مستقبل ن-مثيل-د-أسبارتات
مستقبل ن-مثيل-د-أسبارتات (بالإنجليزية: N-methyl-D-aspartate receptor)، اختصاره «مستقبل إن إم دي إيه» أو «إن إم دي إيه آر»، مستقبل غلوتامات وقناة شاردية توجد في الخلايا العصبية. يمثل هذا المستقبل أحد الأنواع الثلاثة من مستقبلات الغلوتامات المؤثرة في التقلص العضلي، والنوعان الآخران هما «إيه إم بي إيه» ومستقبلات الكاينات.
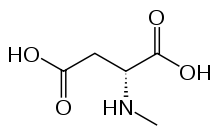
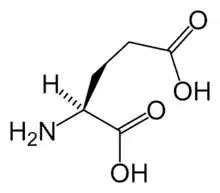
تتشكل الربيطة من الغلوتامات والغليسين (أو دي سيرين) اعتمادًا على مكونات الوحدات الفرعية، ولا يكفي الاتصال بين الربائط عادةً لفتح القنوات الشاردية إذا كانت مغلقةً بشوارد المغنزيوم. تتحرر هذه الشوارد فقط عند إزالة الاستقطاب الكافي للخلية العصبية، وهكذا تؤدي القناة دور «محدد تزامن»، إذ لا تفتح سوى عند اجتماع هذين الشرطين، وبالتالي تسمح للشوارد المشحونة إيجابًا (الكاتيونات) بالانتقال عبر الغشاء الخلوي. من المعتقد أن مستقبل ن-مثيل-د-أسبارتات يؤدي دورًا مهمًا في التحكم باللدونة المشبكية ويتواسط عمل وظائف الذاكرة والتعلم.[1][2]
يعد هذا المركب مستقبلًا أيونوتروبيًا، أي أنه بروتين يسمح بعبور الشوارد عبر الغشاء الخلوي.[3] سمي المستقبل بهذا الاسم لأن الجزيء الناهض ن-مثيل-د-أسبارتات يرتبط انتقائيًا به دونًا عن مستقبلات الغلوتامات الأخرى.
تفتح القنوات الشاردية غير الانتقائية للشوارد الإيجابية نتيجة تنشيط مستقبلات ن-مثيل-د-أسبارتات بمحصلة جهد اعتكاس تقارب 0 ميلي ڤولط. يتحدد فتح وإغلاق القنوات الشاردية باتصال الربيطة في المقام الأول، بينما يعتمد جريان التيار عبرها على الجهد الكهربائي. ترتبط شوارد الزنك والمغنزيوم خارج الخلوية مع مواقع نوعية على المستقبل، مانعةً بذلك مرور بقية الشوارد الإيجابية عبر القناة الشاردية المفتوحة. تزيح إزالة استقطاب الخلية شوارد الزنك والمغنزيوم وتطردها من المسام الغشائية، وهذا يسمح بجريان شوارد الصوديوم والكالسيوم إلى داخل الخلية والبوتاسيوم إلى خارجها اعتمادًا على الجهد الكهربائي. من المعتقد أن تدفق شوارد الكالسيوم عبر مستقبلات ن-مثيل-د-أسبارتات عامل حساس في اللدونة المشبكية بسبب البروتينات التي ترتبط وتتفعل بشوارد الكالسيوم، وهي آلية خلوية للتعلم والذاكرة.[4][5][6][7]
تتأثر فعالية هذا المستقبل بالعديد من الأدوية نفسية المفعول مثل الفينسيكليدين (ب سي ب) والكحول (إيثانول) وديكستروميثورفان (دي إكس إم). يعود التأثير المخدر والمسكن للأدوية مثل الكيتامين وأكسيد النيتروس جزئيًا لتأثيرها على فعالية مستقبل ن-مثيل-د-أسبارتات. يزيد التفعيل المفرط لهذا المستقبل من تراكيز الكالسيوم والزنك في العصارة الخلوية، ما يساهم إلى حد كبير في الموت العصبي. وُجد أن مركبات الكانابينويد تمنع هذه العملية، وذلك يتطلب تدخل بروتين «إتش آي إن تي 1» ليبطل التأثير السمي الناجم عن إنتاج أحادي أكسيد النيتروجين وإطلاق الزنك المتواسط بمستقبل ن-مثيل-د-أسبارتات من خلال تفعيل مستقبل كانابينويد نوع 1 (سي بي 1).[8] يثبط الكانابينويد أيضًا السمية العصبية المحرضة بالميثامفيتامين (الميث) عبر تثبيط التعبير عن مخلّقات أكسيد النتريك وتفعيل الخلايا النجمية، ويبدو أنه ينقص تلف الدماغ الناجم عن الميثامفيتامين من خلال آليات معتمدة على مستقبل كانابينويد نوع 1 وغيرها. يحدث تثبيط الدباق النجمي المحرض بالميثامفيتامين عبر آليات معتمدة على مستقبل كانابينويد نوع 2 الخاص برباعي هيدرو كانابينول.[9] صُنف الميمانتين منذ عام 1989 ضمن المناهضات غير التنافسية لمستقبل ن-مثيل-د-أسبارتات، إذ يدخل قناة المستقبل بعد تفعيلها ليمنع عبور الشوارد.[10][11][12]
يسبب التفعيل المفرط للمستقبل تدفقًا مفرطًا للكالسيوم، وهذا يقود إلى تحفيز زائد يُفترض أنه يشترك في بعض الاضطرابات العصبية التنكسية، لذلك يكون حصر هذه المستقبلات نظريًا مفيدًا في علاج هذه الأمراض.[12][13][14][15] يشارك قصور مستقبلات ن-مثيل-د-أسبارتات (نتيجة عوز الجلوتاثيون أو لأسباب أخرى) في اعتلال اللدونة المشبكية، وقد تحدث تداعيات سلبية أخرى. [16]
تكمن المشكلة الأساسية في استخدام مناهضات مستقبل ن-مثيل-د-أسبارتات للحماية العصبية في أهمية التأثيرات الفسيولوجية للمستقبل لأداء الوظيفة العصبية الطبيعية. تحتاج مناهضات مستقبلات ن-مثيل-د-أسبارتات لحجب التفعيل الزائد دون التداخل مع الوظائف الطبيعية لتكون فعالةً سريريًا، ويمتلك الميمانتين هذه الخاصية.[17]
التاريخ
تلا اكتشاف مستقبلات إن إم دي إيه اصطناع حمض ن-مثيل-د-أسبارتات ودراسته في ستينيات القرن العشرين على يدي جيف واتكنز وزملائه. ثبت تدخل هذه المستقبلات في سبل مشبكية مركزية عديدة في بداية ثمانينيات القرن العشرين.[18][19]
اكتُشفت انتقائية الوحدات الفرعية للمستقبل في بداية التسعينيات، وهذا أدى إلى التعرف على صنف جديد من المكونات التي تثبط الوحدة الفرعية «ن آر 2 بي» انتقائيًا. قادت هذه الاكتشافات إلى انطلاق حملة نشطة في قطاع الصناعات الدوائية.[20] ربط هذا الاستنتاج بين المستقبلات واضطرابات عصبية متنوعة مثل الصرع وداء باركنسون وألزهايمر وداء هانتنغتون وغيرها من الاضطرابات العصبية المركزية.
في عام 2002، اكتشف هيلمر بادنغ ومساعدوه أن النواتج الخلوية لتحريض مستقبلات إن إم دي إيه تعتمد على موقع المستقبل على سطح الخلية العصبية.[21][22] تحفز المستقبلات المتوضعة على المشابك الخلوية التعبير الجيني والأحداث المرتبطة باللدونة والحماية العصبية المكتسبة. تحفز المستقبلات خارج المشبكية إشارات الموت الخلوي: إذ تسبب توقف الانتساخ الخلوي وتعطيل المتقدرات والانحلال البنيوي. يمثل هذا الثلاثي المرضي لإشارات مستقبلات إن إم دي إيه نقطة تحول مشترك في الآلية المرضية لعدة أمراض عصبية تنكسية حادة ومزمنة.[23]
كشف هيلمر بادنغ ومساعدوه الأساس الجزيئي لتأشير المستقبلات خارج المشبكية عام 2020.[24] تشكل المستقبلات خارج الخلوية مع الميلاستاتين 4 (تي آر بّي إم 4) معقد تأشير موت خلوي، بينما تعيق مثبطات الوصل بين المركبين (المعروفة بمثبطات السطح البيني) تشكيل المعقد وتزيل سمية المستقبلات خارج المشبكية.
في عام 1968، اكتُشف صدفةً أن الأمانتيدين يؤدي إلى هجوع ملحوظ في أعراض باركنسون عندما تناولته إحدى النساء المصابات لعلاج الإنفلونزا وأبدى تحسنًا واضحًا في أعراضها.[25] كان هذا الاكتشاف الذي أبلغ عنه سكاواب وآخرون بداية أبحاث الكيمياء الدوائية لمشتقات مركب أدامانتان في سياق علاج الأدواء المؤثرة على الجملة العصبية المركزية. كان الميمانتين، وهو أحد مشتقات الأدامانتان، يُصنع في شركة إيلي ليلي وشركائه عام 1963 بهدف تطوير دواء لانخفاض سكر الدم، لكنه لم يظهر أي فعالية في ذلك. لم تُكتشف الأهمية العلاجية للميمانتين في علاج الأمراض العصبية التنكسية حتى عام 1972، وصُنف الميمانتين عام 1989 ضمن مناهضات مستقبل إن إم دي إيه.
المصادر
- "Subunit arrangement and function in NMDA receptors"، Nature، 438 (7065): 185–192، نوفمبر 2005، Bibcode:2005Natur.438..185F، doi:10.1038/nature04089، PMID 16281028، S2CID 4400777.
- "Memory and the NMDA receptors"، The New England Journal of Medicine، 361 (3): 302–303، يوليو 2009، doi:10.1056/NEJMcibr0902052، PMC 3703758، PMID 19605837.
- "Molecular cloning and characterization of the rat NMDA receptor"، Nature، 354 (6348): 31–37، نوفمبر 1991، Bibcode:1991Natur.354...31M، doi:10.1038/354031a0، PMID 1834949، S2CID 4368947.
- "The glutamate receptor ion channels"، Pharmacological Reviews، 51 (1): 7–61، مارس 1999، PMID 10049997، مؤرشف من الأصل في 27 أكتوبر 2020.
- "Recent development in NMDA receptors"، Chinese Medical Journal، 113 (10): 948–956، أكتوبر 2000، PMID 11775847.
- "NMDA receptor subunits: diversity, development and disease"، Current Opinion in Neurobiology، 11 (3): 327–335، يونيو 2001، doi:10.1016/S0959-4388(00)00215-4، PMID 11399431، S2CID 11929361.
- "NMDA receptor subunits: function and pharmacology"، Current Opinion in Pharmacology، 7 (1): 39–47، فبراير 2007، doi:10.1016/j.coph.2006.08.011، PMID 17088105، مؤرشف من الأصل في 1 سبتمبر 2019.
- "Cannabinoid receptors couple to NMDA receptors to reduce the production of NO and the mobilization of zinc induced by glutamate"، Antioxidants & Redox Signaling، 19 (15): 1766–82، نوفمبر 2013، doi:10.1089/ars.2012.5100، PMC 3837442، PMID 23600761.
- "Δ9-tetrahydrocannabinol prevents methamphetamine-induced neurotoxicity"، PLOS ONE، 9 (5): e98079، 20 مايو 2014، Bibcode:2014PLoSO...998079C، doi:10.1371/journal.pone.0098079، PMC 4028295، PMID 24844285.
{{استشهاد بدورية محكمة}}: الوسيط|إظهار المؤلفين=6غير صالح (مساعدة) - "Mechanism of action of memantine"، Current Opinion in Pharmacology، 6 (1): 61–7، فبراير 2006، doi:10.1016/j.coph.2005.09.007، PMID 16368266.
- "Management of moderate to severe Alzheimer's disease: focus on memantine"، Taiwanese Journal of Obstetrics & Gynecology، 50 (4): 415–23، ديسمبر 2011، doi:10.1016/j.tjog.2011.10.004، PMID 22212311.
- "The chemical biology of clinically tolerated NMDA receptor antagonists"، Journal of Neurochemistry، 97 (6): 1611–26، يونيو 2006، doi:10.1111/j.1471-4159.2006.03991.x، PMID 16805772، S2CID 18376541.
- "NMDA receptor pathways as drug targets"، Nature Neuroscience، 5 (11): 1039–1042، نوفمبر 2002، doi:10.1038/nn936، PMID 12403981، S2CID 41383776.
- "Paradigm shift in neuroprotection by NMDA receptor blockade: memantine and beyond"، Nature Reviews. Drug Discovery، 5 (2): 160–170، فبراير 2006، doi:10.1038/nrd1958، PMID 16424917، S2CID 21379258.
- "NMDA-antagonism (memantine): an alternative pharmacological therapeutic principle in Alzheimer's and vascular dementia"، Current Pharmaceutical Design، 10 (3): 253–259، 01 يناير 2004، doi:10.2174/1381612043386392، PMID 14754385.
- "Synaptic plasticity impairment and hypofunction of NMDA receptors induced by glutathione deficit: relevance to schizophrenia"، Neuroscience، 137 (3): 807–819، فبراير 2006، doi:10.1016/j.neuroscience.2005.10.014، PMID 16330153، S2CID 1417873.
- "Failures and successes of NMDA receptor antagonists: molecular basis for the use of open-channel blockers like memantine in the treatment of acute and chronic neurologic insults"، NeuroRx، 1 (1): 101–110، يناير 2004، doi:10.1602/neurorx.1.1.101، PMC 534915، PMID 15717010.
- "Subunit- and site-specific pharmacology of the NMDA receptor channel"، Progress in Neurobiology، 59 (3): 279–298، أكتوبر 1999، doi:10.1016/S0301-0082(99)00007-6، PMID 10465381، S2CID 24726102.
- "The glutamate story"، British Journal of Pharmacology، 147 (S1): S100–S108، يناير 2006، doi:10.1038/sj.bjp.0706444، PMC 1760733، PMID 16402093.
- "NMDA receptor subunits: function and pharmacology" (PDF)، Current Opinion in Pharmacology، 7 (1): 39–47، فبراير 2007، doi:10.1016/j.coph.2006.08.011، PMID 17088105، مؤرشف من الأصل (PDF) في 21 سبتمبر 2021.
- "Extrasynaptic NMDARs oppose synaptic NMDARs by triggering CREB shut-off and cell death pathways"، Nature Neuroscience، 5 (5): 405–14، مايو 2002، doi:10.1038/nn835، PMID 11953750، S2CID 659716.
- "Synaptic versus extrasynaptic NMDA receptor signalling: implications for neurodegenerative disorders"، Nature Reviews. Neuroscience، 11 (10): 682–96، أكتوبر 2010، doi:10.1038/nrn2911، PMC 2948541، PMID 20842175.
- "Therapeutic targeting of the pathological triad of extrasynaptic NMDA receptor signaling in neurodegenerations"، The Journal of Experimental Medicine، 214 (3): 569–578، مارس 2017، doi:10.1084/jem.20161673، PMC 5339681، PMID 28209726.
- "Coupling of NMDA receptors and TRPM4 guides discovery of unconventional neuroprotectants"، Science، 370 (6513): eaay3302، أكتوبر 2020، doi:10.1126/science.aay3302، PMID 33033186، S2CID 222210921.
- "The lipophilic bullet hits the targets: medicinal chemistry of adamantane derivatives"، Chemical Reviews، 113 (5): 3516–3604، مايو 2013، doi:10.1021/cr100264t، PMC 3650105، PMID 23432396.
 بوابة طب
بوابة طب بوابة علم الأحياء الخلوي والجزيئي
بوابة علم الأحياء الخلوي والجزيئي بوابة علوم عصبية
بوابة علوم عصبية