فوسفوينوسيتيد 3 كيناز
فوسفينوسيتايد 3-كينازات (PI3Ks)، وتسمى أيضًا الفوسفاتيدلينوسيتول 3 كيناز ، هي عائلة من الإنزيمات المشاركة في الوظائف الخلوية مثل نمو الخلايا، والتكاثر، والتمايز، والحركة، والبقاء، والاتجار داخل الخلايا، والتي بدورها تشارك في السرطان.
| Phosphatidylinositol-4,5-bisphosphate 3-kinase | |
|---|---|
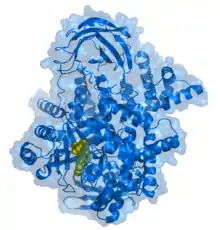 | |
| PIK-93 inhibitor (yellow) bound to the PI3K 110 gamma subunit .[1] | |
| معرف | |
| رمز | PI3K |
| قاعدة بيانات عوائل البروتينات | PF00454 |
| إنتربرو | IPR000403 |
| SMART | SM00146 |
| بروسايت | PDOC00710 |
| قاعدة بيانات التصنيف الهيكلي للبروتينات | 3gmm |
| قاعدة بيانات توجهات البروتينات في الأغشية | 265 |
| قاعدة بيانات توجهات البروتينات في الأغشية | 3ml9 |
PI3Ks أسرة مكونة من الخلايا ذات الصلة إشارة محول الانزيمات قادرة على الفسفرة 3 موقف الهيدروكسيل مجموعة من إينوزيتول حلقة من فوسفتيدلينوستول (PtdIns). [2] المسار، مع الجين الورمي PIK3CA والقامع الورم PTEN ، وتورط في حساسية الأورام السرطانية الأنسولين و IGF1 ، و الحد من السعرات الحرارية . [3] [4]
اكتشاف
اكتشاف PI3Ks التي كتبها لويس كانتلي بدأ وزملاؤه مع التعرف عليهم من فسفوإينوزيتيد لم تكن معروفة سابقا كيناز المرتبطة التورام البروتين الأوسط T. [5] لاحظوا خصوصية الركيزة الفريدة وخصائص الكروماتوجرافي لمنتجات كيناز الدهون، مما أدى إلى اكتشاف أن كيناز الفوسفوينوزيدات هذا لديه قدرة غير مسبوقة على فسفوريلات الفوسفوينوزيدات على موضع 3 'من حلقة الإينوزيتول. [6] في وقت لاحق، أظهر كانتلي وزملاؤه أنه في الجسم الحي يفضل الإنزيم PtdIns (4,5) P2 كركيزة، مما ينتج عن فسفوينوزيتيد PtdIns (3,4,5) P3 [7] تم تحديدها مسبقًا في العدلات [8]
الطبقات
تنقسم عائلة PI3K إلى أربع فئات مختلفة: الفئة الأولى والفئة الثانية والفئة الثالثة والفئة الرابعة. وتستند التصنيفات على البنية الأساسية والتنظيم وخصوصية الركيزة في المختبر . [9]
الفئة الأولى
تعمل الفئة I PI3Ks على تحفيز تحويل فوسفاتيديلنوسيتول (4,5) - ثنائي فوسفات (PI (4,5) P 2 ) إلى فوسفاتيديل لينوسيتول ( 3,4,5) - ثلاثي فوسفات (PI (3,4,5) P 3 ) في الجسم الحي . أثناء وجودها في المختبر، تم أيضًا إثبات أنها تعمل على تحويل فوسفاتيدنلوسيتول (PI) إلى فوسفاتيدنوسيتول 3-فوسفات (PI3P) وفسفاتيديللينوسيتول 4-فوسفات (PI4P) إلى فوسفاتيدنلوسيتول (3,4) -فوسفات (PI (3,4) P 2 ) ، يتم رد فعل هذه ردود الفعل بشدة في الجسم الحي. [10] [11] [12] [13] يتم تنشيط PI3K بواسطة مستقبلات G المقترنة بالبروتين ومستقبلات كيناز التيروزين . [9]
الصف الأول PI3Ks هي غير متماثل جزيئات تتكون من تنظيمية وتحفيزية فرعية . يتم تقسيمها كذلك بين مجموعات فرعية IA و IB على تشابه التسلسل. تتكون الفئة IA PI3Ks من مثاقب غير متماثل بين وحدة فرعية حفازة p110 ووحدة فرعية تنظيمية p85. [14] هناك خمسة أنواع من الوحدة الفرعية التنظيمية p85 ، وهي p85α المعينة، و p55α ، و p50α ، و p85β ، و p55γ . هناك أيضًا ثلاثة أنواع من الوحدة الفرعية الحفازة p110 المعينة p110α أو β أو un الوحدة التحفيزية الفرعية. الوحدات الفرعية التنظيمية الثلاثة الأولى كلها متغيرات لصق من نفس الجين ( Pik3r1 ) ، ويتم التعبير عن الاثنين الآخرين بواسطة جينات أخرى (Pik3r2 و Pik3r3 و p85β و p55γ ، على التوالي). الوحدة الفرعية التنظيمية الأكثر وضوحًا هي p85α ؛ يتم التعبير عن جميع الوحدات الفرعية التحفيزية الثلاثة بواسطة جينات منفصلة ( Pik3ca و Pik3cb و Pik3cd لـ p110α و p110β و p110δ ، على التوالي). يتم التعبير عن أول شكلين متساويين p110 (α و β) في جميع الخلايا، ولكن يتم التعبير عن p110δ في المقام الأول في الكريات البيض ، وقد اقترح أنه تطور بالتوازي مع نظام المناعة التكيفي. تشتمل الوحدات الفرعية p101 التنظيمية والوحدات الحفازة p110γ على الفئة IB PI3Ks ويتم تشفيرها بواسطة جين واحد لكل منها ( Pik3cg لـ p110γ و Pik3r5 لـ p101 ).
تحتوي الوحدات الفرعية p85 على مجالات SH2 و SH3 ( الوراثة المندلية البشرية عبر الإنترنت (OMIM): 171833). ترتبط نطاقات SH2 بشكل تفضيلي ببقايا التيروزين الفسفوريلات في سياق تسلسل الأحماض الأمينية YXXM. [15] [16]
الفئتان الثانية والثالثة
يتم تمييز الصنف الثاني والثالث من PI3Ks عن الصنف الأول من خلال هيكلها ووظيفتها. السمة المميزة للفئة الثانية PI3Ks هي مجال C- المحطة C2. يفتقر هذا المجال إلى بقايا Asp الحرجة لتنسيق ربط Ca 2+ ، مما يشير إلى أن الفئة II PI3Ks تربط الدهون بطريقة Ca 2+ - بشكل مستقل.
تشتمل الفئة الثانية على ثلاثة أشكال متوازنة حفازة (C2α و C2β و C2γ) ، ولكن على عكس الفئتين الأولى والثالثة، لا توجد بروتينات تنظيمية. تحفز الفئة الثانية إنتاج PI (3) P من PI و PI (3,4) P 2 من PI (4) P ؛ ومع ذلك، لا يُعرف سوى القليل عن دورها في الخلايا المناعية. ومع ذلك، فقد ثبت أن PI (3,4) P 2 يلعب دورًا في مرحلة الغزو من كثرة الكريات الخلوية بوساطة الكاثرين. [17] يتم التعبير عن C2α و C2β من خلال الجسم، ولكن التعبير عن C2γ يقتصر على خلايا الكبد .
تنتج الفئة الثالثة PI3Ks فقط PI (3) P من PI [9] ولكنها أكثر تشابهًا مع الفئة الأولى في الهيكل، حيث أنها موجودة كمقومات غير متغيرة لمحفزات فرعية ( Vps34 ) ووحدات تنظيمية فرعية (Vps15 / p150). يبدو أن الفئة الثالثة متورطة في المقام الأول في الاتجار بالبروتينات والحويصلات. ومع ذلك، هناك أدلة تظهر أنها قادرة على المساهمة في فعالية العديد من العمليات المهمة للخلايا المناعية، وليس أقلها البلعمة .
الفئة الرابعة
يشار أحيانًا إلى مجموعة من الإنزيمات ذات الصلة البعيدة بالفئة IV PI3Ks. وهو يتألف من ترنح توسع الشعريات المتحور (ATM) ، ترنح توسع الشعريات و Rad3 (ATR) ، كيناز البروتين المعتمد على DNA (DNA-PK) والهدف من الثدييات من الرابايميسين (mTOR). هم كيناز بروتين سيرين / ثريونين.
الجينات البشرية
| مجموعة | الجين | بروتين | أسماء مستعارة | رقم المفوضية الأوروبية |
|---|---|---|---|---|
| حفاز الفئة 1 | PIK3CA | PI3K ، حفاز، ببتيد ألفا | p110-α | 2.7.1.153 |
| PIK3CB | PI3K ، حفاز، ببتيد بيتا | ص 110 - β | ||
| PIK3CG | PI3K ، حفاز، ببتيد غاما | ص 110 - γ | ||
| PIK3CD | PI3K ، حفاز، ببتيد دلتا | ص 110 - δ | ||
| الفئة 1 التنظيمية | PIK3R1 | PI3K ، الوحدة الفرعية التنظيمية 1 (ألفا) | ص 85-α | لا يوجد |
| PIK3R2 | PI3K ، الوحدة الفرعية التنظيمية 2 (تجريبي) | ص 85-β | ||
| PIK3R3 | PI3K ، الوحدة التنظيمية الفرعية 3 (جاما) | ص 55 γ | ||
| PIK3R4 | PI3K ، الوحدة الفرعية التنظيمية 4 | ص 150 | ||
| PIK3R5 | PI3K ، الوحدة الفرعية التنظيمية 5 | ص 101 | ||
| PIK3R6 | PI3K ، الوحدة الفرعية التنظيمية 6 | ص 87 | ||
| الصف 2 | PIK3C2A | PI3K ، الفئة 2 ، ألفا ببتيد ألفا | PI3K-C2α | 2.7.1.154 |
| PIK3C2B | PI3K ، الفئة 2 ، عديد ببتيد بيتا | PI3K-C2β | ||
| PIK3C2G | PI3K ، الفئة 2 ، ببتيد غاما | PI3K-C2γ | ||
| الفئة 3 | PIK3C3 | PI3K ، الفئة 3 | Vps34 | 2.7.1.137 |
الآلية
تعمل الفوسفاتوينوريدات الثلاثية الفسفورية المختلفة التي تنتجها PI3Ks ( PtdIns3P و PtdIns (3,4) P2 و PtdIns (3,5) P2 و PtdIns (3,4,5) P3 ) في آلية يتم من خلالها مجموعة متنوعة من بروتينات الإشارات، التي تحتوي على نطاقات PX ، ومجالات تجانس البروتين ( نطاقات PH) ، ومجالات FYVE أو نطاقات ربط الفسفوينوزيتيد الأخرى، يتم تجنيدها في أغشية خلوية مختلفة.
الوظيفة
تم ربط PI3Ks بمجموعة متنوعة بشكل غير عادي من الوظائف الخلوية، بما في ذلك نمو الخلايا وانتشارها والتمايز والحركة والبقاء والاتجار داخل الخلايا. ترتبط العديد من هذه الوظائف بقدرة الفئة I PI3Ks على تنشيط بروتين كيناز B (PKB ، المعروف أيضًا باسم Akt) كما هو الحال في مسار PI3K / AKT / mTOR . تنظم الأشكال المتجانسة p110 و p110 جوانب مختلفة من الاستجابات المناعية. PI3Ks هي أيضًا عنصر رئيسي في مسار إشارات الأنسولين . وبالتالي هناك اهتمام كبير بدور إشارات PI3K في داء السكري .
الآلية
يرتبط مجال تجانس البروتين الخاص بـ AKT مباشرة بـ PtdIns (3,4,5) P3 و PtdIns (3,4) P2 ، والتي يتم إنتاجها بواسطة PI3Ks المنشط. [18] نظرًا لأن PtdIns (3,4,5) P3 و PtdIns (3,4) P2 مقيدة بغشاء البلازما، فإن هذا يؤدي إلى نقل AKT إلى غشاء البلازما. وبالمثل، فإن كيناز -1 الذي يعتمد على الفسفوينوسيتيد (PDK1 أو، نادرًا ما يشار إليه باسم PDPK1) يحتوي أيضًا على مجال تجانس pleckstrin يرتبط مباشرة بـ PtdIns (3,4,5) P3 و PtdIns (3,4) P2 ، مما يتسبب في ينتقل أيضًا إلى غشاء البلازما عند تنشيط PI3K. يسمح تفاعل PDK1 المنشط و AKT بأن تصبح AKT فسفرة بواسطة PDK1 على ثريونين 308 ، مما يؤدي إلى التنشيط الجزئي لـ AKT. يحدث التنشيط الكامل لـ AKT عند فسفرة سيرين 473 بواسطة مركب TORC2 من بروتين كيناز mTOR .
ثبت أن مسار PI3K / AKT مطلوب لمجموعة متنوعة للغاية من الأنشطة الخلوية - أبرزها الانتشار الخلوي والبقاء. على سبيل المثال، ثبت أنها تشارك في حماية الخلايا النجمية من موت الخلايا المبرمج الناجم عن السيراميد. [19]
تم تحديد العديد من البروتينات الأخرى التي يتم تنظيمها بواسطة \ (3,4,5) P3 ، بما في ذلك بروتون التيروزين كيناز والمستقبل العام لـ فوسفونيستيديس-1 (GRP1) و الجين OQT المرتبط بـ O-GlcNAc ) ترانسفيراز .
PtdIns (3,4,5) P3 ينشط أيضًا عوامل تبادل النوكليوتيدات الجينية (GEFs) التي تنشط GTPase Rac1 [20] ، مما يؤدي إلى بلمرة الأكتين وإعادة ترتيب الخلايا الهيكلية.[21]
السرطان
يتم تحور فئة IA PI3K p110α في العديد من السرطانات. تتسبب العديد من هذه الطفرات في زيادة نشاط الكيناز. وهو الكيناز الوحيد الأكثر تحولا في ورم أرومي دبقي، وهو أكثر أورام الدماغ الأولية خبيثة. [22] PtdIns (3,4,5) P 3 فوسفاتاز PTEN الذي يعارض إشارات PI3K غائب عن العديد من الأورام. بالإضافة إلى ذلك، يتم تنشيط مستقبلات عامل نمو البشرة EGFR الذي يعمل في الجزء العلوي من PI3K بشكل طافري أو يتم الإفراط في التعبير عنه في السرطان. [23] وبالتالي، يساهم نشاط PI3K بشكل كبير في التحول الخلوي وتطور السرطان .
التعلم والذاكرة
PI3Ks متورطة أيضًا في التقوية طويلة الأجل (LTP). ما إذا كانت مطلوبة للتعبير أو تحريض LTP لا تزال موضع جدل. في الخلايا العصبية للفأر CA1 الحصين، يتم تعقيد بعض PI3Ks مع مستقبلات AMPA ومجزأة في كثافة ما بعد المشابك من المشابك الجلوتاميترجية. [24] يتم فسفرة PI3Ks على نشاط CaMKII المعتمد على مستقبلات NMDA ، [25] ثم يسهل إدخال الوحدات الفرعية AMPA-R GluR1 في غشاء البلازما. هذا يشير إلى أن PI3Ks مطلوبة للتعبير عن LTP. علاوة على ذلك، ألغت مثبطات PI3K تعبير LTP في الحصين CA1 الفئران، لكنها لا تؤثر على تحريضه. [26] والجدير بالذكر أن اعتماد تعبير LTP في المرحلة المتأخرة على PI3Ks يبدو أنه يتناقص بمرور الوقت. [27]
ومع ذلك، وجدت دراسة أخرى أن مثبطات PI3K قمعت الحث، ولكن ليس التعبير، من LTP في الحصين CA1 الماوس. [28] يجند مسار PI3K أيضًا العديد من البروتينات الأخرى في اتجاه التيار، بما في ذلك mTOR ، [29] GSK3β ، [30] و PSD-95 . يؤدي مسار PI3K-mTOR إلى فسفرة p70S6K ، كيناز يسهل النشاط الانتقالي، [31] [32] مما يشير أيضًا إلى أن PI3Ks مطلوبة لمرحلة تخليق البروتين لتحريض LTP بدلاً من ذلك.
تتفاعل PI3Ks مع ركيزة مستقبلات الأنسولين (IRS) لتنظيم امتصاص الجلوكوز من خلال سلسلة من أحداث الفسفرة.
PI 3 كيناز ك كيناز البروتين
يبدو أن العديد من PI3Ks لها نشاط كيناز سيريني / ثريونين في المختبر ؛ ومع ذلك، من غير الواضح ما إذا كان لهذا أي دور في الجسم الحي . [بحاجة لمصدر]
تثبيط
يتم تثبيط جميع PI3Ks عن طريق الأدوية الورتمانين و LY294002 ، على الرغم من أن بعض أعضاء عائلة الفئة PI3K يظهرون حساسية منخفضة. يظهر الورتمانين كفاءة أفضل من LY294002 في مواضع طفرة النقاط الساخنة (GLU542 و GLU545 و HIS1047) [33] [34]
مثبطات PI3K كعلاجات
بما أن الورتمانين و LY294002 مثبطات واسعة النطاق لـ PI3Ks وعدد من البروتينات غير ذات الصلة بتركيزات أعلى، فهي سامة جدًا بحيث لا يمكن استخدامها كعلاجات. وهكذا طور عدد من الشركات الصيدلانية مثبطات PI3K الخاصة بالإيسوفورم. اعتبارًا من يناير 2019 ، تمت الموافقة على ثلاثة مثبطات PI3K من قبل FDA للاستخدام السريري الروتيني في البشر: مثبط PIK3CD أيديليسب (يوليو 2014 ، NDA 206545 ) ، و كوبانليسيب المزدوج PIK3CA و PIK3CD (سبتمبر 2017 ، NDA 209936 ) ، والمزدوج PIK3CD وPIK3CG المانع ديوفيليسب (سبتمبر 2018، NDA 211155 ).
انظر أيضا
- مسار PI3K / AKT / mTOR
المراجع
- ببب: 2chz; Knight ZA, Gonzalez B, Feldman ME, Zunder ER, Goldenberg DD, Williams O, وآخرون (مايو 2006)، "A pharmacological map of the PI3-K family defines a role for p110alpha in insulin signaling"، Cell، 125 (4): 733–47، doi:10.1016/j.cell.2006.03.035، PMC 2946820، PMID 16647110.
- myo-inositol نسخة محفوظة 2020-04-04 على موقع واي باك مشين.
- Giese N (2009)، "Cell pathway on overdrive prevents cancer response to dietary restriction"، PhysOrg.com، مؤرشف من الأصل في 04 أبريل 2020، اطلع عليه بتاريخ 22 أبريل 2009.
- "Tumours with PI3K activation are resistant to dietary restriction"، Nature، 458 (7239): 725–31، أبريل 2009، Bibcode:2009Natur.458..725K، doi:10.1038/nature07782، PMID 19279572.
- "Association of phosphatidylinositol kinase activity with polyoma middle-T competent for transformation"، Nature، 315 (6016): 239–42، 1985، Bibcode:1985Natur.315..239W، doi:10.1038/315239a0، PMID 2987699.
- "Type I phosphatidylinositol kinase makes a novel inositol phospholipid, phosphatidylinositol-3-phosphate"، Nature، 332 (6165): 644–6، أبريل 1988، Bibcode:1988Natur.332..644W، doi:10.1038/332644a0، PMID 2833705.
- "PDGF-dependent tyrosine phosphorylation stimulates production of novel polyphosphoinositides in intact cells"، Cell، 57 (1): 167–75، أبريل 1989، doi:10.1016/0092-8674(89)90182-7، PMID 2467744.
- "An inositol tetrakisphosphate-containing phospholipid in activated neutrophils"، Nature، 334 (6180): 353–6، يوليو 1988، Bibcode:1988Natur.334..353T، doi:10.1038/334353a0، PMID 3393226.
- "Signalling through phosphoinositide 3-kinases: the lipids take centre stage"، Current Opinion in Cell Biology، 11 (2): 219–25، أبريل 1999، doi:10.1016/S0955-0674(99)80029-5، PMID 10209156.
- "The PI3K Pathway in Human Disease"، Cell، 170 (4): 605–635، أغسطس 2017، doi:10.1016/j.cell.2017.07.029، PMID 28802037.
- "Classes of phosphoinositide 3-kinases at a glance"، Journal of Cell Science، 127 (Pt 5): 923–8، مارس 2014، doi:10.1242/jcs.093773، PMID 24587488.
- "PI3K signalling: the path to discovery and understanding"، Nature Reviews. Molecular Cell Biology، 13 (3): 195–203، فبراير 2012، doi:10.1038/nrm3290، PMID 22358332.
- "Signaling by the phosphoinositide 3-kinase family in immune cells"، Annual Review of Immunology، 31 (2): 675–704، يناير 2013، doi:10.1146/annurev-immunol-032712-095946، PMID 23330955.
- "Purification and characterization of phosphoinositide 3-kinase from rat liver"، The Journal of Biological Chemistry، 265 (32): 19704–11، نوفمبر 1990، PMID 2174051.
- "SH2 domains recognize specific phosphopeptide sequences"، Cell، 72 (5): 767–78، مارس 1993، doi:10.1016/0092-8674(93)90404-E، PMID 7680959.
{{استشهاد بدورية محكمة}}: الوسيط|إظهار المؤلفين=6غير صالح (مساعدة) - "Genetic analysis of a phosphatidylinositol 3-kinase SH2 domain reveals determinants of specificity"، Molecular and Cellular Biology، 14 (9): 5929–38، سبتمبر 1994، doi:10.1128/MCB.14.9.5929، PMID 8065326.
- "Phosphoinositides in endocytosis"، Biochimica et Biophysica Acta، 1851 (6): 794–804، يونيو 2015، doi:10.1016/j.bbalip.2014.09.014، PMID 25264171.
- "Direct regulation of the Akt proto-oncogene product by phosphatidylinositol-3,4-bisphosphate"، Science، 275 (5300): 665–8، يناير 1997، doi:10.1126/science.275.5300.665، PMID 9005852.
- "Cannabinoids protect astrocytes from ceramide-induced apoptosis through the phosphatidylinositol 3-kinase/protein kinase B pathway"، The Journal of Biological Chemistry، 277 (39): 36527–33، سبتمبر 2002، doi:10.1074/jbc.M205797200، PMID 12133838.
- "Phosphoinositide 3-kinase-dependent activation of Rac"، FEBS Letters، 546 (1): 93–7، يوليو 2003، doi:10.1016/s0014-5793(03)00454-x، PMID 12829242.
- "Rho GTPases: biochemistry and biology"، Annual Review of Cell and Developmental Biology، 21: 247–69، 2005، doi:10.1146/annurev.cellbio.21.020604.150721، PMID 16212495.
- "Mutational profiling of kinases in glioblastoma"، BMC Cancer، 14: 718، سبتمبر 2014، doi:10.1186/1471-2407-14-718، PMID 25256166.
{{استشهاد بدورية محكمة}}: الوسيط|إظهار المؤلفين=6غير صالح (مساعدة) - "Recent advances in the molecular understanding of glioblastoma"، Journal of Neuro-Oncology، 108 (1): 11–27، مايو 2012، doi:10.1007/s11060-011-0793-0، PMID 22270850.
- "Activation of PI3-kinase is required for AMPA receptor insertion during LTP of mEPSCs in cultured hippocampal neurons"، Neuron، 38 (4): 611–24، مايو 2003، doi:10.1016/S0896-6273(03)00228-9، PMID 12765612.
{{استشهاد بدورية محكمة}}: الوسيط|إظهار المؤلفين=6غير صالح (مساعدة) - "Calmodulin activates phosphatidylinositol 3-kinase"، The Journal of Biological Chemistry، 272 (45): 28183–6، نوفمبر 1997، doi:10.1074/jbc.272.45.28183، PMID 9353264.
- "Phosphatidylinositol 3-kinase is required for the expression but not for the induction or the maintenance of long-term potentiation in the hippocampal CA1 region"، The Journal of Neuroscience، 22 (9): 3359–65، مايو 2002، doi:10.1523/JNEUROSCI.22-09-03359.2002، PMID 11978812.
- "Involvement of multiple phosphatidylinositol 3-kinase-dependent pathways in the persistence of late-phase long term potentiation expression"، Neuroscience، 137 (3): 833–41، فبراير 2006، doi:10.1016/j.neuroscience.2005.10.012، PMID 16326012.
- "Phosphatidylinositol 3-kinase regulates the induction of long-term potentiation through extracellular signal-related kinase-independent mechanisms"، The Journal of Neuroscience، 23 (9): 3679–88، مايو 2003، doi:10.1523/JNEUROSCI.23-09-03679.2003، PMID 12736339.
- "Phosphatidylinositol 3-kinase activation is required for stress protocol-induced modification of hippocampal synaptic plasticity"، The Journal of Biological Chemistry، 283 (5): 2631–43، فبراير 2008، doi:10.1074/jbc.M706954200، PMID 18057005.
- "LTP inhibits LTD in the hippocampus via regulation of GSK3beta"، Neuron، 53 (5): 703–17، مارس 2007، doi:10.1016/j.neuron.2007.01.029، PMID 17329210، مؤرشف من الأصل في 13 أبريل 2020.
{{استشهاد بدورية محكمة}}: الوسيط|إظهار المؤلفين=6غير صالح (مساعدة) - "Signalling through the lipid products of phosphoinositide-3-OH kinase"، Nature، 387 (6634): 673–6، يونيو 1997، Bibcode:1997Natur.387..673T، doi:10.1038/42648، PMID 9192891.
- "Time-restricted role for dendritic activation of the mTOR-p70S6K pathway in the induction of late-phase long-term potentiation in the CA1"، Proceedings of the National Academy of Sciences of the United States of America، 100 (24): 14368–73، نوفمبر 2003، Bibcode:2003PNAS..10014368C، doi:10.1073/pnas.2336098100، PMID 14623952.
- Investigating the Inhibitory Effect of Wortmannin in the Hotspot Mutation at Codon 1047 of PIK3CA Kinase Domain: A Molecular Docking and Molecular Dynamics Approach، Advances in Protein Chemistry and Structural Biology، ج. 102، 01 يناير 2016، ص. 267–97، doi:10.1016/bs.apcsb.2015.09.008، ISBN 9780128047958، PMID 26827608.
- "Deciphering the impact of somatic mutations in exon 20 and exon 9 of PIK3CA gene in breast tumors among Indian women through molecular dynamics approach"، Journal of Biomolecular Structure & Dynamics، 34 (1): 29–41، 02 يناير 2016، doi:10.1080/07391102.2015.1007483، PMID 25679319.
قراءة متعمقة
- "Synthesis and function of 3-phosphorylated inositol lipids"، Annual Review of Biochemistry، 70: 535–602، 2001، doi:10.1146/annurev.biochem.70.1.535، PMID 11395417.
{{استشهاد بدورية محكمة}}: الوسيط|إظهار المؤلفين=6غير صالح (مساعدة) - "PI3K signaling maintains c-myc expression to regulate transcription of E2F1 in pancreatic cancer cells"، Molecular Carcinogenesis، 48 (12): 1149–58، ديسمبر 2009، doi:10.1002/mc.20569، PMID 19603422.
- "Form and flexibility in phosphoinositide 3-kinases"، Biochemical Society Transactions، 37 (Pt 4): 615–26، أغسطس 2009، doi:10.1042/BST0370615، PMID 19614567.
- "Regulation of mRNA export by the PI3 kinase/AKT signal transduction pathway"، Molecular Biology of the Cell، 24 (8): 1208–21، أبريل 2013، doi:10.1091/mbc.E12-06-0450، PMC 3623641، PMID 23427269.
روابط خارجية
- Eukaryotic Linear Motif resource Eukaryotic Linear Motif resource
- بروتيوبيديا Phosphoinositide_3-Kinases لاستكشاف هيكل في 3D التفاعلية
- PI-3+Kinase في المكتبة الوطنية الأمريكية للطب نظام فهرسة المواضيع الطبية (MeSH).
 بوابة الكيمياء الحيوية
بوابة الكيمياء الحيوية بوابة طب
بوابة طب بوابة علم الأحياء
بوابة علم الأحياء