تعبير جيني
التعبير الجيني (ملاحظة 1) هو العملية التي يتم من خلالها استخدام المعلومات الجينية لاصطناع منتجات جينية الوظيفية. هذه المنتجات قد تكون بروتينات (يتم صنعها باستخدام المعلومات من الرنا الرسول)، أو قد تكون أحد الأنواع العديدية من الأحماض النووية الريبوزية، مثل الرنا الريبوسومي (rRNA)، الرنا الناقل (tRNA) والرنا النووي الصغير (snRNA).
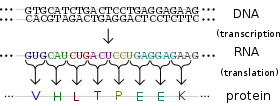
تتم عملية التعبير الجيني في كل صنف نعرفه من أصناف الحياة -حقيقيات النوى وبدائيات النوى، حتى الفيروسات تستغلها لتتضاعف- لتخليق الآلات الجزيئية في الخلية.
يمكن تعديل وضبط العديد من خطوات التعبير الجيني، كالنسخ، التوصيل، ترجمة، وتعديل البروتين ما بعد الترجمة. ضبط التعبير الجيني يسمح للخلية بالتحكم ببنيتها ووظيفتها، حيث أنه يُشَكّل أساس عمل التمايز الخلوي، التشكل الحيوي، وقدرة الكائن الحي على التكيف.
الوظيفة
يستخدم عملية التعبير الجيني المعروف في جميع الكائنات الحية: حقيقيات النوى (بما في ذلك الكائنات متعددة الخلايا)، بدائيات النوى (البكتيريا والبكتيريا العتيقة)، وتعبيرات جينية تستخدمها الفيروسات - لتوليد جزيئات وآليات للحياة. قد يكون التعبير الجيني منظما لعدة خطوات في عملياته الحيوية، بما في ذلك النسخ من الحمض النووي الريبي، والربط بين منتجات من الحمض النووي الريبي، وكذلك الترجمة منه بغرض إنتاج بروتينات معينة.[1] تنظيم الجينات سيمكنهم من السيطرة على تركيب الخلية ووظائفها، ويعد التعبير الجيني أساسا لتمايز الخلايا، من حيث التشكل والتنوع والقدرة على التكيف في أي كائن حي. تنظيم الجينات قد يكون أيضاً بمثابة الركيزة للتغيير التطوري، منذ السيطرة على توقيت والموقع ( توقيت النمو والبلوغ والشيب، وموقع الجينات في الدنا، وكمية من التعبير الجيني يمكن أن يكون لها تأثير عميق على الوظائف (الإجراءات) التي تؤديها الجينات في خلية حية أو في كائن متعدد الخلايا.[2]
علم الوراثة

في علم الوراثة، والتعبير الجيني هو المستوى الأكثر أساسية في التركيب الوراثي الذي يؤدي إلى النمط الظاهري. الشفرة الوراثية المخزنة في الحمض النووي هو «تفسير» حسب التعبير الجيني،[3][4] وخصائص التعبير تؤدي إلى النمط الظاهري للكائن. وغالبا ما يتم التعبير عن هذه الظواهر عن طريق تخليق البروتينات التي تتحكم في شكل الكائن الحي، أو أن يكون بمثابة تحفيز الانزيمات المسارات الأيضية المحددة التي تميز الكائن الحي.[5]
تعبير مغاير
التعبير المغاير Heterologous expression هو التعبير عن جين أو جزء من الجين في كائن حي مضيف، الذي ليس لديه طبيعيًا هذا الجين أو أو جزء من الجين. ويتم هذا بشكل أساسي في تقنية الحمض النووي المأشوب.
الآلية
النسخ
يعني النسخ إنتاج نسخة عن سلسلة الدنا، وتُنجزه إنزيمات بوليميراز الرنا، التي تضيف نكليوتيد ريبوزي إلى سلسلة الرنا النامية تبعًا لقانون تكامل القواعد النكليوتيدية (تتام القواعد النكليوتيدية). يُكمّل الرنا الناتج عن النَسخ السلسلة القالب 3′ ← 5′، باستثناء الثايمين الذي يُستبدل باليوراسيل في الرنا.[6]
في بدائيات النوى، يُنجَز النسخ باستعمال نوع واحد من بوليميراز الرنا، الذي يجب أن يندمج بتتالٍ نكليوتيدي في الدنا يدعى صندوق بربنو بمساعدة العامل سيغما، لبدء النسخ. في حقيقيات النوى، تنفذ عملية النسخ ضمن النواة باستعمال ثلاثة أنواع من بوليميراز الرنا يحتاج كل واحد منها إلى تتالٍ محدد في الدنا يدعى المُحفّز ومجموعة من البروتينات الرابطة للدنا (عوامل النسخ) لبدء العملية. بوليميراز الرنا الأول مسؤول عن نسخ جينات الرنا الريبوزومي. ينسخ بوليميراز الرنا الثاني كل الجينات الرامزة للبروتينات بالإضافة إلى بعض الرنا غير الرامز (الرنا النووي الصغير والرنا النُوَيي الصغير والرنا الطويل غير الرامز). ينسخ بوليميراز الرنا الثالث الرنا الريبوزومي 5 إس وجينات الرنا الناقل، وبعض أنواع الرنا الصغير غير الرامز (مثل 7 إس كي). ينتهي النسخ عندما يصل البوليميراز إلى تسلسل يدعى المنهي.
معالجة الرنا الرسول
في خلايا بدائيات النوى، يؤدي نسخ الجينات الرامزة للبروتين إلى تكوين رنا رسول جاهز للترجمة إلى بروتينات، أما في الخلايا حقيقيات النوى، ينتج عن نسخ الجينات نسخة أولية من الرنا (ما قبل رنا)، عليها أن تخضع أولًا إلى سلسلة من التعديلات لتصبح رنا ناضجًا. تتنوع أنواع وخطوات عمليات النضج بين ما قبل الرنا الرامز وغير الرامز، ورغم أن ما قبل الرنا بنوعيه الرسول والناقل يخضعان للتضفير، تختلف الآلية والخطوات المتبعة. معالجة الرنا غير الرامز موصوفة لاحقًا (معالجة الرنا غير الرامز).[7]
تتضمن معالجة ما قبل الرنا الرسول، التقبيع من الناحية 5′، وهو مجموعة من تفاعلات إنزيمية تضيف ميثيل غوانوسين-7 (قبعة) إلى النهاية 5′ في ما قبل الرنا الرسول، وبذلك تحمي الرنا من التلف بتأثير إنزيمات النوكلياز الخارجية. ترتبط القبعة بعد ذلك بالدايمر المتغاير للمعقد الرابط للقبعة (CBC20/CBC80) الذي يساعد في خروج الرنا الرسول إلى الهيولى ويحمي الرنا من نزع القبعة.[8]
يمثل قص 3′ والتذييل بعديد الأدينيلات عمليتا تعديل أخريان. تحدث هاتان العمليتان إذا كان تسلسل إشارة التذييل بعديد الأدينيلات (5′-AAUAAA-3′) موجودًا ضمن ما قبل الرنا الرسول الذي يكون عادة بين التسلسل الرامز للبروتين والتسلسل المنهي. يُقطع ما قبل الرنا الرسول أولًا ثم تُضاف سلسلة من قرابة 200 أدينين لتشيل الذيل عديد الأدينيلات، الذي يحمي الرنا من التلف. يرتبط الذيل عديد الأدينيلات بالبروتينات الرابطة لعديد الأدينيلات اللازمة لخروج الرنا الرسول وبدء الترجمة مجددًا. في العملية المعاكسة التي تدعى نزع الأدينيلات، يَقصُر الذيل عديد الأدينيلات بتأثير إنزيم النوكلياز الخارجي CCR4-Not 3′-5′، الذي يؤدي عادة إلى تخرب النَسخ بالكامل.[9]
من التعديلات المهمة التي تطرأ على ما قبل الرنا الرسول في حقيقيات النوى تضفير الرنا. غالبية ما قبل الرنا الرسول في حقيقيات النوى مكون من قطع متبدلة تدعى الإكسونات والإنترونات. خلال عملية التضفير، يحفز معقد محفِز مكون من الرنا والبروتين يدعى جسيم التضفير تفاعلَي أسترة تبادلية، مؤديًا إلى إزالة إنترون وتحرره في هيئة تشبه الوَهق، ثم يضفر الإكسونات المتجاورة معًا. في بعض الحالات، يمكن إزالة بعض الإنترونات والإكسونات أو إبقاؤها في الرنا الرسول الناضج. يخلق هذا الأمر المسمى التضفير البديل (الوصل البديل) سلسلة من النسخ المختلفة التي تنشأ عن جين واحد. ولأن هذه النسخ قابلة للترجمة إلى بروتينات مختلفة، يوسع التضفير تعقيد التعبير الجيني في الخلايا حقيقية النوى وحجم البروتيوم (المحتوى البروتيني) في النوع.[10]
قد تكون المعالجة الكبيرة للرنا ميزة تطورية مكنتها النوى في حقيقيات النوى. في بدائيات النوى، تحدث عمليتا النسخ والترجمة معًا، بينما في حقيقيات النوى، يفصل الغشاء النووي العمليتين، ما يوفر وقتًا لعمليات معالجة الرنا.[11]
نضج الرنا غير الرامز
عند معظم الكائنات الحية، تُنسخ الجينات غير الرامزة بصفتها طلائع تخضع لمعالجة إضافية. يُنسخ الرنا الريبوزومي في البداية على هيئة ما قبل رنا ريبوزومي يضم واحدًا أو أكثر من الرنا الريبوزومي. يُقطع ما قبل الرنا ويُعدَل (مَثيلة وتشكل سودويوريدين في 2′-أو) في مواقع محددة من قبل نحو 150 نوعًا من الرنا الصغير المحصور بالنويات، الذي يدعى الرنا الصغير النُوَيي. يرتبط الرنا الصغير النويي بالبروتينات مشكًلا معقدات رنا صغير نويي وبروتين. يقترن الرنا الصغير في هذه المعقدات على أساس قاعدي بالرنا الهدف وبذلك ينجز التعديل في موقع دقيق، ويؤدي الجزء البروتيني من المعقد تفاعلًا محفزًا. في حقيقيات النوى، يقطع معقد خاص (رنا صغير نويي وبروتين) يدعى «ريبونوكلياز إم آر بّي» ما قبل الرنا الريبوزومي 45 إس إلى قطعة 28 إس، وقطعة 5.8 إس، وقطعة 18 إس من الرنا الريبوزومي. تشكل عوامل معالجة الرنا والرنا الريبوزومي تجمعات كبيرة تسمى النويات.[12]
انظر أيضا
مراجع
- Brueckner F, Armache KJ, Cheung A؛ وآخرون (فبراير 2009)، "Structure–function studies of the RNA polymerase II elongation complex"، Acta Crystallogr. D Biol. Crystallogr.، 65 (Pt 2): 112–20، doi:10.1107/S0907444908039875، PMC 2631633، PMID 19171965.
{{استشهاد بدورية محكمة}}: Explicit use of et al. in:|مؤلف=(مساعدة)صيانة CS1: أسماء متعددة: قائمة المؤلفون (link) - Sirri V, Urcuqui-Inchima S, Roussel P, Hernandez-Verdun D (يناير 2008)، "Nucleolus: the fascinating nuclear body"، Histochem. Cell Biol.، 129 (1): 13–31، doi:10.1007/s00418-007-0359-6، PMC 2137947، PMID 18046571.
{{استشهاد بدورية محكمة}}: صيانة CS1: أسماء متعددة: قائمة المؤلفون (link) - Frank DN, Pace NR (1998)، "Ribonuclease P: unity and diversity in a tRNA processing ribozyme"، Annu. Rev. Biochem.، 67: 153–80، doi:10.1146/annurev.biochem.67.1.153، PMID 9759486.
- Ceballos M, Vioque A (2007)، "tRNase Z"، Protein Pept. Lett.، 14 (2): 137–45، doi:10.2174/092986607779816050، PMID 17305600.
- Weiner AM (أكتوبر 2004)، "tRNA maturation: RNA polymerization without a nucleic acid template"، Curr. Biol.، 14 (20): R883–5، doi:10.1016/j.cub.2004.09.069، PMID 15498478.
- "Structure-function studies of the RNA polymerase II elongation complex"، Acta Crystallographica D، 65 (Pt 2): 112–20، فبراير 2009، doi:10.1107/S0907444908039875، PMC 2631633، PMID 19171965.
- Krebs, Jocelyn E. (02 مارس 2017)، Lewin's genes XII، Goldstein, Elliott S.,, Kilpatrick, Stephen T.، Burlington, MA، ISBN 978-1-284-10449-3، OCLC 965781334.
- "Nucleolus: the fascinating nuclear body"، Histochemistry and Cell Biology، 129 (1): 13–31، يناير 2008، doi:10.1007/s00418-007-0359-6، PMC 2137947، PMID 18046571.
- "Ribonuclease P: unity and diversity in a tRNA processing ribozyme"، Annual Review of Biochemistry، 67: 153–80، 1998، doi:10.1146/annurev.biochem.67.1.153، PMID 9759486.
- "tRNA maturation: RNA polymerization without a nucleic acid template"، Current Biology، 14 (20): R883–5، أكتوبر 2004، doi:10.1016/j.cub.2004.09.069، PMID 15498478.
- "tRNase Z"، Protein and Peptide Letters، 14 (2): 137–45، 2007، doi:10.2174/092986607779816050، PMID 17305600.
- "Cis-acting determinants of asymmetric, cytoplasmic RNA transport"، RNA، 13 (5): 625–42، مايو 2007، doi:10.1261/rna.262607، PMC 1852811، PMID 17449729.
- "Gene Expression"، قاموس المعاني، مؤرشف من الأصل في 12 أكتوبر 2019، اطلع عليه بتاريخ 10/2019.
{{استشهاد ويب}}: تحقق من التاريخ في:|تاريخ الوصول=(مساعدة)
وصلات إضافية
- "Genes & Gene Expression"، The Virtual Library of Biochemistry and Cell Biology، BioChemWeb.org، 04 ديسمبر 2005، مؤرشف من الأصل في 02 ديسمبر 2014، اطلع عليه بتاريخ 10 يونيو 2008.
- John Kryk (28 مايو 2008)، "DNA makes RNA"، مؤرشف من الأصل في 14 مايو 2019، اطلع عليه بتاريخ 10 يونيو 2008.
- "Advancing Gene Expression Studies"، Genetic Engineering & Biotechnology News، Mary Ann Liebert, Inc.، 01 أغسطس 2008، مؤرشف من الأصل في 21 ديسمبر 2008.
- "Optimizing Transient Gene Expression"، Genetic Engineering & Biotechnology News، Mary Ann Liebert, Inc.، 01 مارس 2008، مؤرشف من الأصل في 21 ديسمبر 2008.
| مقدمة لعلم الوراثة |  | ||||
|---|---|---|---|---|---|
| النسـخ |
| ||||
| الترجمـة |
| ||||
| مراقبة علم التخلق |
| ||||
 بوابة علم الأحياء الخلوي والجزيئي
بوابة علم الأحياء الخلوي والجزيئي بوابة علم الأحياء
بوابة علم الأحياء