تنظيم التعبير الجيني
تنظيم التعبير الجيني أو التنظيم الجيني (بالإنجليزية: Regulation of gene expression) هو مجموعة متنوعة من الآليات المستخدمة من قبل الخلايا لزيادة أو إنقاص نواتج جينية محددة (بروتين أو رنا). توجد أنظمة تعبير جيني متطورة في علم الأحياء ذات أهداف متعددة كتحفيز مسارات النماء، الاستجابة لمنبهات بيئية أو التأقلم مع مصادر غذاء جديدة. عمليا يمكن التحكم في أي مرحلة من التعبير الجيني، ابتداء من بدء الترجمة مرورا بمعالجة الرنا وانتهاء بتعديل ما بعد الترجمة. غالبا ما يتحكم منظم جين واحدٍ في منظم آخر -وهكذا- مشكلين شبكة تنظيم جينية.
التنظيم الجيني أساسي للفيروسات، بدائيات النوى وحقيقيات النوى لأنه يزيد من تعدد وظائف وقابلية تأقلم الكائن عبر السماح للخلية بالتعبير عن البروتين عند الحاجة وتثبيط إنتاجه عند زوالها. رغم أن باربرا مكلنتوك أظهرت سنة 1951 تآثرا بين موقعين جينيين منشط (Ac) وينقول (Ds) لتكوين لون بذور الذرة، يُعتبر أول اكتشاف لنظام تنظيم جيني هو اكتشاف مشغل lac بواسطة فرنسوا جاكوب وجاك مونو سنة 1961، وفيه لا يُعبَّر عن بعض الإنزيمات المسؤولة عن أيض اللاكتوز في الإشريكية القولونية سوى عند تواجد اللاكتوز وغياب الغلوكوز.[1]
في الكائنات متعددة الخلايا، يقود التنظيم الجيني عمليتي التمايز الخلوي[2] والتخلق الحيوي لدى الجنين وينتج عن ذلك تكون أنواع خلايا مختلفة تملك بروفايلات تعبير جيني مختلفة من نفس التسلسل الجينومي. رغم أن هذا لا يفسر منشأ وأصل التنظيم الجيني إلا أن علماء الأحياء التطوري يضيفونه كتفسير جزئي لطريقة عمل التطور على المستوى الجزيئي، وهو أمر مركزي لعلم الأحياء النمائي التطوري
حادث البدء الذي يقود إلى تغيُّر في التعبير الجيني يشمل تنشيط أو إيقاف نشاط المستقبلات.
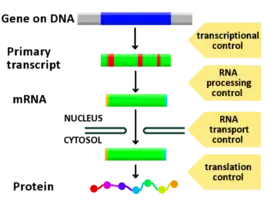
المراحل التي يتم فيها التنظيم الجيني
يمكن التحكم في أي مرحلة من التعبير الجيني، من نسخ الدنا إلى رنا إلى تعديل ما بعد الترجمة للبروتين. هذه قائمة بالمراحل التي يتم فيها تنظيم التعبير الجيني، والمرحلة التي يتم فيها التنظيم بشكل كبير جدا هي بدء الترجمة:
- نطاقات الكروماتين
- النسخ
- تعديل ما بعد النسخ
- نقل الرنا
- الترجمة
- تفكيك الرنا الرسول
تنظيم النسخ
يتحكم تنظيم النسخ بتوقيت حدوث النسخ وكمية الرنا المتشكلة. يمكن تنظيم نسخ الجين من خلال إنزيم بوليميراز الرنا عبر آليات متعددة. تغير عوامل التحديد نوعية بوليميراز الرنا لمحفز ما أو لمجموعة من المحفزات لتجعله أكثر أو أقل قدرة على الارتباط بها (العوامل سيغما المستخدمة في النسخ في طليعيات النوى). ترتبط الكابحات بالمُشغّل، وتشفّر تتاليات سلسة الدنا لتصبح أقرب إلى منطقة المحفز أو متداخلة معها، فتعيق تقدم بوليميراز الرنا وبالتالي التعبير عن الجين. تضع عوامل النسخ العامة بوليميراز الرنا في بداية التسلسل المشفر للبروتين ثم تطلق البوليميراز لنسخ الرنا الرسول. تعزز المنشطات التفاعل بين بوليميراز الرنا ومحفز معين، وتدعم التعبير عن الجين. تفعل المنشطات ذلك من خلال زيادة جذب بوليميراز الرنا للمحفز، من خلال التفاعلات مع الوحدات الفرعية لبوليميراز الرنا أو بشكل غير مباشر عن طريق تغيير بنية الدنا. المعززات مواقع على حلزون الدنا ترتبط بالمنشطات من أجل إغلاق حلقة الدنا وجلب محفز معين إلى مُعقّد البدء. المعززات أكثر شيوعًا في حقيقيات النوى مقارنة بطليعيات النوى، إذ توجد أمثلة قليلة في طليعيات النوى. الكاتمات هي مناطق من تسلسل الدنا، يمكنها عندما ترتبط بعوامل نسخ معينة أن تُسكت التعبير عن الجين.[3]
التنظيم من قبل الرنا
يمكن أن يكون الحمض النووي الريبوزي (الرنا) منظمًا مهمًا للنشاط الجيني، عن طريق الرنا الميكروي أو الرنا المضاد للدلالة أو الرنا غير المُشفر الطويل. يختلف الرنا غير المشفر الطويل عن الرنا الميكروي بأن لكل منهما مواقع ووظائف تحت خلوية نوعية. اكتشف جزيئات الرنا في البداية ضمن النواة وفي الكروماتين، وتبين أن مواقعها ووظائفها متنوعة. يبقى بعضها في الكروماتين حيث يتفاعل مع البروتينات. أما الرنا غير المشفر الطويل فيؤثر على التعبير الجيني في الاضطرابات العصبية مثل داء باركنسون وداء هنتنغتون وداء ألزهايمر، أما البقية، مثل، الناسخات غير المشفِرة الغنية بالبيريميدين، فتلعب دورًا في سرطان الرئة. وبالنظر إلى دورها في المرض، فإن جزيئات الرنا غير المشفِر الطويل واسمات حيوية محتملة قد تكون مفيدة إذا استخدمت باعتبارها هدفًا للأدوية أو العلاج الجيني، على الرغم من عدم وجود أدوية مرخصة تستهدفها حتى الآن. لا يُعرف العدد الدقيق للرنا غير المشفر الطويل في الجينوم البشري، وتتراوح التقديرات المتوفرة من 16,000 إلى 100,000 جين.[4]
مراجع
- Griffiths, Anthony J.F.؛ Wessler, Susan R.؛ Carroll, Sean B.؛ Doebley, John (2015)، An Introduction to Genetic Analysis (ط. 11)، Freeman, W.H. & Company، ص. 400–412، ISBN 9781464109485.
- Jaenish R., Bird A., 2003. "Epigenetic regulation of gene expression: how the genome integrates intrinsic and environmental signals", Nature genetics supplement, 33(245:254)
- Bell JT, Pai AA, Pickrell JK, Gaffney DJ, Pique-Regi R, Degner JF, وآخرون (2011)، "DNA methylation patterns associate with genetic and gene expression variation in HapMap cell lines"، Genome Biology، 12 (1): R10، doi:10.1186/gb-2011-12-1-r10، PMC 3091299، PMID 21251332.
- Vertino PM, Spillare EA, Harris CC, Baylin SB (أبريل 1993)، "Altered chromosomal methylation patterns accompany oncogene-induced transformation of human bronchial epithelial cells" (PDF)، Cancer Research، 53 (7): 1684–9، PMID 8453642.
 بوابة الكيمياء الحيوية
بوابة الكيمياء الحيوية بوابة علم الأحياء
بوابة علم الأحياء بوابة علم الأحياء الخلوي والجزيئي
بوابة علم الأحياء الخلوي والجزيئي بوابة علم الوراثة
بوابة علم الوراثة
| مقدمة لعلم الوراثة |  | ||||
|---|---|---|---|---|---|
| النسـخ |
| ||||
| الترجمـة |
| ||||
| مراقبة علم التخلق |
| ||||
| لمحة عامة |
| ||||||
|---|---|---|---|---|---|---|---|
| هندسة |
| ||||||
| |||||||