محور عصبي
المحور العصبي، أو الليف العصبي أو المحوار، هو امتداد طويل ورفيع للعصبون أو الخلية العصبية عند الفقاريات، ينقل عادةً النبضات الكهربائية التي تُعرف باسم جهود الفعل بعيدًا عن جسم الخلية. تعد وظيفة المحوار نقل المعلومات إلى مختلف العصبونات والعضلات والغدد. في بعض العصبونات الحسية (العصبونات أحادية القطب الكاذبة)، مثل عصبونات اللمس والحرارة، تُدعى المحاور العصبية بالألياف العصبية الواردة وتنتقل النبضات الكهربائية على طول هذه المحاور من المحيط إلى جسم الخلية، ومن جسم الخلية إلى النخاع الشوكي على طول فرع آخر من نفس المحور العصبي. يسبب الخلل في المحاور العصبية العديد من الاضطرابات العصبية الموروثة والمكتسبة، والتي يمكن أن تؤثر على كل من العصبونات المركزية والمحيطية. تُصنف الألياف العصبية في ثلاثة أنواع -مجموعة الألياف العصبية «إيه»، ومجموعة الألياف العصبية «بي» ومجموعة الألياف العصبية «سي». تُحاط المجموعتان «إيه» و«بي» بغمد النخاعين، أما المجموعة «سي» غير نخاعينية. تشمل هذه المجموعات الثلاث كلًا من الألياف الحسية والحركية. يوجد تصنيف آخر يركز على الألياف الحسية فقط، إذ تصنف في أربعة أنواع، النوع Ι والنوع ΙΙ والنوع ΙΙΙ والنوع ΙV.
| المحوار |
|---|
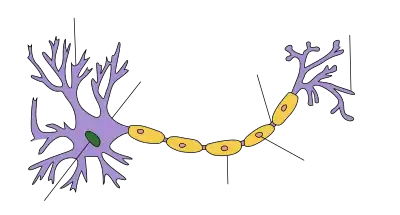
يُعد المحوار واحدًا من نوعي الامتدادات السيتوبلازمية لجسم الخلية العصبية؛ أما النوع الآخر فهو التغصنات العصبية. يتمتع المحوار على عكس التغصنات بالعديد من الميزات، بما في ذلك الشكل (عادةً ما تكون التغصنات مستدقةً بينما تمتاز المحاوير بنصف قطر ثابت)، والطول (تقتصر التغصنات على منطقة صغيرة حول جسم الخلية بينما تكون المحاوير أطول بكثير) والوظيفة (تستقبل التغصنات الإشارات بينما تنقلها المحاوير). لا تملك بعض أنواع العصبونات محاور عصبيةً إذ تنقل إشاراتها عبر التغصنات. في بعض الأنواع، يمكن للمحاوير أن تنبثق من التغصنات العصبية التي تُعرف باسم التغصنات حاملة المحاوير. لا يمكن أن للعصبون أن يحتوي على أكثر من محوار واحد؛ ومع ذلك في بعض اللافقاريات مثل الحشرات أو العلق، يتكون المحوار أحيانًا في مناطق متعددة من العصبون، إذ تعمل بشكل أكثر أو أقل استقلاليةً عن بعضها البعض. يُغطى المحوار بغشاء يُعرف بغمد المحوار؛ وتُدعى السيتوبلازما في المحوار سيتوبلازم المحور العصبي. تتفرع معظم المحاوير، وتكون تفرعاتها غزيرةً في بعض الأحيان. تُسمى نهايات هذه التفرعات بالتغصنات الانتهائية. تُعرف النهاية المنتفخة للتغصنات الانتهائية بالنهاية المحورية التي ترتبط مع جسم عصبون آخر أو تغصناته لتشكل اتصالًا مشبكيًا. تتصل المحاوير مع الخلايا الأخرى –مع عصبونات أخرى عادةً لكن قد تتصل في بعض الأحيان مع خلايا عضلية أو غدية- في مواقع وصل تُدعى المشابك العصبية. في بعض الحالات، قد يشكل محور من عصبون ما تشابكًا مع تغصنات العصبون نفسه، ما يؤدي إلى تشكل مشبك يدعى (Autapse). في المشبك، يقع غشاء المحوار بالقرب من غشاء الخلية المستهدفة، وتعمل البنى الجزيئية الخاصة على نقل الإشارات الكهربائية أو الكهروكيميائية عبر الفجوة. تستمر بعض الموصلات المشبكية على طول المحوار وامتداده – تُسمى هذه الموصلات بالمشابك en passant («العابرة»)، ويمكن أن تصل أعدادها إلى مئات أو حتى آلاف المشابك على طول محوار واحد. تظهر المشابك الأخرى في أطراف نهايات الفروع المحورية العصبية.[1][2]
يستطيع محوار واحد، مع جميع تفرعاته، تعصيب أجزاء متعددة من الدماغ وتوليد آلاف النهايات المشبكية. تشكل مجموعة المحاور سبيلًا عصبيًا في الجهاز العصبي المركزي، وحزمةً عصبيةً في الجهاز العصبي المحيطي. في الثدييات المشيمية، يعد الجسم الثفني أكبر سبيل عصبي من المادة البيضاء في الدماغ، إذ يتكون من 200 مليون محور عصبي تقريبًا في الدماغ البشري.[3]
لمحة تاريخية
يرجع فضل اكتشاف المحور العصبي، عبر تمييزه عن التغصنات العصبية، إلى عالم التشريح الألماني أوتو فريدريش. يعد كل من السويسري ألبرت فون كوليكر والألماني روبرت ريماك أول من حدد وميز القسم الأولي من المحوار. وهب كولكر للمحوار اسمه في عام 1896. وصف لويس أنطوان رانفييه العقد والفجوات الممتدة على طول المحاور العصبية، إذ يُشار الآن إلى ميزات المحوار هذه باسم عقد رانفييه. اقترح عالم التشريح الإسباني سانتياغو رامون إي كاخال أن المحاوير تعد المكونات المخرجة للعصبونات ووصف وظيفتها. طور جوزف إيرلنغر وهربرت غاسر في وقت سابق نظام التصنيف الخاص بالألياف العصبية المحيطية، استنادًا إلى سرعة التوصيل المحوري ووجود النخاعين وحجم الألياف وغيرها. بالإضافة إلى ذلك، استخدم آلن هودجكين وأندرو هكسلي محورًا عصبيًا كبيرًا لحبار (1939) وحصلوا بحلول عام 1952 على وصف كمي كامل للأساس الأيوني في جهد الفعل، ما أدى إلى صياغة نموذج هودجكين-هكسلي. مُنح كل منهما لاحقًا جائزة نوبل لعام 1963 بفضل هذا العمل. تمدد استخدام هذه الصيغ التي توضح التوصيل المحوري العصبي إلى الفقاريات عن طريق معادلات فرانكنهاوزر-هكسلي. تطور فهم الأساس الكيميائي الحيوي لانتشار جهد الفعل، ويشمل العديد من التفاصيل حول القنوات الأيونية الفردية.[4]
المراجع
- Yau KW (ديسمبر 1976)، "Receptive fields, geometry and conduction block of sensory neurones in the central nervous system of the leech"، The Journal of Physiology، 263 (3): 513–38، doi:10.1113/jphysiol.1976.sp011643، PMC 1307715، PMID 1018277.
- Squire, Larry (2013)، Fundamental neuroscience (ط. 4th)، Amsterdam: Elsevier/Academic Press، ص. 61–65، ISBN 978-0-12-385-870-2.
- Luders E, Thompson PM, Toga AW (أغسطس 2010)، "The development of the corpus callosum in the healthy human brain"، The Journal of Neuroscience، 30 (33): 10985–90، doi:10.1523/JNEUROSCI.5122-09.2010، PMC 3197828، PMID 20720105.
- Debanne D, Campanac E, Bialowas A, Carlier E, Alcaraz G (أبريل 2011)، "Axon physiology"، Physiological Reviews، 91 (2): 555–602، doi:10.1152/physrev.00048.2009، PMID 21527732، مؤرشف من الأصل في 09 مارس 2020.
 بوابة علم الأحياء
بوابة علم الأحياء بوابة علم الأحياء الخلوي والجزيئي
بوابة علم الأحياء الخلوي والجزيئي بوابة علوم عصبية
بوابة علوم عصبية بوابة طب
بوابة طب