Desmodus rotundus
Vampire commun, Vampire d'Azara


LC : Préoccupation mineure
Le Vampire commun (Desmodus rotundus), aussi appelé Vampire d'Azara, est une espèce de chauves-souris de la famille des Phyllostomidae. Elle est l'une des trois espèces actuelles de la sous-famille des Desmodontinae, nommées « chauves-souris vampires » du fait de leur hématophagie, les deux autres étant le Vampire à pattes velues (Diphylla ecaudata) et le Vampire à ailes blanches (Diaemus youngi). Ce groupe d'espèces constitue les seuls mammifères parasites connus. Le Vampire commun se nourrit principalement du sang du bétail. Pour une chauve-souris, il est particulièrement bien adapté à la locomotion au sol. Il approche ses proies de nuit, lorsqu'elles sont endormies, et utilise ses dents coupantes comme une lame de rasoir pour découper la peau de ses hôtes ; il lape ensuite le sang coulant de la plaie avec sa langue.
Cette espèce est fortement polygyne, et des mâles dominants défendent un groupe de femelles. C'est l'une des espèces de chauves-souris les plus sociales ; elle présente plusieurs comportements coopératifs, comme le toilettage mutuel ou le partage de la nourriture. Le Vampire commun vit au Mexique, en Amérique centrale et en Amérique du Sud. Il est considéré comme un nuisible par l'Homme, puisqu'il se nourrit sur le bétail et qu'il peut être porteur du virus de la rage. L'espèce dispose d'une grande aire de répartition, elle est abondante, ses effectifs stables et susceptibles de s'adapter aux modifications de son habitat ; pour ces raisons, l'Union internationale pour la conservation de la nature place ce mammifère dans la catégorie « préoccupation mineure ».
Caractères distinctifs

Le Vampire commun mesure environ 9 cm de long, pour une envergure de 18 cm. Il pèse entre 25 et 40 grammes, mais son poids peut considérablement augmenter une fois qu'il s'est alimenté[1]. En captivité, il est capable de consommer en moyenne 59,5 % de son poids corporel en sang en 30 minutes[2]. Cette chauve-souris a le pelage ras et bicolore, le gris argenté sur la face ventrale tranchant avec la fourrure foncée du dos[3]. Ses ailes sont noirâtres avec un dessous gris ou blanc et brun, et elle n'a pas de queue. Les oreilles sont triangulaires. Son nez est une feuille nasale rudimentaire, plissée, formant des bourrelets au-dessus des narines, et la lèvre inférieure est profondément échancrée. La boîte crânienne est relativement grande, mais le museau est réduit pour faire place aux grandes incisives et canines supérieures triangulaires. C'est l'espèce de chauves-souris qui a le moins de dents, ayant pour formule dentaire 1.1.1.12.1.2.1 (dans l'ordre incisives, canines, prémolaires et molaires). Les incisives supérieures n'ont pas d'émail dentaire, pour rester aiguisées comme des rasoirs[3].

Alors que la plupart des autres chauves-souris ont presque complètement perdu la capacité de manœuvrer à terre, les chauves-souris vampires sont une des exceptions parmi les chiroptères[4]. Les espèces de ce groupe peuvent courir en utilisant une démarche unique, dans laquelle les membres antérieurs sont utilisés à la place des membres postérieurs pour se propulser vers l'avant, les ailes étant beaucoup plus puissantes que les pieds postérieurs. Cette capacité a probablement évolué plusieurs fois de façon indépendante dans la lignée de la chauve-souris[4]. Le Vampire commun est muni de trois coussinets sous le pouce, qui lui sert de sole[3]. Il est également capable de sauter dans diverses directions, à différentes distances et à différents rythmes[5]. Lorsqu'il saute, les pattes arrière du vampire lui donnent une première impulsion, mais l'essentiel de la poussée est réalisée par les membres pectoraux. L'arrière du corps décolle, le centre de gravité est ramené vers l'avant, et les membres postérieurs servent alors à maintenir à l'équilibre alors que l'animal repose sur ses pouces et pousse sur ses bras[5].
Les sens principaux (vue, odorat et ouïe) sont bien développés chez le Vampire commun : la vue est comparable à celle d'un rat. Ces chauves-souris utilisent probablement ce sens pour s'orienter à longue distance ; les structures nasales sont grandes et le bulbe olfactif bien développé ; la cochlée (oreille interne) est très sensible aux basses fréquences[3]. Le Vampire commun utilise l'écholocation, volant la bouche ouverte pour émettre ses ultrasons situés entre 45 et 100 kHz avec deux ou trois harmoniques[3]. Cette chauve-souris peut identifier une bande métallique d'un centimètre de large à 50 cm de distance, ce qui est d'une précision modérée par rapport aux autres chauves-souris[6].
Écologie et comportement
Alimentation

Le Vampire commun est hématophage et boit principalement le sang de mammifères, et notamment du bétail comme les bovins ou les équidés[7], mais aussi les chèvres, moutons ou cochons[8]. Il se nourrit aussi sur des animaux sauvages comme les tapirs, mais préfère les animaux domestiques, et privilégie les chevaux sur les bovins quand il a le choix[9]. Les femelles, notamment celles en œstrus, sont également préférées aux mâles, peut-être à cause de leurs hormones[10]. Le Vampire commun peut très occasionnellement se nourrir sur les humains[8],[11]. Dans le désert d'Atacama et sur les îles très arides au large du Pérou, où il n'existe aucun mammifère terrestre, le Vampire commun peut se nourrir sur les jeunes Manchots de Humboldt (Spheniscus humboldti)[12],[13] et sur les Otaries à crinière (Otaria flavescens)[14].

Le Vampire commun s'alimente la nuit[7], en utilisant l'écholocation et l'olfaction pour repérer ses proies[15]. Il chasse sur une distance de 5 à 8 km autour de son gîte[16]. Une fois la cible repérée, le vampire atterrit sur elle, ou saute sur elle depuis le sol[7],[16]. Les zones de morsures privilégiées sur le bétail sont les épaules, le cou, la base des cornes, la base des oreilles, le museau, les pattes, la queue, la vulve et l'anus[8]. Le Vampire commun dispose de senseurs de chaleur dans son nez qui l'aident à détecter les vaisseaux sanguins passant près de la surface de la peau[9]. Il perce la peau de l'animal à l'aide de ses incisives, soulevant un petit rabat de peau[16], et lape le sang avec sa langue, qui a des rainures latérales adaptées à ce mode d'alimentation[17]. Si le sang ne s'écoule pas suffisamment, le Vampire commun peut mordre à nouveau plus en profondeur ou s'enfoncer plus profondément dans la blessure avec sa langue. La blessure est indolore, et la salive de cette chauve-souris possède une enzyme qui empêche le sang de coaguler et qui fait que les plaies continuent de saigner pendant plusieurs heures[8],[18].
Le nourrissage durerait entre 9 et 40 minutes, le temps pour la chauve-souris de prélever une vingtaine de millilitres de sang. Le Vampire commun protège son hôte et ne laisse pas ses congénères partager sa proie quand il se nourrit. Il est ainsi rare de voir deux vampires ou davantage se nourrir sur un même hôte, à l'exception des mères et de leur progéniture[15],[19], mais jusqu'à huit individus ont déjà été observés se nourrissant sur le même hôte, et jusqu'à sept se succédant sur la même blessure sur une période de trois heures[8].
Reproduction

Au sein d'un gîte, les mâles entrent en compétition pour les femelles, et chacun défend une partie du gîte en poussant, chassant, mordant les autres mâles s'approchant[17]. Ils défendent les sites attirant les femelles, mais celles-ci changent souvent de colonies. Certains mâles sont plus dominants que d'autres, s'accouplant avec davantage de femelles et engendrant plus de descendance[20]. Pour copuler, le mâle monte sur le dos de la femelle, maintient les ailes de celle-ci repliées à l'aide de ses propres ailes, et lui tient la nuque dans sa bouche ; l'acte dure trois à quatre minutes[17]. En œstrus, la femelle ne produit qu'un ovule[21], et n'a donc en général qu'un petit par portée[7],[16], qui vient au monde après une gestation d'environ sept mois[21]. Le Vampire commun peut se reproduire toute l'année, bien que le nombre de conceptions et de naissances connaisse un pic pendant la saison des pluies[7],[16].

Le jeune est principalement élevé par la femelle. Les mères laissent leur petit pour aller chasser, et l'appellent à leur retour au gîte pour l'allaiter[7]. À l'âge de six mois, le jeune accompagne sa mère quand elle part en chasse, mais n'est pas totalement sevré avant neuf mois[7]. Les jeunes femelles restent habituellement dans leur groupe natal jusqu'à l'âge adulte, à moins que leur mère ne meure ou ne se déplace[20]. Les mouvements occasionnels de femelles non-apparentées entre les groupes conduit à la formation de plusieurs lignées maternelles au sein des groupes[20]. Les jeunes mâles ont tendance à vivre dans leur groupe natal jusqu'à ce qu'ils soient âgés d'un ou deux ans. Ils sont parfois chassés du groupe par les mâles adultes résidents[20].
Comportements sociaux

Le partage de nourriture par régurgitation chez le Vampire commun a été étudié à la fois en laboratoire et sur le terrain, et s'explique par le niveau de parenté des individus concernés, l'association et la réciprocité de l'aide[22]. Dans une étude de terrain menée au Costa Rica de 1978 à 1983[9], des Vampires communs changeaient fréquemment de gîtes, se retrouvant à dormir dans des arbres avec des individus apparentés et non-apparentés[22]. Le degré de parenté génétique au sein des groupes était faible (r = 0,03 à 0,11), mais 95 % des partages de nourriture observés dans ce contexte impliquaient des parents proches (cousins germains ou plus proches). La plupart des partages de nourriture observés (70 %) concernaient des mères qui alimentaient leur jeune, mais les événements de partage non-maternels étaient biaisés vers la parentèle, suggérant que les chauves-souris vampires préfèrent aider leurs proches. Toutefois, la fréquence d'interaction entre les individus, apparentés ou non, s'est également avéré jouer un rôle dans la mise en place de ces interactions[22], et la réciprocité de l'aide semble jouer un rôle plus important que la parenté ou l'insistance des individus receveurs[23].
Le Vampire commun pratique également le toilettage mutuel[15] : deux chauves-souris se lèchent simultanément l'une l'autre, et renforcent leurs liens sociaux[24]. Les individus qui se toilettent mutuellement partagent aussi leur nourriture. Il a été suggéré que les vampires peuvent déterminer si un autre individu a réellement besoin de manger en léchant son abdomen[24].
En 2019, une étude montre que les liens sociaux entre les chauves-souris vampires captifs persistent dans la nature Les chauves-souris qui partagent de la nourriture avec leurs compagnons de cage restent proches après leur libération. La persistance de nombreuses relations dans différents environnements physiques et sociaux suggère que la structure sociale est causée à la fois par des contraintes extrinsèques et par la fidélité intrinsèque du partenaire[25].
Prédateurs, parasites et maladies
_feeding_on_Common_Vampire_Bat_(Desmodus_rotundus)_(cropped).jpg.webp)
Les prédateurs non-humains du Vampire incluent le chat domestique[11], des rapaces nocturnes tels la Chouette effraie (Tyto alba) ou la Chevêche des terriers (Athene cunicularia) et des serpents comme Pseudelaphe flavirufa, le Boa constricteur (Boa constrictor), le Fer de lance commun (Bothrops atrox)[21] ou le Boa arc-en-ciel (Epicrates cenchria)[26]. La chauve-souris carnivore Chrotopterus auritus a également été observée mangeant un Vampire commun, et au Mexique le Quiscale à longue queue (Quiscalus mexicanus) et le Geai enfumé (Psilorhinus morio) ont été signalés comme ayant attrapé et mangé des Vampires communs capturés au filet japonais quand ceux-ci ont été relâchés au matin[21].
Parmi les ectoparasites, on compte de nombreux acariens (notamment des genres Radfordiella, Periglischrus, Ornithonyssus, Chirnyssoides, Ornithodoros, Ixodes, Amblyomma, Euschoengastia, Trombicula et Nycterinastes), des puces (Polygenis) et des mouches piqueuses (Basilia, Trichobius, Megistopoda, Speiseria, Trichobioides et Strebla)[21]. Pour les parasites internes, on recense des nématodes (comme Biacantha desmoda, B. normaliae[21],[27] et des œufs du genre Capillaria[28]), la bactérie Grahamella brumpti ainsi que plusieurs trypanosomes (T. cruzi, T. vespertilionis, T. pessoai et T. megadermae)[21]. Le Vampire commun peut également contaminer le bétail avec certaines maladies, comme les trypanosomiases à T. hippicum ou à T. equinum, ou même parfois transmettre le virus de la rage. Il n'en est cependant pas un porteur à long terme puisqu'il est également affecté par ce virus et meurt s'il contracte la rage[29].
Répartition et habitat
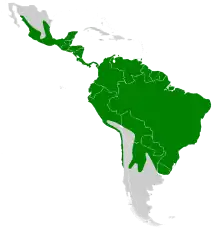
Cette espèce se rencontre au Mexique, en Amérique centrale et en Amérique du Sud, en plaine et jusqu'à environ 1 500 m d'altitude. Au Nord, elle peut être trouvée jusqu'à 280 km au sud de la frontière entre les États-Unis et le Mexique, et des données fossiles sont connues de Floride et des États américains limitrophes du Mexique. Le Vampire commun est l'espèce de chauves-souris la plus commune dans le Sud-Est du Brésil[30]. Son aire de répartition s'étend au sud jusqu'en Uruguay, dans le nord de l'Argentine et le centre du Chili. Dans les Antilles, l'espèce est uniquement présente sur Trinité. Le Vampire commun préfère les climats chauds et humides[7],[19], et trouve sa nourriture dans les forêts tropicales et subtropicales et les prairies ouvertes. Les colonies s'abritent dans les arbres, les grottes, les bâtiments abandonnés, les vieux puits et les mines[7],[31]. Le Vampire commun peut partager ses gîtes avec près de 45 autres espèces de chauves-souris[21], mais il semble être le plus dominant sur ces sites de repos. Le Vampire commun occupe les endroits les plus sombres et les plus élevés, et les autres espèces de chauves-souris se déplacent pour prendre ces emplacements vacants quand le Vampire sort[31].
Taxinomie

Le Vampire commun est décrit par Étienne Geoffroy Saint-Hilaire en 1810 sous le protonyme de Phyllostoma rotundum, qui lui donne le nom de « Phyllostome à feuille arrondie »[32]. Différents noms scientifiques lui sont attribués avant qu'Oldfield Thomas ne lui donne le nom de Desmodus rotundus en 1901. Elle est l'unique représentant actuel du genre Desmodus, mais d'autres espèces, fossiles, ont été décrites. Plusieurs taxons distincts ont été décrits avant d'être placés en synonymie avec Desmodus rotundus[3] :
- Rhinolophus ecaudatus Schinz, 1821
- Desmodus rufus Wied, 1826
- Edostoma cinerea D'Orbigny, 1834
- Desmodus d'orbigny Waterhouse, 1839
- Desmodus murinus Wagner, 1840
- Desmodus fuscus Burmeister, 1854
- Desmodus mordax Burmeister, 1879
Le Vampire commun appartient à la sous-famille des Desmodontinae, aux côtés de deux autres espèces actuelles : le Vampire à pattes velues (Diphylla ecaudata) et le Vampire à ailes blanches (Diaemus youngi). Les chauves-souris de cette sous-famille constituent les « vampires » vrais, par opposition aux « faux-vampires » de la famille des Megadermatidae et de la Chauve-souris javelot (Vampyrum spectrum), aussi appelée « faux-vampire de Linné ». Les trois espèces de Desmodontinae sont des hématophages spécialisés, se nourrissant du sang des animaux à sang chaud[33]. Le Vampire commun se nourrit cependant plus sur les mammifères que les deux autres espèces, qui exploitent principalement les oiseaux[34],[35]. Ces trois chauves-souris se ressemblent, mais le Vampire commun peut se distinguer, entre autres, par son pouce plus long[34].
Deux sous-espèces sont parfois distinguées[3] :
Le Vampire commun et l'être humain
Transmission du virus de la rage
Un bactériologiste de Trinidad, Joseph Lennox Pawan (en), trouve la première chauve-souris vampire infectée par le virus de la rage en mars 1932 et montre rapidement que diverses espèces de chauves-souris, dont le Vampire commun, sont capables de transmettre la maladie à l'humain. Cette découverte marque une étape importante vers le développement d'un vaccin et vaut à Pawan la décoration de l'Ordre de l'Empire britannique[36]. En 1980, la Smithsonian Institution écrivait que les plus grandes prévalences de rage chez les chauves-souris vampires se trouvent dans les grandes populations d'Amérique du Sud, et que sous ces latitudes le danger de transmission ne concerne pas tant la population humaine, mais plutôt le bétail[37]. Au début du XXIe siècle cependant, la transmission du virus de la rage à l'humain a considérablement augmenté. À titre d'exemple, entre 2000 et 2007, sur 380 cas de rage recensés par un institut sanitaire péruvien, 293 (soit 77 %) concernent des variants rabiques provenant de chauves-souris vampires, les 87 autres (soit 23 %) concernant des chiens[38]. Des épidémies de rage existent aussi chez les communautés vivant dans la forêt amazonienne, mais sont peu étudiées et documentées en raison de contraintes techniques et culturelles[38]. Bien qu'il ait été jugé possible que le virus puisse être transmis sans morsure ni contact direct[39],[40], c'est normalement le mode de transmission de la maladie. Ainsi, la manipulation de ces bêtes doit se faire avec les précautions prophylactiques nécessaires, comme pour tout autre animal sauvage[41].
Dans la recherche pharmaceutique
La salive de la chauve-souris vampire contient des composés fluidifiant très rapidement le sang permettant à cet hématophage de s'alimenter. Une enzyme se nommant Desmodus Salivary Plasminogen Activator (DSPA), parfois surnommée « desmotéplase » ou « draculine », présente des caractéristiques prometteuses dans la prise en charge des accidents vasculaires cérébraux et est plus efficace que ses analogues pharmaceutiques, les agents thrombolytiques mimant l'action de la t-PA et de l'u-PA (action thrombolytique 180 fois plus importante que ces derniers)[42],[43],[44].
Le Vampire
Cette chauve-souris fait l'objet d'un court-métrage de Jean Painlevé, paru en 1945 et intitulé Le Vampire. Ce réalisateur, spécialisé dans le tournage en milieu sous-marin, signe là son seul film concernant un mammifère, terrestre de surcroît[45]. Ce documentaire de neuf minutes, en noir et blanc, présente un Vampire commun détenu en captivité et mal nourri se sustenter sur un Cobaye (Cavia porcellus)[45]. La scène est introduite par des images de Nosferatu le vampire de Friedrich Wilhelm Murnau, et entrecoupées d'images empruntées à un autre documentaire de Painlevé sur Trypanosoma gambiense, parasite sanguin africain proche cousin d'espèces transmises par le Vampire commun, pour dramatiser les effets de la morsure du vampire[46]. Vers la fin du documentaire, Painlevé montre ce qu'il nomme le « salut du Vampire », la chauve-souris étendant une de ses ailes avant d'aller dormir, qu'il rapproche du « salut hitlérien »[46],[47],[48]. La chauve-souris en question, gardée à l'Institut Pasteur, a été gazée par ses gardiens lorsque la guerre éclate, qui craignaient qu'elle ne s'échappe durant des bombardements et ne répande des maladies[46].
Menaces et protection
Le Vampire commun a probablement connu une expansion démographique massive avec l'arrivée des Européens et l'élevage de bétail en Amérique latine[49]. L'espèce dispose d'une grande aire de répartition, n'est pas exploitée commercialement, et même si elle est victime de sa réputation de porteuse de la rage, l'Union internationale pour la conservation de la nature juge les effectifs stables et ne considère pas ce phénomène comme une menace majeure, traitant par conséquent le Vampire commun comme espèce de « préoccupation mineure »[50]. Les pertes que le Vampire commun inflige aux animaux d'élevage, notamment à cause de la rage, fait que l'espèce est la cible de mesures d'éradication, allant de l'obturation de l'entrée des grottes à la destruction de la végétation indigène ou l'utilisation de poison tel le coumaphène, pratiques qui nuisent à de nombreux autres chiroptères, parfois gravement menacés[51].
Annexes
Bibliographie
- (en) Arthur M. Greenhall, Gerhard Joermann, Uwe Schmidt et Michael R. Seidel, « Desmodus rotundus », Mammalian Species, vol. 202, , p. 1-6 (DOI 10.2307/3503895, lire en ligne).
Références taxinomiques
- (en) Référence Mammal Species of the World (3e éd., 2005) : Desmodus rotundus E. Geoffroy, 1810 (consulté le ).
- (en) Référence Catalogue of Life : Desmodus rotundus (E. Geoffroy, 1810) (consulté le ).
- (fr+en) Référence ITIS : Desmodus rotundus (E. Geoffroy, 1810) (consulté le ).
- (en) Référence Paleobiology Database : Desmodus rotundus Geoffroy 1810 (consulté le ).
Liens externes
- (en) Référence Animal Diversity Web : Desmodus rotundus (consulté le ).
- (en) Référence North American Mammals : Desmodus rotundus (consulté le ).
- (en) Référence NCBI : Desmodus rotundus (taxons inclus) (consulté le ).
- (en) Référence uBio : Desmodus rotundus Thomas 1901 (consulté le ).
- (en) Référence UICN : espèce Desmodus rotundus (É. Geoffroy, 1810) (consulté le ).
Notes et références
- (en) Cet article est partiellement ou en totalité issu de l’article de Wikipédia en anglais intitulé « Common vampire bat » (voir la liste des auteurs).
- (en) Gerald Carter, « Foraging vampire bats can expect big meals or none at all », sur social bat.org, (consulté le )
- (en) Cristina Busch, « Consumption of blood, renal function and utilization of free water by the vampire bat, Desmodus rotundus », Comparative biochemistry and physiology. A, Comparative physiology, Elsevier, vol. 90, no 1, , p. 141-146 (ISSN 0300-9629, OCLC 01237399, PMID 2900098, DOI 10.1016/0300-9629(88)91019-5)
- Greenhall et al. (1983), p. 1
- (en) Daniel K. Riskin et John W. Hermanson, « Biomechanics: Independent evolution of running in vampire bats », Nature, vol. 434, no 7031, , p. 292–292 (DOI 10.1038/434292a)
- (en) Joe Scott Altenbach, Locomotor morphology of the vampire bat, Desmodus rotundus - Special publication no 6, American Society of Mammalogists, , 137 p. (lire en ligne)
- (en) Uwe Schmidt et Christel Schmidt, « Echolocation Performance of the Vampire Bat (Desmodus rotundus) », Zeitschrift für Tierpsychologie, vol. 45, no 4, , p. 349–358 (DOI 10.1111/j.1439-0310.1977.tb02025.x)
- (en) R.D. Lord, « A Taste for Blood: The Highly Specialized Vampire Bat Will Dine on Nothing Else », Wildlife Conservation, vol. 96, , p. 32-38
- Greenhall et al. (1983), p. 4
- (en) Gerald S. Wilkinson, « Food Sharing in Vampire Bats », Scientific American, vol. 262, no 21, , p. 76-82 (lire en ligne)
- (en) W. A. Schutt, F. Muradali, N. Mondol, K. Joseph et K. Brockmann, « Behavior and Maintenance of Captive White-Winged Vampire Bats, Diaemus youngi », Journal of Mammalogy, vol. 80, no 1, , p. 71–81 (DOI 10.2307/1383209)
- (en) H. Delpietro, F. Konolsaisen, N. Marchevsky et G. Russo, « Domestic cat predation on vampire bats (Desmodus rotundus) while foraging on goats, pigs, cows and human beings », Applied Animal Behaviour Science, vol. 39, no 2, , p. 141–150 (DOI 10.1016/0168-1591(94)90134-1)
- (en) Guillermo Luna-Jorquera et Boris M. Culik, « Penguins bled by vampires », Journal of Ornithology, vol. 136, no 4, , p. 471–472 (DOI 10.1007/BF01651597)
- (en) Ella Davies, « Vampire bats attack penguin chicks », BBC Nature (consulté le )
- (en) Alessandro Catenazzi et Maureen A. Donnelly, « Sea lion Otaria flavescens as host of the common vampire bat Desmodus rotundus », Marine Ecology Progress Series, vol. 360, , p. 285–289 (DOI 10.3354/meps07393, lire en ligne)
- (en) J. Wilkinson, « Bat Blood Donors », dans D. MacDonald et S. Norris, The Encyclopedia of Mammals. Facts on File, (ISBN 0-87196-871-1), p. 766-767
- (en) R.M. Nowak, Walker's Mammals of the World, Baltimore/London, Johns Hopkins University Press, , 1629 p. (ISBN 0-8018-3970-X), p. 1629
- (en) Michael Mulheisen et Rebecca Anderson, « Desmodus rotundus », Animal Diversity Web (consulté le )
- (en) T. Cartwright, « The Plasminogen Activator of Vampire Bat Saliva », Blood, vol. 43, no 3, , p. 317-326 (lire en ligne)
- (en) Gerald S. Wilkinson, « The Social Organization of the Common Vampire Bat 1: Pattern and Cause of Association », Behavioral Ecology and Sociobiology, vol. 17, no 2, , p. 111–21 (DOI 10.1007/BF00299243, lire en ligne)
- (en) Gerald S. Wilkinson, « The Social Organization of the Common Vampire Bat II: Mating system, genetic structure, and relatedness », Behav. Ecol. Sociobiol, vol. 17, no 2, , p. 123–34 (lire en ligne)
- Greenhall et al. (1983), p. 3
- (en) Gerald Carter et Gerald Wilkinson, « Does food sharing in vampire bats demonstrate reciprocity? », Communicative & Integrative Biology, vol. 6, no 6, , e25783 (DOI 10.4161/cib.25783)
- (en) Gerald G. Carter et Gerald S. Wilkinson, « Food sharing in vampire bats: reciprocal help predicts donations more than relatedness or harassment », Proceedings of the Royal Society B: Biological Sciences, vol. 280, no 1753, , p. 20122573–20122573 (DOI 10.1098/rspb.2012.2573, lire en ligne)
- (en) Gerald S. Wilkinson, « Social grooming in the common vampire bat, Desmodus rotundus », Animal Behaviour, vol. 34, no 6, , p. 1880–1889 (DOI 10.1016/S0003-3472(86)80274-3, lire en ligne)
- (en) Simon P Ripperger, Gerald G Carter, Niklas Duda, Alexander Koelpin, Björn Cassens, Rüdiger Kapitza, Darija Josic, Jineth Berrío-Martínez, Rachel A Page et Frieder Mayer, « Vampire bats that cooperate in the lab maintain their social networks in the wild », Current Biology, Royaume-Uni, Cell Press et Elsevier, vol. 29, no 23, et , p. 4139-4144.E4 (ISSN 0960-9822 et 1879-0445, OCLC 45113007, PMID 31679938, DOI 10.1016/J.CUB.2019.10.024)
- (en) Sarah Martin-Solano, Theofilos Toulkeridis, Aaron Addison et Wilmer E. Pozo-Rivera, « Predation of Desmodus rotundus Geoffroy, 1810 (Phyllostomidae, Chiroptera) by Epicrates cenchria (Linnaeus, 1758) (Boidae, Reptilia) in an Ecuadorian Cave », Subterranean Biology, Pensoft Publishers (d), vol. 19, , p. 41-50 (ISSN 1768-1448 et 1314-2615, OCLC 57902976, DOI 10.3897/SUBTBIOL.19.8731)
- (en) Mirna C. Oviedo, Geraldine Ramallo, Lucía E. Claps et M. Daniela Miotti, « A new species of Biacantha (Nematoda: Molineidae), a parasite of the common vampire bat from the Yungas, Argentina. », Journal of Parasitology, vol. 98, no 6, , p. 1209–1215 (DOI 10.1645/GE-3155.1)
- (en) Andrés Rojas et Ricardo Guerrero, « Capillaria sp., isolated from Desmodus rotundus (Chiroptera: Phyllostomidae) in Costa Rica », Mastozoología Neotropical, vol. 14, no 1, , p. 101-102 (lire en ligne)
- Greenhall et al. (1983), p. 3-4
- (en) Eleonora Trajano, « Movements of Cave Bats in Southeastern Brazil, with Emphasis on the Population Ecology of the Common Vampire Bat, Desmodus rotundus (Chiroptera) », Biotropica, vol. 28, no 1, , p. 121 (DOI 10.2307/2388777)
- (en) Timothy J. Wohlgenant, « Roost Interactions Between the Common Vampire Bat (Desmodus rotundus) and Two Frugivorous Bats (Phyllostomus discolor and Sturnira lilium) in Guanacaste, Costa Rica », Biotropica, vol. 26, no 3, , p. 344 (DOI 10.2307/2388857)
- (fr) Étienne Geoffroy Saint-Hilaire, « Description des roussettes et des céphalotes, deux nouveaux genres de la famille des chauve-souris », Annales du Muséum National d'Histoire Naturelle, vol. 15, , p. 86-108 (lire en ligne)
- (en) John Frederick Eisenberg et Redford Kent Hubbard, Mammals of the Neotropics, Volume 3, University of Chicago Press, , 187–88 p. (ISBN 0-226-19542-2)
- (en) Arthur M. Greenhall et William A. Schutt, « Diaemus youngi », Mammalian Species, no 533, , p. 1 (DOI 10.2307/3504240)
- (en) Arthur M. Greenhall, Uwe Schmidt et Gerhard Joermann, « Diphylla ecaudata », Mammalian Species, no 227, , p. 1 (DOI 10.2307/3504022)
- (en) « Joseph Lennox Pawan », Caribbean Council for Science and Technology (consulté le )
- (en) Department of Systematic Biology, Vertebrate Zoology, « Bat Facts », Smithsonian Institution, (consulté le )
- (en) Rene Edgar Condori-Condori, Daniel G. Streicker, Cesar Cabezas-Sanchez et Andres Velasco-Villa, « Enzootic and Epizootic Rabies Associated with Vampire Bats, Peru », Emerging Infectious Diseases, vol. 19, no 9, (ISSN 1080-6040, DOI 10.3201/eid1909.130083)
- (en) Denny G. Constantine, « Rabies Transmission by Nonbite Route », Public Health Reports (1896-1970), vol. 77, no 4, , p. 287 (DOI 10.2307/4591470)
- (en) Sharon L. Messenger, Jean S. Smith et Charles E. Rupprecht, « Emerging Epidemiology of Bat‐Associated Cryptic Cases of Rabies in Humans in the United States », Clinical Infectious Diseases, vol. 35, no 6, , p. 738–747 (ISSN 1058-4838, DOI 10.1086/342387)
- (en) CDC, « Learning about bats and rabies », (consulté le )
- (en) Ana Z. Fernandez, Alfonso Tablante, Fulvia Bartoli, Suzette Beguin, H.C. Hemker et Rafael Apitz-Castro, « Expression of biological activity of draculin, the anticoagulant factor from vampire bat saliva, is strictly dependent on the appropriate glycosylation of the native molecule », Biochimica et Biophysica Acta - General Subjects, vol. 1425, no 2, , p. 291–299 (DOI 10.1016/S0304-4165(98)00082-8)
- (en) G.T. Liberatore, A. Samson, C. Bladin, W.-D. Schleuning et R.L. Medcalf, « Vampire Bat Salivary Plasminogen Activator (Desmoteplase): A Unique Fibrinolytic Enzyme That Does Not Promote Neurodegeneration », Stroke, vol. 34, no 2, , p. 537–543 (ISSN 0039-2499, DOI 10.1161/01.STR.0000049764.49162.76, lire en ligne)
- (en) Robert L. Medcalf, « Desmoteplase: discovery, insights and opportunities for ischaemic stroke », British Journal of Pharmacology, Wiley-Blackwell, vol. 165, no 1, , p. 75-89 (ISSN 0007-1188 et 1476-5381, OCLC 01240522, PMID 21627637, PMCID 3252968, DOI 10.1111/J.1476-5381.2011.01514.X)
- (fr) Roxane Hamery, Jean Painlevé, le cinéma au cœur de la vie, Presses universitaires de Rennes, , 312 p. (lire en ligne), p. 13
- (en) Peter J. Bloom, French Colonial Documentary : Mythologies of Humanitarianism, University of Minnesota Press, , 265 p. (lire en ligne), p. 100-102
- (en) Jonathan Kahana, The Documentary Film Reader : History, Theory, Criticism, Oxford University Press, , 1056 p. (lire en ligne), p. 976
- (fr) Marc Guiga, « Fiche film - Vampire (Le) », sur Images de la culture (consulté le )
- (en) A. D. Ditchfield, « The comparative phylogeography of Neotropical mammals: patterns of intraspecific mitochondrial DNA variation among bats contrasted to nonvolant small mammals », Molecular Ecology, vol. 9, no 9, , p. 1307–1318 (ISSN 0962-1083, DOI 10.1046/j.1365-294x.2000.01013.x)
- Union internationale pour la conservation de la nature
- (en) Ludmilla M. S. Aguiar, Daniel Brito et Ricardo B. Machado, « Do Current Vampire Bat (Desmodus rotundus) Population Control Practices Pose a Threat to Dekeyser's Nectar Bat's (Lonchophylla dekeyseri) Long-Term Persistence in the Cerrado? », Acta Chiropterologica, vol. 12, no 2, , p. 275–282 (ISSN 1508-1109, DOI 10.3161/150811010X537855)
 Portail des mammifères
Portail des mammifères .svg.png.webp) Portail de l’Amérique
Portail de l’Amérique