سكوالين سينثيز
سكوالين سينثيز ويعرف اختصارا (SQS) أو فيرنزل-ثنائي الفسفات: فيرنزل-ثنائي الفسفات ناقل فيرنزل هو إنزيم يقع على غشاء الشبكة الإندوبلازمية. يشارك السكوالين سينثيز في مسار التخليق الحيوي الأيزوبينيدي، ويحفز التفاعل من خطوتين حيث يتم تحويل جزيئين متطابقين من بيروفوسفات الفيرنزل إلى السكوالين، مع استهلاك جزئ فوسفات ثنائي نيوكليوتيد الأدينين وأميد النيكوتين[2] يتم التحفيز بواسطة SQS وتعد أول خطوة الزامية في تخليق ستيرول، حيث يتم تحويل السكوالين بشكل حصري إلى ستيرولات مختلفة، مثل الكوليسترول، عبر مسار معقد متعدد الخطوات. ينتمي SQS إلى عائلة سكوالين / فايتوين سينثاز وهم من البروتينات.
| farnesyl-diphosphate farnesyltransferase 1 | |
|---|---|
| المعرفات | |
| الرمز | FDFT1 |
| أنتريه | 2222 |
| HUGO | 3629 |
| أوميم | 184420 |
| RefSeq | NM_004462 |
| يونيبروت | P37268 |
| بيانات أخرى | |
| رقم التصنيف الإنزيمي | 2.5.1.21 |
| الموقع الكروموسومي | Chr. 8 p23.1-p22 |
التنوع
يتواجد سكوالين سينثيز في الحيوانات والنباتات والخميرة.[3] من حيث البنية والميكانيك، يشبه السكوالين سينثيز بشكل وثيق فايتوين سينثيز (PHS)، نوع آخر من البرينل ترانسفيريز. يؤدي PHS دورًا مماثلًا لسكوالين سينثيز في النباتات والبكتيريا، مما يحفز توليف الفايتوين، وهي سلائف للمركبات الكاروتينية.[4]
الشكل الهيكلي والبنية
._Key_residues_in_the_central_channel_are_shown_as_spheres..png.webp)
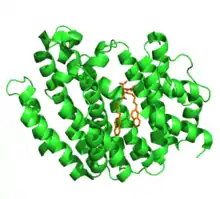
يتواجد سكوالين المصنع بشكل حصري على غشاء الشبكة الاندوبلازمية [5] ويرتكز سكوالين سينثيز على الغشاء عبر نطاق كروبني طرفي قصير يمتد عبر الغشاء [6] ويبرز النطاق النيتروجيني الطرفي التحفيزي في السايتوسول (العصاة الخلوية: مكون سائل من مكونات السيتوبلازم الموجود في الخلية) حيث تكون الركائز القابلة للذوبان مرتبطة.[2] تشكل الثديات من SQS ما يقرب من 47 كي دالتون وتتألف تقريبا من 416 من الأحماض الأمينية. تم تحديد التركيب البلوري لقياس سكوالين سينثيز البشري في عام 2000، وكشف أن البروتين يتكون بالكامل من الفا هيلكس.حيث يتم طي الانزيم في مجال واحد، ويتميز بقناة مركزية كبيرة. تقع المواقع النشطة لكل من التفاعلات النصفية المحفزة بواسطة SQS داخل هذه القناة. أحد طرفي القناة مفتوح أمام العصارة الخلوية، في حين يشكل الطرف الآخر كاره للماء ومبتعد عنه.[5] يحتوي SQS على اثنين من متواليات الأسبارتات الغنية، والتي يعتقد أنها تشارك مباشرة في الآلية التحفيزية.[7] هذه الأشكال الغنية الأسبارتية هي واحدة من العديد من السمات الهيكلية المحفوظة في الأنزيمات الحيوية الأيزوبرويدانية من الدرجة الأولى، على الرغم من أن هذه الإنزيمات لا تتشارك في تسلسل التماثل.[5]
آلية العمل
يحفز سكوالين سينثيز عملية الثنائية الجزيئية الاختزالية ل فيرنزل بايروفوسفات (FPP) حيث يتم تحويل جزيئين متماثلين من FPP إلى جزيء واحد من السكوالين. يحدث التفاعل في خطوتين، مرورا بالمركب الوسطي بريسكوالين بايروفوسفات (PSPP). و (FPP) عبارة عن مركب يحتوي على 15 ذرة كربون، في حين أن السكوالين غير قابل للذوبان، والايزوبرينويد 30 كربونه.[2][4] هذا التفاعل هو توليف التربين من الذيل إلى الذيل، لأن كلا جزيئين FPP كلاهما يرتبطان بموضع الكربونه رقم 4 ويشكلان ترابط بينهما 4-4. هذا يتناقض مع ارتباط 4-1 اذي هو أكثر شيوعا في التخليق الحيوي من 4-4 .[8][9] تتطلب آلية التفاعل من SQS جزئ كاتيون موجب الشحنة ثنائي التكافؤ، وغالبا ما يكون ايون المغنيسيوم، لتسهيل ربط مجموعات بيروفوسفات على FPP.[10]
تكثيف فيرنزل بايروفوسفات (FPP)
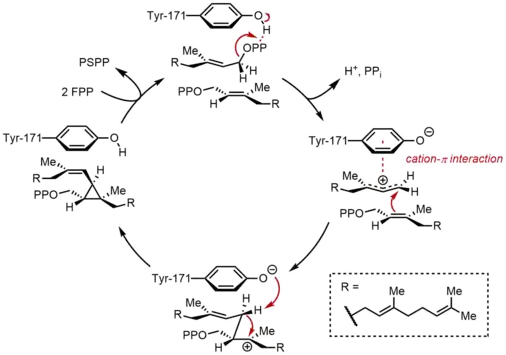
في النصف الأول من التفاعل، يرتبط جزيئين متطابقين من إلى سينزاس سكوالين (SQS) بطريقة متسلسلة. ترتبط جزيئات الـ FPP بمناطق متميزة من الإنزيم، ولها ارتباطات ملزمة مختلفة.[11] بدءا من الجزء العلوي من الدورة الحفازة أدناه، يبدأ التفاعل مع تأين FPP لتوليدكاتيون كاربوني. وتلعب بقايا التيروزين (Tyr-171) دورًا حاسمًا في هذه الخطوة من خلال العمل كمانح للبروتونات لتسهيل استخراج البيروفوسفات. علاوة على ذلك، فإن الأنيون الفينول الناتج يمكن أن يساعد على استقرار الكرب الناتجة عن طريق تفاعلات باي - كاتيون، والتي ستكون قوية بشكل خاص بسبب طبيعتها الغنية جدا بالإلكترون من الأنيون الفينول. ثم يتم الهجوم على الكاتيون الأليكلي الناتج عن أولفين من جزيء ثانٍ من FPP ، مما يوفر ارتباطًا كهربيًا ثالثًا. إن الأنيون الفينول الذي تم إنشاؤه من قبل يعمل كقاعدة لتجريد بروتون من هذا المعقد لتشكيل منتج بروبان حلقي، بروسكولين بيروفوسفات (PSPP). PSPP تم إنشاؤه لا يزال مرتبطا مع SQS للتفاعل الثاني.[5][10] تم إثبات أهمية وجود بقايا التيروزين في هذه العملية من خلال دراسات الطفرات على الفئران SQS (rSQS)، [7] وحقيقة أن Tyr-171 محفوظ في جميع SQSs المعروفة (و PHSs).[2] في rSQS ، تم تحويل Tyr-171 إلى بقايا عطرية فينيل انالين وتريبتوفان، وكذلك مخلفات تحتوي على الهيدروكسيل سيرين (حمض اميني). لم يتمكن أي من هذه المسوخات من تحويل FPP إلى PSPP أو السكوالين، مما يدل على أن الحلقات العطرية أو الكحولات وحدها غير كافية لتحويل FPP إلى PSPP.
إعادة ترتيب PSPP والحد منه
في التفاعل نصف الثاني من SQS ، ينتقل بروسكولين بيروفسفات (PSPP) إلى موقع رد فعل ثاني داخل SQS. محافظا على PSPP في القناة المركزية لـ SQS ويُعتقد أنه يحمي الوسط التفاعلي من التفاعل مع الماء.[5] من PSPP ، يتم تشكيل السكوالين عن طريق سلسلة من إعادة ترتيب الكاتيون الكربوني.[12][13] تبدأ العملية بتأين بيروفوسفاتي، مما يعطي كاتيونوبروبيلكاربنيل الكاتيوني. يعيد الأيونات ترتيبها بواسطة هجرة 1 و 2 من بروبان الحلقي C - C إلى الكاتيون الكربوني، تشكيل الرابطة هو مبين باللون الأزرق لإعطاء الالكاتيون الكربوني البيوتل الحلقي. في وقت لاحق، يحدث هجرة ل 1 و 2 ثانية لتشكيل كاتيونوبروبيلكاربنيل الكاتيوني آخر، مع الكاتيون يستريح على الكربون الثلاثي. ثم يتم فتح هذا الخلاط الناتج بعد ذلك بواسطة جزئ فوسفات ثنائي نيوكليوتيد الأدينين وأميد النيكوتين، مما يعطي السكوالين، والذي يتم إطلاقه بعد ذلك بواسطة SQS في غشاء الشبكة الإندوبلازمية.[2]
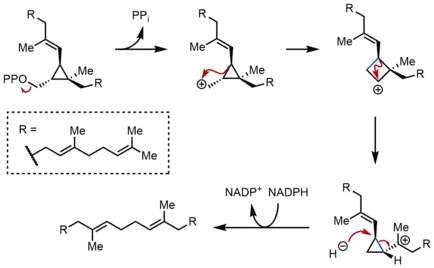
بينما يمكن إعادة ترتيب سايكلوبروبكاركارينز - سايكلوبروبلكاربينل من خلال وسيطة الكاتيون بيوتيل الحلقي بشكل منفصل، لا يمكن حصر الكاتيون بيوتيل الحلقي المفترض في الدراسات النموذجية. وهكذا، فإن الكاتيون بيوتيل الحلقي قد يكون في الواقع حالة انتقال بين الكاتيلين سايكلوبروبيلكاربينل، بدلاً من وسيط منفصل. إن الطبيعة الفراغية للوسيطة والهندسة الأوليفينية في المنتج النهائي تمليها الطبيعة السوبافية لحدوث 1 و 2 نوبات ومتطلبات الستيريو. في حين تم اقتراح آليات أخرى، فإن الآلية المبينة أعلاه مدعومة بعزل رينيلقول، وهو الكحول المتكون من احتجاز ثاني الكاتيون بيوتيل الحلقي بالماء.
التنظيم
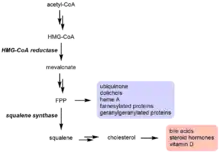
FPP عبارة عن وسيط استقلابي مهم في مسار الميفالونيت الذي يمثل نقطة فرع رئيسية في مسارات التيربونايد.[2][14] ويستخدم FPP لتشكيل عدة فئات مهمة من المركبات بالإضافة إلى الستيرولات (عبر السكوالين)، بما في ذلك اليوبيكوينون [15] والدوليكول[16] يحفز SQS الخطوة الإلزامية الأولى في التخليق الحيوي للسترول من FPP ، وبالتالي فهي مهمة للتحكم في التدفق نحو منتجات يوبيكوينون والستيرولات مقابل غير الستيرولات. ويرتبط نشاط SQS ارتباطًا وثيقًا بنشاط اختزال HMG-CoA ، الذي يحفز خطوة تحديد المعدل لمسار الميفالونيت.وتكون مستويات عالية من الكولسترول الضار للجيسم مشتقة وتمنع نشاط HMG-CoA ا، فلم يعد هناك حاجة للميفالونيت لإنتاج الستيرولات. ومع ذلك، لوحظ وجود نشاط اختزال ل HMG-CoA المتبقي حتى مع مستويات من الكوليستيرول الضار للجسم عالية جدا، بحيث يمكن صنع FPP لتشكيل منتجات غير سترولية ضرورية لنمو الخلايا.[17] لمنع استخدام هذه الـ FPP المتبقية لتكوين الستيرولات عندما تكون الستيرويد وفيرة، وينخفض نشاط SQS بشكل ملحوظ عندما تكون مستويات LDL عالية.[18] هذا القمع لنشاط SQS يفكر بشكل أفضل كآلية للتحكم في التدفق، بدلاً من طريقة لتنظيم مستويات الكوليسترول. هذا لأن HMG-CoA المختزل هو عامل التحكم الأكثر أهمية لتنظيم توليف الكوليسترول (نشاطه يثبط بنسبة 98٪ عندما تكون مستويات LDL عالية).
التنظيم من قبل الستيرولات
يحدث تنظيم SQS في المقام الأول على مستوى النسخ الجيني لسكوالسن.[2] ويعتبر عنصر بروتين ربط عنصر ستيرول (SREBP) لعوامل النسخ أمرًا أساسيًا لتنظيم الجينات المشاركة في توازن الكوليسترول، وهو مهم للتحكم في مستويات نسخ SQS. عندما تكون مستويات الستيرول منخفضة، يتم تشريح شكل خامل من SREBP لتشكيل عامل النسخ النشط، والذي ينتقل إلى النواة للحث على نسخ الجين SQS. من بين عوامل النسخ الثلاث المعروفة SREBP ، فقط SREBP-1a و SREBP-2 ينشطان نسخ الجين SQS في كبد الفأر المعدّل وراثيًا.[19][20] المزروع في خلايا HepG2 ، ويبدو SREBP-1a أكثر أهمية من SREBP-2 في التحكم في تنشيط محفز SQS.[21] ومع ذلك، فقد ثبت أن محفزات SQS تستجيب بشكل مختلف لـ SREBP-1a و SREBP-2 في أنظمة تجريبية مختلفة.
وبصرف النظر عن SREBPs ، هناك حاجة إلى عوامل نسخ ملحق لتنشيط أقصى من المروج SQS. وقد كشفت الدراسات المروجة باستخدام فحوصات جين المراسل لوسيفيريز أن عوامل النسخ Sp1 ، و NF-Y و / أو CREB مهمة أيضا لتنشيط المرشد SQS. ويكون أيضا مطلوب NF-Y و / أو CREB لـ SREBP-1a لتنشيط محفز SQS بالكامل، على الرغم من أن Sp1 مطلوب أيضًا لـ SREBP-2 للقيام بذلك.
خريطة المسار التفاعلي
اضغط على الجين, أو البروتينات أو نواتج الأيض بالأسفل لتوصلك إلى المقالة المتعلقة. [§ 1]
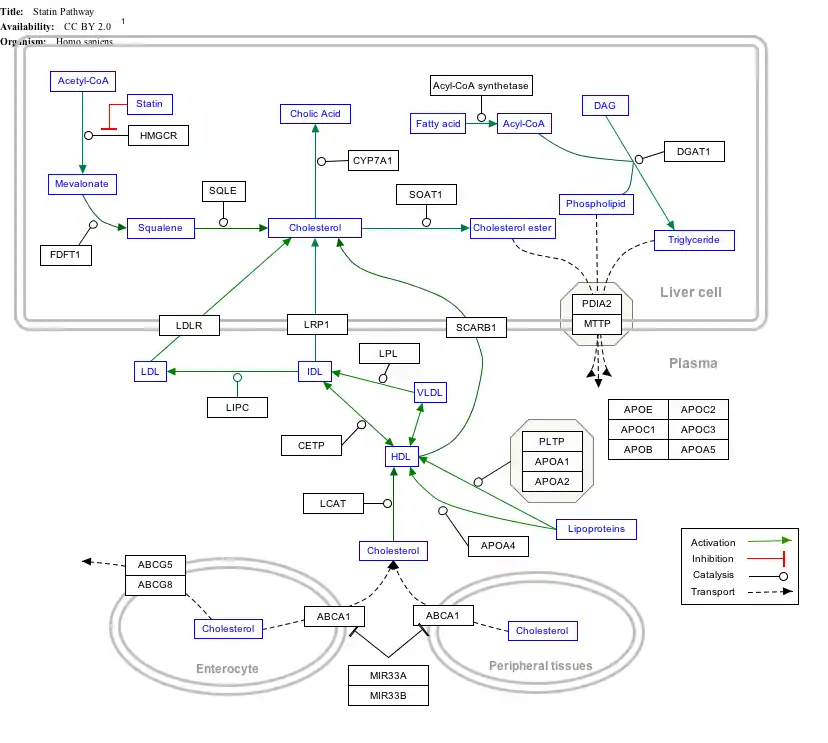
- The interactive pathway map can be edited at WikiPathways: "Statin_Pathway_WP430".
الوظيفة البيولوجية
سكوالين سينثيز (SQS) هو إنزيم يشارك في مسار التخليق الحيوي الايزوبرنويدي.و يحفز سينثيز SQS نقطة التفرع بين الاستيثاق الحيوي ستيرول والغيرسترول، ويلزم البيروفوسفات فيرنزل (FPP) حصرا لإنتاج الستيرولات.[2] من المواد المهمة التي ينتجها هذا المسار هي الكوليسترول، الذي يستخدم في أغشية الخلايا ولتخليق الهرمونات.[22] تتنافس SQS مع العديد من الإنزيمات الأخرى لاستخدام FPP ، حيث أنها مقدمة لمجموعة متنوعة من التيربينويات. ويؤدي الانخفاض في نشاط SQS إلى الحد من تدفق FPP إلى مسار الستيرول، وزيادة إنتاج منتجات غير ستيرولية. تشمل منتجات نسترول الهامة: يوبيكوينون، dolichols ، هيما الف، وبروتينات فيرنزيلية.[23]
وقد أثبت تطوير الفئران بالضربة القاضية سكوالين سينثيز أن فقدانه قاتل، وأن الإنزيم ضروري لتطور الجهاز العصبي المركزي.[24]
العلاقة المرضية
يهدف سكوالين سينثيز لتنظيم مستويات الكوليسترول. وقد تبين زيادة التعبير عن SQS لرفع مستويات الكوليسترول في الفئران.[24] لذلك، مثبطات SQS لها أهمية كبيرة في علاج فرط كوليسترول الدم والوقاية من مرض القلب التاجي (CHD).[25] وقد اقترح أيضًا أن المتغيرات في هذا الإنزيم قد تكون جزءًا من الارتباط الوراثي مع فرط كوليسترول الدم.[26]
مثيطات سكوالين سينثيز
ثبت أن مثبطات سكوالين سينثيز تقلل من تخليق الكولسترول، وكذلك لتقليل مستويات ثلاثي الجليسريد في البلازما.[22][27] قد توفر مثبطات SQS بديلا لمثبطات اختزال HMG-CoA (الستاتينات)، التي لها آثار جانبية إشكالية لبعض المرضى.[28] مثبطات سكوالين سينثيز التي تم بحثها لاستخدامها في الوقاية من أمراض القلب والأوعية الدموية وتشمل لاباكويستات (TAK-475)، وحمض الزاراجونيك، و RPR 107393.[29][30] على الرغم من الوصول إلى التجارب السريرية من المرحلة الثانية، فقد توقفت لاباكويستات بحلول عام 2008.[31][32]
يجري حاليا التحقيق في تثبيط سكوالين سينثيز للمثانة في المكورات العنقودية الذهبية كعلاج مضاد للبكتيريا.[33]
كائنات نموذجية
تم استخدام الكائنات الحية النموذجية في دراسة وظيفة FDFT1. وتم إنشاء سطر فاصل شرطي للفأرة يسمى Fdft1tm1a (KOMP) Wtsi في معهد ويلكوم ترست سانجر .[34] وخضعت الحيوانات الذكور والإناث لشاشة ظاهرية قياسية [35] لتحديد آثار الحذف.[36][37][38][39] الشاشات الإضافية التي تم القيام بها: - التنميط المناعي العميق [40]
| Characteristic | Phenotype |
|---|---|
| All data available at.[35][40] | |
| علم الدم 6 Weeks | Normal |
| Insulin | Normal |
| Homozygous viability at P14 | Abnormal |
| سيادة (وراثة) lethal study | Abnormal |
| Body weight | Normal |
| Neurological assessment | Normal |
| Grip strength | Normal |
| علم المسوخ | Normal |
| Indirect calorimetry | Normal |
| اختبار تحمل الجلوكوز | Normal |
| استجابة جذع الدماغ السمعية | Normal |
| DEXA | Normal |
| تصوير شعاعي | Normal |
| Eye morphology | Normal |
| كيمياء سريرية | Normal |
| علم الدم 16 Weeks | Normal |
| Peripheral blood leukocytes 16 Weeks | Normal |
| Heart weight | Normal |
| سلمونيلا infection | Normal |
| Cytotoxic T Cell Function | Normal |
| Spleen Immunophenotyping | Normal |
| Mesenteric Lymph Node Immunophenotyping | Normal |
| Bone Marrow Immunophenotyping | Normal |
| Epidermal Immune Composition | Normal |
| Trichuris Challenge | Normal |
المراجع
- Ichikawa M, Yokomizo A, Itoh M, Sugita K, Usui H, Shimizu H, Suzuki M, Terayama K, Kanda A (مارس 2011)، "Discovery of a new 2-aminobenzhydrol template for highly potent squalene synthase inhibitors"، Bioorg. Med. Chem.، 19 (6): 1930–49، doi:10.1016/j.bmc.2011.01.065، PMID 21353782.
- Tansey TR, Shechter I (ديسمبر 2000)، "Structure and regulation of mammalian squalene synthase"، Biochim. Biophys. Acta، 1529 (1–3): 49–62، doi:10.1016/S1388-1981(00)00137-2، PMID 11111077.
- Nakashima T, Inoue T, Oka A, Nishino T, Osumi T, Hata S (مارس 1995)، "Cloning, expression, and characterization of cDNAs encoding Arabidopsis thaliana squalene synthase"، Proc. Natl. Acad. Sci. U.S.A.، 92 (6): 2328–32، Bibcode:1995PNAS...92.2328N، doi:10.1073/pnas.92.6.2328، PMC 42476، PMID 7892265.
- Tansey TR, Shechter I (2001)، "Squalene synthase: structure and regulation"، Prog. Nucleic Acid Res. Mol. Biol.، Progress in Nucleic Acid Research and Molecular Biology، 65: 157–95، doi:10.1016/S0079-6603(00)65005-5، ISBN 9780125400657، PMID 11008488.
- Pandit J, Danley DE, Schulte GK, Mazzalupo S, Pauly TA, Hayward CM, Hamanaka ES, Thompson JF, Harwood HJ (سبتمبر 2000)، "Crystal structure of human squalene synthase. A key enzyme in cholesterol biosynthesis"، J. Biol. Chem.، 275 (39): 30610–7، doi:10.1074/jbc.M004132200، PMID 10896663.
- Jennings SM, Tsay YH, Fisch TM, Robinson GW (يوليو 1991)، "Molecular cloning and characterization of the yeast gene for squalene synthetase"، Proc. Natl. Acad. Sci. U.S.A.، 88 (14): 6038–42، Bibcode:1991PNAS...88.6038J، doi:10.1073/pnas.88.14.6038، PMC 52017، PMID 2068081.
- Gu P, Ishii Y, Spencer TA, Shechter I (مايو 1998)، "Function-structure studies and identification of three enzyme domains involved in the catalytic activity in rat hepatic squalene synthase"، J. Biol. Chem.، 273 (20): 12515–25، doi:10.1074/jbc.273.20.12515، PMID 9575210.
- Poulter CD (1990)، "Biosynthesis of non-head-to-tail terpenes. Formation of 1'-1 and 1'-3 linkages"، Accounts of Chemical Research، 23 (3): 70–77، doi:10.1021/ar00171a003.
- Lin FY, Liu CI, Liu YL, Zhang Y, Wang K, Jeng WY, Ko TP, Cao R, Wang AH, Oldfield E (ديسمبر 2010)، "Mechanism of action and inhibition of dehydrosqualene synthase"، Proc. Natl. Acad. Sci. U.S.A.، 107 (50): 21337–42، Bibcode:2010PNAS..10721337L، doi:10.1073/pnas.1010907107، PMC 3003041، PMID 21098670.
- Beytia E, Qureshi AA, Porter JW (مارس 1973)، "Squalene synthetase. 3. Mechanism of the reaction"، J. Biol. Chem.، 248 (5): 1856–67، PMID 4348553.
- Mookhtiar KA, Kalinowski SS, Zhang D, Poulter CD (أبريل 1994)، "Yeast squalene synthase. A mechanism for addition of substrates and activation by NADPH"، J. Biol. Chem.، 269 (15): 11201–7، PMID 8157649.
- Blagg, Brian S. J.؛ Jarstfer, Michael B.؛ Rogers, Daniel H.؛ Poulter, C. Dale (04 يوليو 2002)، "Recombinant Squalene Synthase. A Mechanism for the Rearrangement of Presqualene Diphosphate to Squalene"، Journal of the American Chemical Society (باللغة الإنجليزية)، 124 (30): 8846–8853، doi:10.1021/ja020411a، مؤرشف من الأصل في 10 يناير 2020.
- Jarstfer, Michael B.؛ Blagg, Brian S. J.؛ Rogers, Daniel H.؛ Poulter, C. Dale (25 ديسمبر 1996)، "Biosynthesis of Squalene. Evidence for a Tertiary Cyclopropylcarbinyl Cationic Intermediate in the Rearrangement of Presqualene Diphosphate to Squalene"، Journal of the American Chemical Society (باللغة الإنجليزية)، 118 (51): 13089–13090، doi:10.1021/ja963308s، مؤرشف من الأصل في 10 يناير 2020.
- Brown, Michael S.؛ Goldstein, Joseph L. (1980)، "Multivalent feedback regulation of HMG CoA reductase, a control mechanism coordinating isoprenoid synthesis and cell growth"، J. Lipid Res.، 21 (5): 505–517، PMID 6995544.
- Olson, Robert E. (01 يناير 1967)، Robert S. Harris, Ira G. Wool, John A. Loraine, G. F. Marrian and Kenneth V. Thimann (المحرر)، Biosynthesis of Ubiquinones in Animals*، Academic Press، ج. 24، ص. 551–574، doi:10.1016/s0083-6729(08)60221-6، مؤرشف من الأصل في 10 يناير 2020.
- Gough, Doreen P.؛ Hemming, F. W. (01 يونيو 1970)، "The characterization and stereochemistry of biosynthesis of dolichols in rat liver"، Biochemical Journal، 118 (1): 163–166، doi:10.1042/bj1180163، ISSN 0264-6021، PMC 1179092، PMID 4319540.
- Faust, Jerry R.؛ Goldstein, Joseph L.؛ Brown, Michael S. (01 يناير 1979)، "Synthesis of ubiquinone and cholesterol in human fibroblasts: Regulation of a branched pathway"، Archives of Biochemistry and Biophysics، 192 (1): 86–99، doi:10.1016/0003-9861(79)90074-2، مؤرشف من الأصل في 24 فبراير 2020.
- Faust, Jerry R.؛ Goldstein, Joseph L.؛ Brown, Michael S. (01 أكتوبر 1979)، "Squalene synthetase activity in human fibroblasts: Regulation via the low density lipoprotein receptor"، Proceedings of the National Academy of Sciences of the United States of America، 76 (10): 5018–5022، Bibcode:1979PNAS...76.5018F، doi:10.1073/pnas.76.10.5018، ISSN 0027-8424، PMC 413070، PMID 228272.
- Guan, G.؛ Jiang, G.؛ Koch, R. L.؛ Shechter, I. (15 سبتمبر 1995)، "Molecular cloning and functional analysis of the promoter of the human squalene synthase gene"، The Journal of Biological Chemistry، 270 (37): 21958–21965، doi:10.1074/jbc.270.37.21958، ISSN 0021-9258، PMID 7665618.
- Guan, Guimin؛ Dai, Pei-Hua؛ Osborne, Timothy F.؛ Kim, Jae B.؛ Shechter, Ishaiahu (11 أبريل 1997)، "Multiple Sequence Elements are Involved in the Transcriptional Regulation of the Human Squalene Synthase Gene"، Journal of Biological Chemistry (باللغة الإنجليزية)، 272 (15): 10295–10302، doi:10.1074/jbc.272.15.10295، ISSN 0021-9258، PMID 9092581، مؤرشف من الأصل في 31 يوليو 2019.
- Guan, G.؛ Dai, P.؛ Shechter, I. (15 مايو 1998)، "Differential transcriptional regulation of the human squalene synthase gene by sterol regulatory element-binding proteins (SREBP) 1a and 2 and involvement of 5' DNA sequence elements in the regulation"، The Journal of Biological Chemistry، 273 (20): 12526–12535، doi:10.1074/jbc.273.20.12526، ISSN 0021-9258، PMID 9575211.
- Kourounakis AP, Katselou MG, Matralis AN, Ladopoulou EM, Bavavea E (2011)، "Squalene synthase inhibitors: An update on the search for new antihyperlipidemic and antiatherosclerotic agents"، Curr. Med. Chem.، 18 (29): 4418–39، doi:10.2174/092986711797287557، PMID 21864285.
- Paradise EM, Kirby J, Chan R, Keasling JD (يونيو 2008)، "Redirection of flux through the FPP branch-point in Saccharomyces cerevisiae by down-regulating squalene synthase"، Biotechnol. Bioeng.، 100 (2): 371–8، doi:10.1002/bit.21766، PMID 18175359.
- Okazaki H, Tazoe F, Okazaki S, Isoo N, Tsukamoto K, Sekiya M, Yahagi N, Iizuka Y, Ohashi K, Kitamine T, Tozawa R, Inaba T, Yagyu H, Okazaki M, Shimano H, Shibata N, Arai H, Nagai RZ, Kadowaki T, Osuga J, Ishibashi S (سبتمبر 2006)، "Increased cholesterol biosynthesis and hypercholesterolemia in mice overexpressing squalene synthase in the liver"، J. Lipid Res.، 47 (9): 1950–8، doi:10.1194/jlr.M600224-JLR200، PMID 16741291.
- Davidson MH (يناير 2007)، "Squalene synthase inhibition: a novel target for the management of dyslipidemia"، Curr Atheroscler Rep، 9 (1): 78–80، doi:10.1007/BF02693932، PMID 17169251.
- Do R, Kiss RS, Gaudet D, Engert JC (يناير 2009)، "Squalene synthase: a critical enzyme in the cholesterol biosynthesis pathway"، Clin. Genet.، 75 (1): 19–29، doi:10.1111/j.1399-0004.2008.01099.x، PMID 19054015.
- Hiyoshi H, Yanagimachi M, Ito M, Saeki T, Yoshida I, Okada T, Ikuta H, Shinmyo D, Tanaka K, Kurusu N, Tanaka H (نوفمبر 2001)، "Squalene synthase inhibitors reduce plasma triglyceride through a low-density lipoprotein receptor-independent mechanism"، Eur. J. Pharmacol.، 431 (3): 345–52، doi:10.1016/S0014-2999(01)01450-9، PMID 11730728.
- Seiki S, Frishman WH (2009)، "Pharmacologic inhibition of squalene synthase and other downstream enzymes of the cholesterol synthesis pathway: a new therapeutic approach to treatment of hypercholesterolemia"، Cardiol Rev، 17 (2): 70–6، doi:10.1097/CRD.0b013e3181885905، PMID 19367148.
- Charlton-Menys V, Durrington PN (2007)، "Squalene synthase inhibitors : clinical pharmacology and cholesterol-lowering potential"، Drugs، 67 (1): 11–6، doi:10.2165/00003495-200767010-00002، PMID 17209661.
- Amin D, Rutledge RZ, Needle SN, Galczenski HF, Neuenschwander K, Scotese AC, Maguire MP, Bush RC, Hele DJ, Bilder GE, Perrone MH (مايو 1997)، "RPR 107393, a potent squalene synthase inhibitor and orally effective cholesterol-lowering agent: comparison with inhibitors of HMG-CoA reductase"، J. Pharmacol. Exp. Ther.، 281 (2): 746–52، PMID 9152381.
- Gibbs, Edwina (29 أكتوبر 2007)، "UPDATE 2-US FDA tells Takeda to stop some TAK-475 trials"، Reuters، مؤرشف من الأصل في 24 سبتمبر 2015، اطلع عليه بتاريخ 05 مارس 2013.
- "Discontinuation of Development of TAK-475, A Compound for Treatment of Hypercholesterolemia"، Takeda Pharmaceutical Company Limited، 28 مارس 2008، مؤرشف من الأصل في 24 سبتمبر 2015، اطلع عليه بتاريخ 05 مارس 2013.
- Liu CI, Liu GY, Song Y, Yin F, Hensler ME, Jeng WY, Nizet V, Wang AH, Oldfield E (مارس 2008)، "A cholesterol biosynthesis inhibitor blocks Staphylococcus aureus virulence"، Science، 319 (5868): 1391–4، Bibcode:2008Sci...319.1391L، doi:10.1126/science.1153018، PMC 2747771، PMID 18276850.
- Gerdin AK (2010)، "The Sanger Mouse Genetics Programme: high throughput characterisation of knockout mice"، Acta Ophthalmologica، 88: 925–7، doi:10.1111/j.1755-3768.2010.4142.x.
- "International Mouse Phenotyping Consortium"، مؤرشف من الأصل في 30 نوفمبر 2018.
- Skarnes WC, Rosen B, West AP, Koutsourakis M, Bushell W, Iyer V, Mujica AO, Thomas M, Harrow J, Cox T, Jackson D, Severin J, Biggs P, Fu J, Nefedov M, de Jong PJ, Stewart AF, Bradley A (يونيو 2011)، "A conditional knockout resource for the genome-wide study of mouse gene function"، Nature، 474 (7351): 337–42، doi:10.1038/nature10163، PMC 3572410، PMID 21677750.
- Dolgin E (يونيو 2011)، "Mouse library set to be knockout"، Nature، 474 (7351): 262–3، doi:10.1038/474262a، PMID 21677718.
- Collins FS, Rossant J, Wurst W (يناير 2007)، "A mouse for all reasons"، Cell، 128 (1): 9–13، doi:10.1016/j.cell.2006.12.018، PMID 17218247.
- White JK, Gerdin AK, Karp NA, Ryder E, Buljan M, Bussell JN, Salisbury J, Clare S, Ingham NJ, Podrini C, Houghton R, Estabel J, Bottomley JR, Melvin DG, Sunter D, Adams NC, Sanger Institute Mouse Genetics Project, Tannahill D, Logan DW, Macarthur DG, Flint J, Mahajan VB, Tsang SH, Smyth I, Watt FM, Skarnes WC, Dougan G, Adams DJ, Ramirez-Solis R, Bradley A, Steel KP (2013)، "Genome-wide generation and systematic phenotyping of knockout mice reveals new roles for many genes"، Cell، 154 (2): 452–64، doi:10.1016/j.cell.2013.06.022، PMC 3717207، PMID 23870131.
- "Infection and Immunity Immunophenotyping (3i) Consortium"، مؤرشف من الأصل في 24 فبراير 2020.
روابط خارجية
- Farnesyl-Diphosphate+Farnesyltransferase في المكتبة الوطنية الأمريكية للطب نظام فهرسة المواضيع الطبية (MeSH).
 بوابة الكيمياء
بوابة الكيمياء بوابة الكيمياء الحيوية
بوابة الكيمياء الحيوية