malate peuvent également jouer un rôle important dans la fonction photosynthétique de certaines plantes grasses (photosynthèse de type CAM qui utilise le malate comme forme de stockage de CO2 en prévision des périodes les plus chaudes de la journée).
Ce sont les molécules du réticulum endoplasmique lisse qui engendrent les vacuoles. Les vacuoles peuvent aussi protéger la plante contre les prédateurs, car elles renferment parfois des composés toxiques ou désagréables au goût. La vacuole joue un rôle primordial dans la croissance de la cellule végétale.
La pression osmotique est directement proportionnelle à la quantité de molécules et d’ions dissous. Dans la vacuole, la pression osmotique est toujours supérieure à la pression atmosphérique (2 à 20 atm). La pression osmotique des vacuoles est globalement stable, sauf au niveau des stomates où elle est régulée pour ouvrir ou fermer l’ostiole. L’état de turgescence est l’état dans lequel la membrane est plaquée contre la paroi, grâce à la pression osmotique de la vacuole. Il est possible in vitro de sortir de l’état de turgescence en plaçant la cellule dans un milieu hypertonique. L’eau sort alors de la vacuole : c’est la plasmolyse. Ce phénomène a aussi lieu lorsque la plante souffre de sécheresse.
Lors de la croissance de la cellule, c’est principalement la vacuole qui grandit.
Acides carboxyliques
Les acides carboxyliques confèrent un pH faible aux vacuoles, en général proche de pH5. Les principaux sont l'acide citrique, l'acide oxalique et l'acide malique.
Les plantes grasses vivent en milieu très sec, et donc doivent maintenir leurs stomates fermés durant la journée. Elles ne peuvent donc pas importer de CO2 pour la photosynthèse. Elles stockent donc le CO2 la nuit dans la vacuole sous forme d’acide malique. Le jour il est restitué et utilisé pour la photosynthèse.
Glucides
- Saccharose : La concentration en saccharose de la vacuole suit un cycle de 24h. Elle est maximale vers midi, où la photosynthèse est importante. Ce cycle recommence donc tous les jours.
- Fructosane : Dans les organes de réserve (tubercule), la périodicité n’est plus d’une journée mais de plusieurs mois.
Les acides aminés et les protéines
Bien que riches en acides aminés, les vacuoles sont relativement pauvres en protéines. Elles ne contiennent que 5 % des protéines totales de la cellule (alors qu’elles représentent 80 % du volume de la cellule). Ce sont toutes des protéines hydrolytiques (protéases, estérase…) capables de dégrader des molécules.
Pigments flavonoïdes
La principale famille des pigments flavonoïdes est la famille des anthocyanes. Les pigments sont responsables de la couleur des fleurs : ce sont des molécules absorbant certaines longueurs d’onde lumineuse. En modifiant le pH, on peut modifier la délocalisation électronique de la molécule, et donc la couleur. Voilà pourquoi une fleur vieillissante change de couleur, en effet son pH augmente[réf. nécessaire].
Les pigments peuvent donc aussi être utilisés en tant qu’indicateur de pH. Les anthocyanes étant hydrophobes, ils sont stockés dans la vacuole sous forme d’anthocyanosides.
Alcaloïdes et tannins
Les tannins et les alcaloïdes sont présents dans les tissus de surface, et ont un rôle de défense. Grâce à leur effet dénaturant pour les protéines, les tannins rendent la plante peu digeste, en bloquant les enzymes digestives des herbivores. Les alcaloïdes sont des substances azotées. Ils sont toxiques (morphine, nicotine, pavot, opium…).
Les alcaloïdes ont aussi une propriété antimitotique, c'est-à-dire qu’ils bloquent la mitose, ce qui permet de réguler la croissance de la plante.
Vacuole des graines
Lors de la maturation de la graine, la teneur en eau baisse fortement, et donc la vacuole se fragmente. Lors de la germination, il y a une entrée d’eau, et les fragments de vacuole fusionnent. Dans les vacuoles fragmentées, on trouve une grande quantité de protéines de réserve et de protéase. Les protéases dégraderont les protéines de réserves en acides aminés lors de la germination. Ces acides aminés serviront ensuite à synthétiser de nouvelles protéines.
Les protéines de réserve sont peu polaires car elles sont stockées dans un milieu pauvre en eau. Il existe trois catégories de protéines de réserve : les prolamines, les globulines, et les lectines.
Les protéines de réserve et les protéases sont des protéines sécrétoires. Elles sont synthétisées dans l'ergatoplasme et rejoignent la vacuole par trafic vésiculaire (cf. bio cell animale). Elles possèdent une séquence d’adressage à la vacuole située sur le groupement amine terminal, le groupement carboxyle terminal, ou plus rarement à l’intérieur de la séquence des protéines.
Dans la vacuole fragmentée, il n'y a pas de présence simultanée de protéines et de protéases. Il ne faut pas que les protéines soient dégradées avant la germination. Il y a donc deux types de vacuoles, les vacuoles lytiques, qui stockent les protéases, et les vacuoles de réserve, qui stockent les protéines de réserve.
- Les vésicules SDV (lisses et denses) contiennent les protéines de réserve et sont adressées aux vacuoles de réserve.
- Les vésicules CCV (recouvertes de clathrine) contiennent les protéases, et sont donc adressées aux vacuoles lytiques.
Il existe des particules de reconnaissance entre les vésicules et les vacuoles : les particules vSNARE (vésicule), et tSNARE (target = cible).
Organismes unicellulaires
- Les vacuoles contractiles sont présentes chez certains Eucaryotes unicellulaires (anciennement appelés Protozoaires) vivant en eau douce. Ces vacuoles permettent d'évacuer l'eau en excès dans le cytoplasme mais également la locomotion par contraction antiphasique.
- La vacuole de phagocytose se rencontre chez les unicellulaires se nourrissant par invagination de la membrane plasmique autour d'une proie. Ce type de vacuole entre en coalescence avec des phytolysosome, et, après digestion s'ouvre à l'extérieur, rejetant les éléments non digérés.
Notes et références
- Venes, Donald (2001). Taber's Cyclopedic Medical Dictionary (Twentieth Edition), (F.A. Davis Company, Philadelphia), p. 2287.
- Marc-André Selosse, « Les végétaux existent-ils encore ? », Pour la science, no 77, , p. 11 (lire en ligne).
- Docteur Roger Prat, professeur en physiologie végétale
 Portail de la biologie cellulaire et moléculaire
Portail de la biologie cellulaire et moléculaire