مستقبلات جابا
مستقبلات جابا هي فئة من المستقبلات التي تستجيب للناقل العصبي حمض جاما أمينوبوتيريك (جابا)، وهو المركب المثبط الرئيسي في الجهاز الفقري العصبي المركزي عند الناضجين. تؤثر مستقبلات جابا على الإدراك من خلال التنسيق مع العمليات الجلوتاماترجيكية.[1] هناك نوعان من مستقبلات جابا: جابا أ وجابا ب. مستقبلات جابا هي قنوات أيونوتروبية (تعرف أيضا باسم مستقبلات مضادة للأيونات)؛ في حين أن مستقبلات جابا «ب» هي مستقبلات مقترنة بالبروتين G، وتسمى أيضا مستقبلات ميتابوتروبيكية.
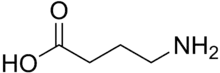
القنوات الأيونوتروبية: جابا أ
اكتُشف منذ فترة طويلة أن الاستجابة السريعة لجابا في الخلايا العصبية التي تم غلقها من قبل بيكوكولين وبيكروتوكسين ترجع إلى التفعيل المباشر لقناة أنيونية.[2][3][4][5][6] سميت هذه القناة فيما بعد بمستقبل جابا.[7] مستقبلات جابا سريعة الاستجابة [8][9][10] هي أعضاء في عائلة من القنوات الأيونوتروبية ذات الحلقة سيس (Cys-loop ligand-gated ion channels). تمتلك أعضاء هذه العائلة الكبيرة والتي تشمل مستقبلات الأسيتيل كولين النيكوتينية ومستقبلات جابا والجلايسين ومستقبلات 5-HT3 ، حلقة مميزة تتكون من رابطة ثنائي الكبريتيد بين اثنين من رواسب السيستين.

في مستقبلات جابا «أ» الأيونوتروبية، يؤدي ارتباط جزيئات جابا بمواقع ارتباطها في الجزء خارج الخلية من المستقبِل إلى فتح المسام الانتقائية لأيون الكلوريد. زيادة مرور الكلوريد تدفع جهد الغشاء نحو عكس جهد أيون الكلور وهو حوالي -65 ملي فولت في الخلايا العصبية، مما يحول دون إطلاق جهد حركة جديد. هذه الآلية هي المسئولة عن التأثير المهدئ لمنبهات الألوستيريك جابا «أ». بالإضافة إلى ذلك، تفعيل مستقبلات جابا يؤدي إلى ما يسمى تثبيط المحول، مما يقلل من استثارة الخلية مستقلةً عن التغيرات في جهد الغشاء.
كانت هناك تقارير عديدة عن مستقبلات جابا المستثارة. وفقا لنظرية استثارة جابا، فإن هذه الظاهرة ناتجة عن زيادة تركيز أيونات الكلوريد داخل الخلية إما أثناء تطور الجهاز العصبي [11][12] أو في بعض أنواع الخلايا.[13][14][15] بعد هذه الفترة من التطور، يتم تنظيم مضخة الكلوريد وإدراجها في غشاء الخلية، لتضخ أيونات الكلوريد في فضاء من الأنسجة خارج الخلية. تنتج فتحات أخرى عن طريق ارتباط جابا بالمستقبلات ثم تنتج استجابات مثبطة. الإفراط في استثارة هذه المستقبلات يؤدي إلى إعادة تشكيل المستقبل، في نهاية المطاف ينثني مستقبل جابا داخل نفسه. ونتيجة لذلك، يتم تثبيط المزيد من الارتباط بمستقبلات جابا وتستقل عن الجهد المثبط بعد المشبكي.
مع ذلك، تم اتهام نظرية استثارة جابا باحتمالية أن تكون مصطنعة نتيجة ظروف تجريبية، حيث أن معظم البيانات التي تم الحصول عليها في المختبر في شريحة من الدماغ كانت عرضة للأوساط غير الفسيولوجية مثل نقص التمثيل الغذائي للطاقة والأضرار العصبية. نشأ الجدل عندما أظهرت عدد من الدراسات أن جابا في شرائح دماغ حديثي الولادة يصبح مثبطا إذا تم تزويد الجلوكوز في سائل التروية بأجسام كيتون، أو البيروفات، أو اللاكتات، [16][17] أو أن استثارة جابا نتجت من تلف الخلايا العصبية.[18] شككت دراسات لاحقة من منشئي ومؤيدي نظرية استثارة جابا في هذه النتائج،[19][20][21] ولكن الحقيقة ظلت بعيدة المنال حتى أمكن توضيح الآثار الحقيقية لجابا بشكل موثوق في الدماغ الحية السليمة. منذ ذلك الحين، وباستخدام التكنولوجيا مثل في كهربية الجسم الحي/ التصوير وعلم الوراثة الضوئية، ذكرت اثنتين من الدراسات على الجسم الحي تأثير جابا على دماغ حديثي الولادة، وكلتاهما أظهرت أن غابا يعد في الواقع مثبط بشكل عام، مع ذكر أن تفعيلها في دماغ القوارض النامية لا يؤدي إلى تنشيط الجهاز كله، [22] وبدلا من ذلك يؤدي إلى انخفاض النشاط.[23][24]
الفئة الفرعية جابا أـ ρ (جابا C سابقا)
هي فئة فرعية من مستقبلات جابا المؤثرة، غير حساسة للمغيرات المعيارية النموذجية لقنوات مستقبلات جابا مثل: البنزوديازيبينات والباربيتورات.[25][26][27] أطلق عليها مستقبلات جاباC.[28][29] تحدث الاستجابات الأصلية لمستقبلات جابا C في خلايا الشبكية ثنائية القطبية أو الأفقية في فصائل الفقاريات.[30][31][32][33]
تتكون مستقبلات جابا C بشكل حصري من الوحدات الفرعية ρ (رو) التي ترتبط بالوحدات الفرعية: مستقبلات جابا أ. على الرغم من أن مصطلح «مستقبلات جابا C» كثيرا ما يستخدم،[34][35][36] فإن جابا c يمكن أن ينظر إليه باعتباره أحد أشكال عائلة مستقبلات جابا أ.[8] جادل آخرون بأن الاختلافات بين مستقبلات غابا C وغابا أ كبيرة بما فيه الكفاية لتبرير الحفاظ على التمييز بين هذين النوعين من مستقبلات جابا.[37][38] مع ذلك، منذ أن مستقبلات جابا C ترتبط ارتباطا وثيقا بتسلسل وبنية ووظيفة مستقبلات جابا أ، ومنذ أن مستقبلات جابا أ الأخرى إلى جانب تلك التي تحتوي على وحدات ρ الفرعية تبدو مماثلة لجابا С في شكلها الصيدلي، أوصت لجنة التسميات في الاتحاد الدولي لعلم الأدوية الأساسية والسريرية (IUPHAR) أن مصطلح جابا c لم يعد مستخدم وينبغي أن تكون هذه المستقبلات ρ بمثابة الفئة الفرعية ρ لمستقبلات جابا أ (جابا أ -ρ).[39]
مستقبلات مرتبطة بالبروتين G : جابا ب
هناك استجابة بطيئة لجابا بوساطة مستقبلات جابا ب، [40] تم تعريفها أصلا على أساس الخصائص الدوائية.[41]
في الدراسات التي تركز على السيطرة على إطلاق الموصلات العصبية، لوحظ أن مستقبلات جابا كانت مسئولة عن تحوير أثار الإطلاق في مجموعة متنوعة من تحضيرات الأنسجة المستقلة. لم يتم حظر قدرة جابا هذه في منع الإطلاق العصبي في هذه التحضيرات من قبل بيكوكولين، ولم تُحاكي من قبل إيسوجوفاسين، ولم تعتمد على أيونات الكلوريد، والتي تعد كلها من سمات مستقبلات جابا. كان الاكتشاف الأكثر إثارة للدهشة اكتشاف أن باكلوفين (β-باراكلوروفينيل جابا)، المستخدم سريريا لعلاج التشنج،[42][43] يحاكي تأثير جابا بطريقة مجسمة.
في وقت لاحق قدمت الدراسات دليلا مباشرا على مواقع ارتباط باكلوفين على الأغشية العصبية المركزية. أكد استنساخ [ الحمض النووي C] [44] أن مستقبلات جابا ب ينتمي إلى عائلة المستقبلات المقترنة بالبروتين G.[45] تم استعراض معلومات إضافية عن مستقبلات جابا ب في أماكن أخرى.[46][47][48][49][50][51][52][53]
المراجع
- Farahmandfar, Maryam؛ Akbarabadi, Ardeshir؛ Bakhtazad, Atefeh؛ Zarrindast, Mohammad-Reza (01 مارس 2016)، "Recovery from ketamine-induced amnesia by blockade of GABA-A receptor in medial prefrontal cortex of mice"، Neuroscience، doi:10.1016/j.neuroscience.2016.02.056، ISSN 1873-7544، PMID 26944606.
- Kuffler SW, Edwards C (نوفمبر 1958)، "Mechanism of gamma aminobutyric acid (GABA) action and its relation to synaptic inhibition"، J. Neurophysiol.، 21 (6): 589–610، PMID 13599049، مؤرشف من الأصل في 03 أغسطس 2004.
- Kravitz EA, Kuffler SW, Potter DD (سبتمبر 1963)، "Gamma-aminobutyric acid and other blocking compounds in crustacea. III. Their relative concentrations in separated motor and inhibitory axons"، J. Neurophysiol.، 26: 739–51، PMID 14065325، مؤرشف من الأصل في 26 مايو 2020.
- Krnjević K, Schwartz S (1967)، "The action of gamma-aminobutyric acid on cortical neurones"، Exp Brain Res، 3 (4): 320–36، doi:10.1007/BF00237558، PMID 6031164.
- Takeuchi A, Takeuchi N (أغسطس 1967)، "Anion permeability of the inhibitory post-synaptic membrane of the crayfish neuromuscular junction"، J. Physiol. (Lond.)، 191 (3): 575–90، doi:10.1113/jphysiol.1967.sp008269، PMC 1365493، PMID 6051794، مؤرشف من الأصل في 11 مايو 2020.
- Takeuchi A, Takeuchi N (نوفمبر 1969)، "A study of the action of picrotoxin on the inhibitory neuromuscular junction of the crayfish"، J. Physiol. (Lond.)، 205 (2): 377–91، doi:10.1113/jphysiol.1969.sp008972، PMC 1348609، PMID 5357245، مؤرشف من الأصل في 19 فبراير 2020.
- Takeuchi A, Onodera K (مارس 1972)، "Effect of bicuculline on the GABA receptor of the crayfish neuromuscular junction"، Nature New Biol.، 236 (63): 55–6، doi:10.1038/236055a0، PMID 4502428.
- Barnard EA, Skolnick P, Olsen RW, Mohler H, Sieghart W, Biggio G, Braestrup C, Bateson AN, Langer SZ (يونيو 1998)، "International Union of Pharmacology. XV. Subtypes of gamma-aminobutyric acidA receptors: classification on the basis of subunit structure and receptor function"، Pharmacol. Rev.، 50 (2): 291–313، PMID 9647870، مؤرشف من الأصل في 6 أغسطس 2009.
- Hevers W, Lüddens H (أغسطس 1998)، "The diversity of GABAA receptors. Pharmapoo and electrophysiological properties of GABAA channel subtypes"، Mol. Neurobiol.، 18 (1): 35–86، doi:10.1007/BF02741459، PMID 9824848.
- Sieghart W, Sperk G (أغسطس 2002)، "Subunit composition, distribution and function of GABA(A) receptor subtypes"، Curr Top Med Chem، 2 (8): 795–816، doi:10.2174/1568026023393507، PMID 12171572.
- Ben-Ari Y, Khazipov R, Leinekugel X, Caillard O, Gaiarsa JL (نوفمبر 1997)، "GABAA, NMDA and AMPA receptors: a developmentally regulated 'ménage à trois'"، Trends Neurosci.، 20 (11): 523–9، doi:10.1016/S0166-2236(97)01147-8، PMID 9364667.
- Taketo M, Yoshioka T (2000)، "Developmental change of GABA(A) receptor-mediated current in rat hippocampus"، Neuroscience، 96 (3): 507–14، doi:10.1016/S0306-4522(99)00574-6، PMID 10717431.
- Tomiko SA, Taraskevich PS, Douglas WW (فبراير 1983)، "GABA acts directly on cells of pituitary pars intermedia to alter hormone output"، Nature، 301 (5902): 706–7، doi:10.1038/301706a0، PMID 6828152.
- Cherubini E, Gaiarsa JL, Ben-Ari Y (ديسمبر 1991)، "GABA: an excitatory transmitter in early postnatal life"، Trends Neurosci.، 14 (12): 515–9، doi:10.1016/0166-2236(91)90003-D، PMID 1726341.
- Lamsa K, Taira T (سبتمبر 2003)، "Use-dependent shift from inhibitory to excitatory GABAA receptor action in SP-O interneurons in the rat hippocampal CA3 area"، J. Neurophysiol.، 90 (3): 1983–95، doi:10.1152/jn.00060.2003، PMID 12750426.
- Rheims, Sylvain؛ Holmgren, Carl D.؛ Chazal, Genevieve؛ Mulder, Jan؛ Harkany, Tibor؛ Zilberter, Tanya؛ Zilberter, Yuri (أغسطس 2009)، "GABA action in immature neocortical neurons directly depends on the availability of ketone bodies"، Journal of Neurochemistry، 110 (4): 1330–1338، doi:10.1111/j.1471-4159.2009.06230.x، PMID 19558450.
- Holmgren, Carl D.؛ Mukhtarov, Marat؛ Malkov, Anton E.؛ Popova, Irina Y.؛ Bregestovski, Piotr؛ Zilberter, Yuri (فبراير 2010)، "Energy substrate availability as a determinant of neuronal resting potential, GABA signaling and spontaneous network activity in the neonatal cortex"، Journal of Neurochemistry، 112 (4): 900–912، doi:10.1111/j.1471-4159.2009.06506.x، PMID 19943846.
- Dzhala, V.؛ Valeeva, G.؛ Glykys, J.؛ Khazipov, R.؛ Staley, K. (21 مارس 2012)، "Traumatic Alterations in GABA Signaling Disrupt Hippocampal Network Activity in the Developing Brain"، Journal of Neuroscience، 32 (12): 4017–4031، doi:10.1523/JNEUROSCI.5139-11.2012.
- Kirmse, K.؛ Witte, O. W.؛ Holthoff, K. (24 نوفمبر 2010)، "GABA Depolarizes Immature Neocortical Neurons in the Presence of the Ketone Body -Hydroxybutyrate"، Journal of Neuroscience، 30 (47): 16002–16007، doi:10.1523/JNEUROSCI.2534-10.2010، PMID 21106838.
- Ruusuvuori, E.؛ Kirilkin, I.؛ Pandya, N.؛ Kaila, K. (17 نوفمبر 2010)، "Spontaneous Network Events Driven by Depolarizing GABA Action in Neonatal Hippocampal Slices are Not Attributable to Deficient Mitochondrial Energy Metabolism"، Journal of Neuroscience، 30 (46): 15638–15642، doi:10.1523/JNEUROSCI.3355-10.2010، PMID 21084619.
- Tyzio, R.؛ Allene, C.؛ Nardou, R.؛ Picardo, M. A.؛ Yamamoto, S.؛ Sivakumaran, S.؛ Caiati, M. D.؛ Rheims, S.؛ Minlebaev, M.؛ Milh, M.؛ Ferre, P.؛ Khazipov, R.؛ Romette, J.-L.؛ Lorquin, J.؛ Cossart, R.؛ Khalilov, I.؛ Nehlig, A.؛ Cherubini, E.؛ Ben-Ari, Y. (05 يناير 2011)، "Depolarizing Actions of GABA in Immature Neurons Depend Neither on Ketone Bodies Nor on Pyruvate"، Journal of Neuroscience، 31 (1): 34–45، doi:10.1523/JNEUROSCI.3314-10.2011، PMID 21209187.
- Kirmse, Knut؛ Kummer, Michael؛ Kovalchuk, Yury؛ Witte, Otto W.؛ Garaschuk, Olga؛ Holthoff, Knut (16 يوليو 2015)، "GABA depolarizes immature neurons and inhibits network activity in the neonatal neocortex in vivo"، Nature Communications، 6: 7750، doi:10.1038/ncomms8750.
- Valeeva, G.؛ Tressard, T.؛ Mukhtarov, M.؛ Baude, A.؛ Khazipov, R. (01 يونيو 2016)، "An Optogenetic Approach for Investigation of Excitatory and Inhibitory Network GABA Actions in Mice Expressing Channelrhodopsin-2 in GABAergic Neurons"، Journal of Neuroscience، 36 (22): 5961–5973، doi:10.1523/JNEUROSCI.3482-15.2016، PMID 27251618.
- Zilberter, M. (05 أكتوبر 2016)، "Reality of Inhibitory GABA in Neonatal Brain: Time to Rewrite the Textbooks?"، Journal of Neuroscience، 36 (40): 10242–10244، doi:10.1523/JNEUROSCI.2270-16.2016، PMID 27707962.
- Sivilotti L, Nistri A (1991)، "GABA receptor mechanisms in the central nervous system"، Prog. Neurobiol.، 36 (1): 35–92، doi:10.1016/0301-0082(91)90036-Z، PMID 1847747.
- Bormann J, Feigenspan A (ديسمبر 1995)، "GABAC receptors"، Trends Neurosci.، 18 (12): 515–9، doi:10.1016/0166-2236(95)98370-E، PMID 8638289.
- Johnston GA (سبتمبر 1996)، "GABAc receptors: relatively simple transmitter -gated ion channels?"، Trends Pharmacol. Sci.، 17 (9): 319–23، doi:10.1016/0165-6147(96)10038-9، PMID 8885697.
- Drew CA, Johnston GA, Weatherby RP (ديسمبر 1984)، "Bicuculline-insensitive GABA receptors: studies on the binding of (-)-baclofen to rat cerebellar membranes"، Neurosci. Lett.، 52 (3): 317–21، doi:10.1016/0304-3940(84)90181-2، PMID 6097844.
- Zhang D, Pan ZH, Awobuluyi M, Lipton SA (مارس 2001)، "Structure and function of GABA(C) receptors: a comparison of native versus recombinant receptors"، Trends Pharmacol. Sci.، 22 (3): 121–32، doi:10.1016/S0165-6147(00)01625-4، PMID 11239575.
- Feigenspan A, Wässle H, Bormann J (يناير 1993)، "Pharmacology of GABA receptor Cl- channels in rat retinal bipolar cells"، Nature، 361 (6408): 159–62، doi:10.1038/361159a0، PMID 7678450.
- Qian H, Dowling JE (يناير 1993)، "Novel GABA responses from rod-driven retinal horizontal cells"، Nature، 361 (6408): 162–4، doi:10.1038/361162a0، PMID 8421521.
- Lukasiewicz PD (يونيو 1996)، "GABAC receptors in the vertebrate retina"، Mol. Neurobiol.، 12 (3): 181–94، doi:10.1007/BF02755587، PMID 8884747.
- Wegelius K, Pasternack M, Hiltunen JO, Rivera C, Kaila K, Saarma M, Reeben M (يناير 1998)، "Distribution of GABA receptor rho subunit transcripts in the rat brain"، Eur. J. Neurosci.، 10 (1): 350–7، doi:10.1046/j.1460-9568.1998.00023.x، PMID 9753143.
- Shimada S, Cutting G, Uhl GR (أبريل 1992)، "gamma-Aminobutyric acid A or C receptor? gamma-Aminobutyric acid rho 1 receptor RNA induces bicuculline-, barbiturate-, and benzodiazepine-insensitive gamma-aminobutyric acid responses in Xenopus oocytes"، Mol. Pharmacol.، 41 (4): 683–7، PMID 1314944، مؤرشف من الأصل في 8 سبتمبر 2008.
- Kusama T, Spivak CE, Whiting P, Dawson VL, Schaeffer JC, Uhl GR (مايو 1993)، "Pharmacology of GABA rho 1 and GABA alpha/beta receptors expressed in Xenopus oocytes and COS cells"، Br. J. Pharmacol.، 109 (1): 200–6، doi:10.1111/j.1476-5381.1993.tb13554.x، PMC 2175610، PMID 8388298.
- Kusama T, Wang TL, Guggino WB, Cutting GR, Uhl GR (مارس 1993)، "GABA rho 2 receptor pharmacological profile: GABA recognition site similarities to rho 1"، Eur. J. Pharmacol.، 245 (1): 83–4، doi:10.1016/0922-4106(93)90174-8، PMID 8386671.
- Chebib M, Johnston GA (أبريل 2000)، "GABA-Activated ligand gated ion channels: medicinal chemistry and molecular biology"، J. Med. Chem.، 43 (8): 1427–47، doi:10.1021/jm9904349، PMID 10780899.
- Bormann J (يناير 2000)، "The 'ABC' of GABA receptors"، Trends Pharmacol. Sci.، 21 (1): 16–9، doi:10.1016/S0165-6147(99)01413-3، PMID 10637650.
- Olsen RW, Sieghart W (سبتمبر 2008)، "International Union of Pharmacology. LXX. Subtypes of γ-Aminobutyric AcidA Receptors: Classification on the Basis of Subunit Composition, Pharmacology, and Function. Update"، Pharmacological Reviews، 60 (3): 243–60، doi:10.1124/pr.108.00505، PMC 2847512، PMID 18790874.
- Bowery NG, Bettler B, Froestl W, Gallagher JP, Marshall F, Raiteri M, Bonner TI, Enna SJ (يونيو 2002)، "International Union of Pharmacology. XXXIII. Mammalian gamma-aminobutyric acid(B) receptors: structure and function"، Pharmacological Reviews، 54 (2): 247–64، doi:10.1124/pr.54.2.247، PMID 12037141.
- Bowery NG, Hill DR, Hudson AL, Doble A, Middlemiss DN, Shaw J, Turnbull M (يناير 1980)، "(-)Baclofen decreases neurotransmitter release in the mammalian CNS by an action at a novel GABA receptor"، Nature، 283 (5742): 92–4، doi:10.1038/283092a0، PMID 6243177.
- Bein HJ (1972)، "Pharmacological differentiations of muscle relaxants"، في Birkmayer W (المحرر)، Spasticity: A Topical Survey، Hans Hubert Bern, Switzerland، ص. 76–89، ISBN 3-456-00390-0.
- Keberle H, Faigle JW (1972)، "Synthesis and structure-activity relationship of the gamma-aminobutyric acid derivatives"، في Birkmayer W (المحرر)، Spasticity: A Topical Survey، Hans Hubert Bern, Switzerland، ص. 76–89، ISBN 3-456-00390-0.
- Hill DR, Bowery NG (مارس 1981)، "3H-baclofen and 3H-GABA bind to bicuculline-insensitive GABA B sites in rat brain"، Nature، 290 (5802): 149–52، doi:10.1038/290149a0، PMID 6259535.
- Kaupmann K, Huggel K, Heid J, Flor PJ, Bischoff S, Mickel SJ, McMaster G, Angst C, Bittiger H, Froestl W, Bettler B (مارس 1997)، "Expression cloning of GABA(B) receptors uncovers similarity to metabotropic glutamate receptors"، Nature، 386 (6622): 239–46، doi:10.1038/386239a0، PMID 9069281.
- Enna SJ (أكتوبر 1997)، "GABAB receptor agonists and antagonists: pharmacological properties and therapeutic possibilities"، Expert Opin Investig Drugs، 6 (10): 1319–25، doi:10.1517/13543784.6.10.1319، PMID 15989503.
- Bowery, N. G.؛ Enna, S. J. (1997)، The GABA receptors، Totowa, NJ: Humana Press، ISBN 0-89603-458-5.
- Kaupmann K, Malitschek B, Schuler V, Heid J, Froestl W, Beck P, Mosbacher J, Bischoff S, Kulik A, Shigemoto R, Karschin A, Bettler B (ديسمبر 1998)، "GABA(B)-receptor subtypes assemble into functional heteromeric complexes"، Nature، 396 (6712): 683–7، doi:10.1038/25360، PMID 9872317.
- Kaupmann K, Schuler V, Mosbacher J, Bischoff S, Bittiger H, Heid J, Froestl W, Leonhard S, Pfaff T, Karschin A, Bettler B (ديسمبر 1998)، "Human γ-aminobutyric acid type B receptors are differentially expressed and regulate inwardly rectifying K+ channels"، Proc. Natl. Acad. Sci. U.S.A.، 95 (25): 14991–6، doi:10.1073/pnas.95.25.14991، PMC 24563، PMID 9844003، مؤرشف من الأصل في 9 يناير 2020.
- Marshall FH, Jones KA, Kaupmann K, Bettler B (أكتوبر 1999)، "GABA receptors - the first 7TM heterodimers"، Trends Pharmacol. Sci.، 20 (10): 396–9، doi:10.1016/S0165-6147(99)01383-8، PMID 10498952.
- Marshall FH, White J, Main M, Green A, Wise A (أغسطس 1999)، "GABA(B) receptors function as heterodimers"، Biochem. Soc. Trans.، 27 (4): 530–5، PMID 10917635.
- Bowery NG, Enna SJ (يناير 2000)، "gamma-aminobutyric acid(B) receptors: first of the functional metabotropic heterodimers"، J. Pharmacol. Exp. Ther.، 292 (1): 2–7، PMID 10604925، مؤرشف من الأصل في 7 أكتوبر 2008.
- Enna SJ (2001)، "GABAB receptor signaling pathways"، في Möhler H (المحرر)، Pharmacology of GABA and Glycine Neurotransmission (Handbook of Experimental Pharmacology) (Volume 150)، Berlin: Springer، ص. 329–342، ISBN 3-540-67616-3.
وصلات خارجية
- IUPHAR GPCR Database - GABAB receptors
- GABA+Receptor في المكتبة الوطنية الأمريكية للطب نظام فهرسة المواضيع الطبية (MeSH).
 بوابة علوم عصبية
بوابة علوم عصبية