انقراض البرمي-الثلاثي
انقراض العصر البرمي الثلاثي المعروف باسم الموت العظيم[3] هو انقراض جماعي وقع قبل 251.4 مليون سنة،[4] في الفترة بين العصرين الجيولوجيين البرمي والثلاثي. كان هذا الحدث أكبر حالة انقراض شهدتها الأرض، والذي قضى على نحو 96% من كل أنواع الأحياء البحرية،[5] وانقراض نحو 70% من كل أنواع الفقاريات الأرضية.[6]
| انقراض البرمي-الثلاثي | |
|---|---|
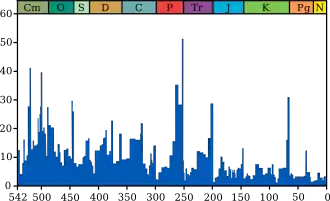
يبين الرسم البياني الأزرق نسبة واضحة (ليس عدد مطلق) لأجناس الحيوانات البحرية المنقرضة خلال أي فترة زمنية معينة. ولا تمثل جميع الأنواع البحرية، بل فقط التي تحجرت بسهولة. التسميات لأحداث الانقراض "الخمسة الكبرى" هي روابط قابلة للنقر؛ لمزيد من التفاصيل انظر احداث الانقراض.(معلومات المصدر والصورة)
| |
| نوع الانقراض | انقراض جماعي |
| عصر الانقراض | البرمي - الثلاثي |
| تاريخ الانقراض | قبل 251.4 مليون سنة |
| سبب الانقراض | تغير تدريجي للمناخ اصطدم نيزك متفجر انفجارات بركانية كبيرة |
| نسبة الكائنات المنقرضة |
96% من الأنواع البحرية 70% من كل أنواع الفقاريات الأرضية. |
كما يُعد هذا الانقراض هو الانقراض الجماعي الوحيد المعروف للحشرات.[7] انقرضت نحو 57% من كل العائلات و83% من جميع الأجناس. نظرًا لفقد الكثير من التنوع البيولوجي، استغرق الأمر وقتًا أطول لعودة الحياة على الأرض مقارنةً بحالات الانقراض الأخرى.[5] وقد وصفت هذا الحدث بأنه «أم كل الانقراضات الشاملة».[8]
يعتقد الباحثون أن هذا الانقراض تم من خلال مرحلة إلى ثلاث مراحل.[4][6][9][10] هناك العديد من الآليات المقترحة لحدوث هذا الانقراض، أولى مراحل الانقراض سببه حدوث تغير بيئي تدريجي، في حين المرحلة التالية ترجع إلى حدوث كارثة. المراحل الأخيرة تشمل اصطدام نيزك كبير أو نيازك متعددة، مع ثورات البراكين الضخمة ونيران الفحم أو الغاز والإنفجارات من مصاطب سيبيريا[11]، واندفاع غاز الميثان من قاع المحيطات مسببًا تأثير البيت الزجاجي. شملت أيضًا التغيرات التدريجية تغير مستوى سطح البحر وحدوث نقص في الأكسجين وزيادة التصحر، والتغير في دوران المحيطات بسبب التغيرات المناخية.
أنماط الانقراض
اللافقاريات البحرية
تكبدت اللافقاريات البحرية أكبر الخسائر خلال انقراض العصر البرمي الثلاثي. وعثر على أدلة على ذلك في عينات من أقسام جنوب الصين على حدود العصر البرمي الثلاثي. هنا اختفى 286 من أصل 329 جنسًا من اللافقاريات البحرية داخل المنطقتين الرسوبية الأخيرتين اللتين احتوتا على مخروطيات الأسنان من العصر البرمي. وربما كان الانخفاض في التنوع ناجمًا عن زيادة حادة في حالات الانقراض، بدلًا من انخفاض في الانتواع.[12][13]
أثر الانقراض في المقام الأول على الكائنات الحية ذات الهياكل العظمية المكونة من كربونات الكالسيوم، وخاصة تلك التي تعتمد على مستويات ثابتة من ثاني أكسيد الكربون لإنتاج هياكلها العظمية. كانت هذه الكائنات الحية عرضة لتأثيرات تحمض المحيطات الناتجة عن زيادة ثاني أكسيد الكربون في الغلاف الجوي.[14]
بين الكائنات القاعية، ضاعف حدث الانقراض معدلات الانقراض الطبيعي، وبالتالي تسبب في الحد الأقصى من فقدان الأنواع إلى الأصنوفات التي لديها معدل انقراض مرتفع طبيعي (ضمنيًا، الأصناف ذات معدل دوران مرتفع). كان معدل انقراض الكائنات البحرية كارثيًا.[15]
تضمنت مجموعات اللافقاريات البحرية الباقية على قيد الحياة عضديات الأرجل (تلك التي لها مفصل)، والتي شهدت انخفاضًا بطيئًا في أعدادها منذ الانقراض البرمي الثلاثي؛ رتب السيراتيتيدا للأمونيت؛ و (زنابق البحر)، التي كادت أن تنقرض ولكنها أصبحت وفيرة ومتنوعة فيما بعد.
كانت المجموعات التي معدلات بقائها على قيد الحياة أعلى تتمتع عمومًا بالتحكم النشط في الدوران، وآليات تبادل الغازات المعقدة، والتكلس الخفيف؛ عانت الكائنات الحية الأكثر تكلسًا ذات أجهزة التنفس الأبسط من خسارة أكبر في تنوع الأنواع. في حالة عضديات الأرجل، على الأقل، كانت الأصنوفات الباقية بشكل عام صغيرة ونادرة من مجتمع متنوع سابقًا.
عانت الأمونيت، التي كانت في حالة تدهور طويل الأمد لمدة 30 مليون سنة منذ الرودي (منتصف العصر البرمي)، من انقراض انتقائي قبل 10 ملايين سنة من الحدث الرئيسي، في نهاية مرحلة الكابتاني. في هذا الانقراض الأولي، الذي قلل بشكل كبير من التفاوت، أو من مجموعة النباتات البيئية المختلفة، كانت العوامل البيئية هي المسؤولة على ما يبدو. انخفض التنوع والتفاوت أكثر حتى حدود العصر البرمي- الثلاثي. كان الانقراض في(العصر البرمي-الثلاثي) غير انتقائي، بما يتفق مع منشئ الفاجعة. خلال العصر الثلاثي، ارتفع التنوع بسرعة، لكن التفاوت ظل منخفضًا.[16]
أصبح نطاق المورفوسبيس الذي تشغله الأمونيت، أي نطاق أنواعها أو أشكالها أو هياكلها المحتملة، أكثر تقييدًا مع تقدم العصر البرمي. بعد بضعة ملايين من السنين في العصر الثلاثي، عادت هياكل الأمونيت لتشغل نطاقها الأصلي مرة أخرى، ولكن جرى تقاسم البارامترات بشكل مختلف بين الأفرع الحيوية.[17]
اللافقاريات الأرضية
كان لدى العصر البرمي تنوع كبير في الحشرات وأنواع اللافقاريات الأخرى، بما في ذلك أكبر الحشرات الموجودة على الإطلاق. تُعد نهاية العصر البرمي أكبر انقراض جماعي معروف للحشرات؛ ووفقًا لبعض المصادر، فهو الانقراض الجماعي الوحيد للحشرات. انقرضت ثمانية أو تسعة رتب حشرية وانخفض تنوع عشرة أخرى بشكل كبير. بدأت شبكيات الأجنحة القديمة (وهي الحشرات التي تمتلك أجزاء فموية ثاقبة وماصة) في الانخفاض خلال منتصف العصر البرمي، وارتبطت هذه الانقراضات بتغيير في النباتات. حدث الانخفاض الأكبر في أواخر العصر البرمي وربما لم يكن سببه المباشر التحولات الزهرية المرتبطة بالطقس.[18]
أغلب مجموعات الحشرات الأحفورية التي عُثر عليها بعد حدود العصر البرمي-الثلاثي تختلف اختلافًا كبيرًا عن تلك التي عُثر عليها من قبل: فمن بين مجموعات حشرات حقبة الحياة القديمة، لم يكتشف في الرواسب من بعد الانقراض سوى الغلوسيليتروديا، والميوموبتيرا، ومستقيمات الأجنحة البدائية. انقرضت كل من خارجيات الأجنحة، وأحاديات الذنب، وشبكيات الأجنحة القديمة، وسرغاميات، واليعسوبيات الضخمة بنهاية العصر البرمي. وفي رواسب أواخر العصر الثلاثي الموثقة توثيقًا جيدًا، تألفت الحفريات بشكل ساحق من مجموعات الحشرات الأحفورية الحديثة.[19]
النباتات الأرضية
استجابة النظام البيئي للنبات
السجل الجيولوجي للنباتات الأرضية متناثر ويعتمد في الغالب على دراسات حبوب اللقاح والجراثيم. النباتات محصنة نسبيًا ضد الانقراض الجماعي، مع تأثير جميع الانقراضات الجماعية الكبرى «غير الملحوظ» على مستوى السلالة. وحتى الانخفاض الملحوظ في تنوع الأنواع (بنسبة 50%) قد يكون راجعًا في معظمه إلى العمليات التصنيفية. ومع ذلك، حدثت إعادة ترتيب هائلة للنظم البيئية، مع تغيرات عميقة في وفرة النباتات وتوزيعاتها واختفاء جميع الغابات تقريبًا؛ وبالكاد نجت نباتات حقبة الحياة القديمة من هذا الانقراض.
عند حدود العصر البرمي-الثلاثي، تغيرت المجموعات الزهرية المهيمنة، مع دخول العديد من مجموعات النباتات البرية إلى تدهور مفاجئ، مثل كوردايت (عاريات البذور)، والسرخس اللساني (سرخسيات بذرية). استبدِل بأجناس عاريات البذور السائدة بعد الحدود نباتات ذئبية والتي وجِدت لإعادة استعمار المناطق المضطربة.[20]
تشير دراسات علم الطلع أو دراسات حبوب اللقاح من شرق جرينلاند لطبقات الصخور الرسوبية التي وضِعت خلال فترة الانقراض إلى وجود غابات قليلة الكثافة لعاريات البذور قبل وقوع الحدث. في نفس الوقت الذي تراجعت فيه الحيوانات البحرية الكبيرة اللافقارية، ماتت هذه الغابات القليلة الكثافة وأعقبها ارتفاع في تنوع النباتات العشبية الأصغر بما في ذلك النباتات الذئبية، وكل من كفعان وإسويطيات. في وقت لاحق، أصبحت مجموعات أخرى من عاريات البذور مهيمنة مرة أخرى ولكنها عانت مرة أخرى من موت كبير. حدثت هذه التحولات في النباتات الدورية عدة مرات خلال فترة الانقراض وبعدها. تشير هذه التقلبات في النباتات السائدة بين الأصناف الخشبية والعشبية إلى ضغوط بيئية مزمنة أدت إلى فقدان معظم أنواع نباتات الغابات القليلة الكثافة. لم تتطابق حالات التعاقب والانقراض في المجتمعات النباتية مع التحول في قيم دلتا سي 13 (δ13C) ولكنها حدثت بعد سنوات عديدة. استغرقت استعادة غابات عاريات البذور من 4 إلى 5 ملايين سنة.[21]
فجوة الفحم
لا توجد رواسب للفحم معروفة من أوائل العصر الثلاثي، وتلك الموجودة في العصر الثلاثي الأوسط رقيقة ومنخفضة. شُرحت «فجوة الفحم» بعدة طرق. وقد أشير إلى أن الفطريات والحشرات والفقاريات الجديدة الأكثر عدوانية قد تطورت وقتلت أعدادًا هائلة من الأشجار. عانت هذه المحللات نفسها من خسائر فادحة في الأنواع أثناء الانقراض ولا تعتبر سببًا محتملًا لفجوة الفحم. يمكن ببساطة أن تكون جميع النباتات التي تشكل الفحم قد انقرضت بسبب انقراض العصر البرمي الثلاثي، وأن الأمر استغرق 10 ملايين سنة لتتكيف مجموعة جديدة من النباتات مع الظروف الرطبة والحمضية لرخاخ الخث. العوامل اللاأحيائية (العوامل التي لا تسببها الكائنات الحية)، مثل انخفاض هطول الأمطار أو زيادة مدخلات الرواسب الفتاتية، قد تكون مسؤولة أيضًا.[22]
من ناحية أخرى، قد يعكس نقص الفحم ندرة جميع الرواسب المعروفة من أوائل العصر الثلاثي. وربما تكون النظم البيئية المنتجة للفحم، بدلًا من اختفائها، قد انتقلت إلى المناطق التي ليس لدينا فيها سجل رسوبي في أوائل العصر الثلاثي. ففي شرق أستراليا، على سبيل المثال، ظل المناخ البارد هو المعيار لفترة طويلة، مع تكيف النظام البيئي للخث مع هذه الظروف. انقرض ما يقرب من 95% من هذه النباتات المنتجة للخث محليًا عند حدود العصر البرمي الثلاثي، واختفت رواسب الفحم في أستراليا وأنتاركتيكا بشكل ملحوظ قبل حدود العصر البرمي الثلاثي.[23]
انظر أيضًا
مراجع
- Munnecke, A.؛ Samtleben, C.؛ Bickert, T. (2003)، "The Ireviken Event in the lower Silurian of Gotland, Sweden-relation to similar Palaeozoic and Proterozoic events"، Palaeogeography, Palaeoclimatology, Palaeoecology، 195 (1): 99–124، doi:10.1016/S0031-0182(03)00304-3.
- Jeppsson, L.؛ Calner, M. (2007)، "The Silurian Mulde Event and a scenario for secundo—secundo events"، Earth and Environmental Science Transactions of the Royal Society of Edinburgh، 93 (02): 135–154، doi:10.1017/S0263593300000377.
- Barry, Patrick L. (28 يناير 2002)، "The Great Dying"، Science@NASA، Science and Technology Directorate, Marshall Space Flight Center, NASA، مؤرشف من الأصل في 16 فبراير 2012، اطلع عليه بتاريخ 26 مارس 2009.
- Jin YG, Wang Y, Wang W, Shang QH, Cao CQ, Erwin DH (2000)، "Pattern of Marine Mass Extinction Near the Permian–Triassic Boundary in South China"، Science، 289 (5478): 432–436، doi:10.1126/science.289.5478.432، PMID 10903200.
{{استشهاد بدورية محكمة}}: صيانة CS1: أسماء متعددة: قائمة المؤلفون (link) - Benton M J (2005)، When Life Nearly Died: The Greatest Mass Extinction of All Time، Thames & Hudson، ISBN 978-0500285732.
- Sahney, S. and Benton, M.J. (2008)، "Recovery from the most profound mass extinction of all time" (PDF)، Proceedings of the Royal Society: Biological، 275 (1636): 759، doi:10.1098/rspb.2007.1370، PMC 2596898، PMID 18198148، مؤرشف من الأصل (PDF) في 12 أغسطس 2018.
{{استشهاد بدورية محكمة}}: صيانة CS1: أسماء متعددة: قائمة المؤلفون (link) - Sole, R. V., and Newman, M., 2002. "Extinctions and Biodiversity in the Fossil Record - Volume Two, The earth system: biological and ecological dimensions of global environment change" pp. 297-391, Encyclopedia of Global Environmental Change John Wiley & Sons.
- Erwin DH (1993)، The great Paleozoic crisis; Life and death in the Permian، Columbia University Press، ISBN 0231074670.
- Yin H, Zhang K, Tong J, Yang Z, Wu S، "The Global Stratotype Section and Point (GSSP) of the Permian-Triassic Boundary"، Episodes، 24 (2): 102–114.
{{استشهاد بدورية محكمة}}: صيانة CS1: أسماء متعددة: قائمة المؤلفون (link) - Yin HF, Sweets WC, Yang ZY, Dickins JM,، "Permo-Triassic Events in the Eastern Tethys"، Cambridge Univ. Pres, Cambridge, 1992.
{{استشهاد بدورية محكمة}}: صيانة CS1: extra punctuation (link) صيانة CS1: أسماء متعددة: قائمة المؤلفون (link) - Darcy E. Ogdena and Norman H. Sleep (2011)، "Explosive eruption of coal and basalt and the end-Permian mass extinction."، Proceedings of the National Academy of Sciences of the United States of America، 109: 59–62، Bibcode:2012PNAS..109...59O، doi:10.1073/pnas.1118675109، مؤرشف من الأصل في 19 فبراير 2019.
- Dolenec T, Lojen S, Ramovs A (2001)، "The Permian–Triassic boundary in Western Slovenia (Idrijca Valley section): Magnetostratigraphy, stable isotopes, and elemental variations"، Chemical Geology، 175 (1): 175–190، Bibcode:2001ChGeo.175..175D، doi:10.1016/S0009-2541(00)00368-5.
- Musashi M, Isozaki Y, Koike T, Kreulen R (2001)، "Stable carbon isotope signature in mid-Panthalassa shallow-water carbonates across the Permo–Triassic boundary: Evidence for 13C-depleted ocean"، Earth and Planetary Science Letters، 193 (1–2): 9–20، Bibcode:2001E&PSL.191....9M، doi:10.1016/S0012-821X(01)00398-3.
- Visscher H, Brinkhuis H, Dilcher DL, Elsik WC, Eshet Y, Looy CW, Rampino MR, Traverse A (1996)، "The terminal Paleozoic fungal event: Evidence of terrestrial ecosystem destabilization and collapse"، Proceedings of the National Academy of Sciences، 93 (5): 2155–2158، Bibcode:1996PNAS...93.2155V، doi:10.1073/pnas.93.5.2155، PMC 39926، PMID 11607638.
- Foster, C.B.؛ Stephenson, M.H.؛ Marshall, C.؛ Logan, G.A.؛ Greenwood, P.F. (2002)، "A revision of Reduviasporonites Wilson 1962: Description, illustration, comparison and biological affinities"، Palynology، 26 (1): 35–58، doi:10.2113/0260035.
- Twitchett RJ, Looy CV, Morante R, Visscher H, Wignall PB (2001)، "Rapid and synchronous collapse of marine and terrestrial ecosystems during the end-Permian biotic crisis"، Geology، 29 (4): 351–354، Bibcode:2001Geo....29..351T، doi:10.1130/0091-7613(2001)029<0351:RASCOM>2.0.CO;2، ISSN 0091-7613.
- Wang, X-D & Sugiyama, T. (ديسمبر 2000)، "Diversity and extinction patterns of Permian coral faunas of China"، Lethaia، 33 (4): 285–294، doi:10.1080/002411600750053853.
- Bambach, R.K.؛ Knoll, A.H.؛ Wang, S.C. (ديسمبر 2004)، "Origination, extinction, and mass depletions of marine diversity"، Paleobiology، 30 (4): 522–542، doi:10.1666/0094-8373(2004)030<0522:OEAMDO>2.0.CO;2، ISSN 0094-8373، مؤرشف من الأصل في 22 فبراير 2016.
- Jin YG, Wang Y, Wang W, Shang QH, Cao CQ, Erwin DH (2000)، "Pattern of marine mass extinction near the Permian–Triassic boundary in south China"، Science، 289 (5478): 432–436، Bibcode:2000Sci...289..432J، doi:10.1126/science.289.5478.432، PMID 10903200.
- Twitchett RJ, Looy CV, Morante R, Visscher H, Wignall PB (2001)، "Rapid and synchronous collapse of marine and terrestrial ecosystems during the end-Permian biotic crisis"، Geology، 29 (4): 351–354، Bibcode:2001Geo....29..351T، doi:10.1130/0091-7613(2001)029<0351:RASCOM>2.0.CO;2، ISSN 0091-7613.
- McKinney, M.L. (1987)، "Taxonomic selectivity and continuous variation in mass and background extinctions of marine taxa"، Nature، 325 (6100): 143–145، Bibcode:1987Natur.325..143M، doi:10.1038/325143a0، S2CID 13473769.
- Cascales-Miñana, B.؛ Cleal, C. J. (2011)، "Plant fossil record and survival analyses"، Lethaia، 45: 71–82، doi:10.1111/j.1502-3931.2011.00262.x.
- Labandeira CC, Sepkoski JJ (1993)، "Insect diversity in the fossil record"، Science، 261 (5119): 310–315، Bibcode:1993Sci...261..310L، CiteSeerX 10.1.1.496.1576، doi:10.1126/science.11536548، PMID 11536548.
| ظواهر |
|  |
|---|---|---|
| نماذج |
| |
| أسباب |
| |
| نظريات ومفاهيم |
| |
| أحداث انقراض كبيرة | ||
| انقراضات اخرى |
| |
| أنواع منقرضة |
| |
| منظمات | ||
| انظر أيضًا | ||
 بوابة علم الأحياء التطوري
بوابة علم الأحياء التطوري بوابة علوم الأرض
بوابة علوم الأرض بوابة علم الأحياء القديمة
بوابة علم الأحياء القديمة بوابة الفضاء
بوابة الفضاء بوابة الأنواع المنقرضة والمهددة بالانقراض
بوابة الأنواع المنقرضة والمهددة بالانقراض