Paraves
Paraves are a widespread group of theropod dinosaurs that originated in the Middle Jurassic period. In addition to the extinct dromaeosaurids, troodontids, anchiornithids, and possibly the scansoriopterygids, the group also contains the avialans, which include diverse extinct taxa as well as the over 10,000 species of living birds.[1] Basal members of Paraves are well known for the possession of an enlarged claw on the second digit of the foot, which was held off the ground when walking in some species.[2] A number of differing scientific interpretations of the relationships between paravian taxa exist. New fossil discoveries and analyses make the classification of Paraves an active subject of research.[3]
| Paravians | |
|---|---|
 | |
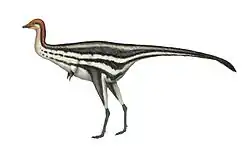
| Examples of basal paravian dinosaurs | |
| Scientific classification | |
| Domain: | Eukaryota |
| Kingdom: | Animalia |
| Phylum: | Chordata |
| Clade: | Dinosauria |
| Clade: | Saurischia |
| Clade: | Theropoda |
| Clade: | Pennaraptora |
| Clade: | Paraves Sereno, 1997 |
| Subgroups | |
| Synonyms | |
| |

Description
Like other theropods, all paravians are bipedal, walking on their two hind legs.[4]
The teeth of basal paravians were curved and serrated, but not blade-like except in some specialized species such as Dromaeosaurus albertensis. The serrations on the front edge of dromaeosaurid and troodontid teeth were very small and fine, while the back edge had serrations which were very large and hooked.[5]
Most of the earliest paravian groups were carnivorous, though some smaller species (especially among the troodontids and early avialans) are known to have been omnivores, and it has been suggested that an omnivorous diet was the ancestral state for this group, with strict carnivory evolving in some specialized lineages.[6][5] Fossils also suggest that legs and feet covered with feathers was an ancestral condition, possibly having originated in the Coelurosauria, even if this trait was later lost in more advanced birds.[7]
Wings
Paravians generally have long, winged forelimbs, though these have become smaller in many flightless species and some extinct lineages that evolved before flight. The wings usually bore three large, flexible, clawed fingers in early forms.[5] The fingers became fused and stiffened and the claws highly reduced or lost in some advanced lineages. An increasingly asymmetric wrist joint, a trend that can be traced back to primitive coelurosaurs, allowed the forelimbs to elongate and an elaboration of their plumage, traits that made the evolution of flapping flight possible.[8]
Many early members of Paraves had both well-developed wings and long feathers on the hind legs, which in some cases, formed a second set of airfoils. These species, most famously represented by Microraptor gui, have often been referred to as "four winged dinosaurs".[9][10][11] Though it has been suggested that these hind wings would have prevented some paravians from getting around on the ground, and that they must have lived in trees, there is very little evidence that any of the earliest paravians were capable of climbing. This apparent paradox was addressed by later studies which showed that early paravians like Microraptor were capable of flapping flight and powered launching from the ground into the air without relying on climbing. Microraptor in particular also seems to represent a case of flight evolving independently of the bird lineage within Paraves.[12]
Feet and sickle claw
Most theropods walked with three toes contacting the ground, but fossilized footprint tracks confirm that many basal paravians, including dromaeosaurids, troodontids, and some early avialans, held the second toe off the ground in a hyperextended position, with only the third and fourth toes bearing the weight of the animal. This is called functional didactyly.[2] The enlarged second toe bore an unusually large, curved sickle-shaped claw (held off the ground or 'retracted' when walking). This claw was especially large and flattened from side to side in the large-bodied predatory eudromaeosaurs.[13] In these early species, the first toe (hallux) was usually small and angled inward toward the center of the body, but only became fully reversed in more specialized members of the bird lineage.[5] One species, Balaur bondoc, possessed a first toe which was highly modified in parallel with the second. Both the first and second toes on each foot of B. bondoc were held retracted and bore enlarged, sickle-shaped claws.[14]

One of the best-known features of paravians is the presence of an enlarged and strongly curved "sickle claw" on a hyper-extendible second toe, modified to hold the sickle claw clear of the ground when walking, most notably developed in the dromaeosaurids and troodontids. While this characteristic claw and its associated modifications to the anatomy of the foot (such as a shortened metatarsus in eudromaeosaurs) had been known since the mid-20th century, their possible functions were the subject mainly of speculation, and few actual studies were published. Initial speculation regarded the claws as slashing implements used to disembowel large prey. In this scenario, the shortened upper foot would serve as an anchor point for powerful tendons to improve kicking ability. However, subsequent studies of the actual claw shape showed that the underside of the claw was only weakly keeled and would not have been an effective cutting instrument. Instead, it appeared to be more of a hooking implement. Manning et al. suggested in 2006 that the claws were similar to crampons and were used for climbing, and in the case of larger species or individuals, climbing up the flanks of very large prey.[15]
A larger study of sickle-claw function, published in 2011 by Fowler and colleagues, concluded that the earlier study by Manning and colleagues was correct and that the "sickle claws" would have been ineffective as cutting weapons. They compared the claw and overall foot anatomy of various primitive species with modern birds to shed light on their actual function. Fowler and colleagues showed that many modern predatory birds also have enlarged claws on the second toes. In modern raptors, these claws are used to help grip and hold prey of sizes smaller than or equal to the predator, while the birds use their body weight to pin their prey to the ground and eat it alive.[5] Fowler and colleagues suggested that this behavior is entirely consistent with the anatomy of advanced dromaeosaurids like Deinonychus, which had slightly opposing first toes and strong tendons in the toes and foot. This makes it likely that advanced dromaeosaurids also used their claws to puncture and grip their prey to aid in pinning it to the ground, while using shallow wing beats and tail movements to stabilize themselves.[5] Other lines of evidence for this behavior include teeth which had large, hooked serrations only on the back edge (useful in pulling flesh upward rather than slicing it) and large claws on the wings (for greater maneuvering of prey while mantling it with the wings).[5]
In more primitive dromaeosaurids and in troodontids, the feet were not as specialized and the claws were not as large or as hooked. Additionally, the toe joints allowed more range of motion than the simple up-down movements of advanced dromaeosaurids. This makes it likely that these species specialized in smaller prey that could be pinned using only the inner toes, not requiring the feet to be as strong or sturdy.[5]
Dichotomy in body sizes
Extreme examples of miniaturization and progenesis are found in Paraves.[16] The ancestors of Paraves first started to shrink in size in the early Jurassic 200 million years ago, and fossil evidence shows that this theropod line evolved new adaptations four times faster than other groups of dinosaurs,[17] and was shrinking 160 times faster than other dinosaur lineages were growing.[18] Turner et al. (2007) suggested that extreme miniaturization was ancestral for the clade, whose common ancestor has been estimated to have been around 65 centimetres (26 in) long and 600–700 grams (21–25 oz) in mass. In Eumaniraptora, both Dromaeosauridae and Troodontidae went later through four independent events of gigantism, three times in dromaeosaurids and once in troodontids, while the body mass continued to decrease in many forms within Avialae.[19] Fossils show that all the earliest members of Paraves found to date started out as small, while Troodontidae and Dromaeosauridae gradually increased in size during the Cretaceous period.[19]
Evolution
Relationships
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Cladogram following the results of a phylogenetic study by Lefèvre et al., 2017[20] |
| |||||||||||||||||||||||||||||||||||||||||||||
| Cladogram following the results of a phylogenetic study by Cau et al., 2017[21] |
Paraves is a branch-based clade defined to include all dinosaurs which are more closely related to birds than to oviraptorosaurs. The ancestral paravian is the earliest common ancestor of birds, dromaeosaurids, and troodontids which was not also ancestral to oviraptorosaurs. Paraves often comprises three major sub-groups: Avialae, including Archaeopteryx and modern birds, as well as the dromaeosaurids and troodontids, which may or may not form a natural group.
The name 'Paraves' (Greek pará, par' 'beside, near' + Latin aves, plural of avis 'bird') was coined by Paul Sereno in 1997.[22] The clade was defined by Sereno in 1998 as a branch-based clade containing all Maniraptora closer to Neornithes (which includes all the birds living in the world today) than to Oviraptor.[23]
In 1997, a node-based clade called Eumaniraptora ("true maniraptorans") was named by Padian, Hutchinson and Holtz. They defined their clade to include only avialans and deinonychosaurs. Paraves and Eumaniraptora are generally considered to be synonyms, though some phylogenetic studies suggest that the two groups have a similar but not identical content; Agnolín and Novas (2011) recovered scansoriopterygids and alvarezsaurids as paravians that were not eumaniraptorans,[24] while Turner, Makovicky and Norell (2012) recovered Epidexipteryx as the only known non-eumaniraptoran paravian.[1]
Since the 1960s, the dromaeosaurids and troodontids have often been classified together in a group or clade named the Deinonychosauria, initially based primarily on the presence of a retractable second toe with sickle-claw (now also known to be present in some avialans). The name Deinonychosauria was coined by Ned Colbert and Dale Russell in 1969, and defined as a clade (all theropods closer to dromaeosaurids than to birds) by Jacques Gauthier in 1986.
However, several more recent studies have cast doubt on the hypothesis that dromaeosaurids and troodontids were more closely related to each other than either was to birds, instead finding that troodontids were more closely related to birds than to dromaeosaurids.[3][25][26] Because Deinonychosauria was originally defined as all animals closer to dromaeosaurids than to birds without specific reference to troodontids, this would render Deinonychosauria a synonym of Dromaeosauridae.[26] The clade containing avialans, microraptorians, unenlagiids, Anchiornis, and Xiaotingia to the exclusion of Eudromaeosauria was named Averaptora by Agnolín and Novas (2013), defined as all animals closer to Passer than to Dromaeosaurus.[27] Most studies use a similar definition for Avialae,[28] which Agnolín and Novas redefine as the least inclusive clade including Archaeopteryx and modern birds.[27] Averaptora additionally contains troodontids according to Cau et al. (2017) and other phylogenies in which find Eudromaeosauria to be an outgroup to a clade including Troodontidae and Avialae.[21]
In 2015 Chatterjee created Tetrapterygidae in the second edition of his book The Rise of Birds: 225 Million Years of Evolution, where he included Microraptor, Xiaotingia, Aurornis, and Anchiornis; together they were proposed to be the sister group of the Avialae.[29] Paraves, Eumaniraptora and Averaptora are often considered to be synonyms, depending on the interpretation of paravian systematics. Deinonychosauria will become a synonym of Dromaeosauridae when troodontids are found to form a clade with Avialae to the exclusion of Dromaeosauridae. Tetrapterygidae is a polyphyletic grouping of four-winged basal paravian genera.
Origin and early evolution
Paravians diverged from other maniraptorans around 165 Mya. Then, around 110–90 Mya, the ancestors of Neornithes (modern birds) split from the other paravians.[30]
Other than the crown group of modern birds, which are direct descendants in the stem lineage of Paraves, there are no extant survivors or genetic material, so their entire phylogeny is inferred only from the fossil record.[31] The prototypical fossil is Archaeopteryx, of which 11 specimens have been found, both complete and partial.[32]
See also
- Origin of birds
- Origin of avian flight
- Evolution of birds
- Biogeography of paravian dinosaurs
- The thecodont hypothesis of bird origins
- Proavis, a hypothetical dinosaur-bird intermediary
- Temporal paradox (paleontology)
- List of fossil bird genera
- Protoavis texensis, a putative but possibly chimeric Triassic bird
References
- Alan Hamilton Turner, Peter J. Makovicky and Mark Norell (2012). "A review of dromaeosaurid systematics and paravian phylogeny". Bulletin of the American Museum of Natural History. 371: 1–206. doi:10.1206/748.1. hdl:2246/6352. S2CID 83572446.
- Li, Rihui; Lockley, M.G.; Makovicky, P.J.; Matsukawa, M.; Norell, M.A.; Harris, J.D.; Liu, M. (2007). "Behavioral and faunal implications of Early Cretaceous deinonychosaur trackways from China". Naturwissenschaften. 95 (3): 185–91. Bibcode:2008NW.....95..185L. doi:10.1007/s00114-007-0310-7. PMID 17952398. S2CID 16380823.
- Agnolin, Federico L.; Motta, Matias J.; Brissón Egli, Federico; Lo Coco, Gastón; Novas, Fernando E. (2019-02-12). "Paravian Phylogeny and the Dinosaur-Bird Transition: An Overview". Frontiers in Earth Science. 6. doi:10.3389/feart.2018.00252. hdl:11336/130197. ISSN 2296-6463.
- Mayr, G. (Oct 2016). Avian Evolution: The Fossil Record of Birds and its Paleobiological Significance (1 ed.). p. Ch. 2. ISBN 978-1119020769.
- Fowler, D.W.; Freedman, E.A.; Scannella, J.B.; Kambic, R.E. (2011). "The Predatory Ecology of Deinonychus and the Origin of Flapping in Birds". PLOS ONE. 6 (12): e28964. Bibcode:2011PLoSO...628964F. doi:10.1371/journal.pone.0028964. PMC 3237572. PMID 22194962.
- Zanno, L.E.; Makovicky, P.J. (2011). "Herbivorous ecomorphology and specialization patterns in theropod dinosaur evolution". Proc Natl Acad Sci USA. 108 (1): 232–237. Bibcode:2011PNAS..108..232Z. doi:10.1073/pnas.1011924108. PMC 3017133. PMID 21173263.
- 125-Million-Year-Old Biplanes: New Evidence Suggests the Earliest Bird Species Had Feathers on their Hind Limbs
- The asymmetry of the carpal joint and the evolution of wing folding in maniraptoran theropod dinosaurs
- Hu, Dongyu; Lianhi, Hou; Zhang, Lijun; Xu, Xing (2009). "A pre-Archaeopteryx troodontid theropod from China with long feathers on the metatarsus". Nature. 461 (7264): 640–643. Bibcode:2009Natur.461..640H. doi:10.1038/nature08322. PMID 19794491. S2CID 205218015.
- Xing, X.; Zhou, Z.; Wang, X.; Kuang, X.; Zhang, F.; Du, X. (2003). "Four-winged dinosaurs from China" (PDF). Nature. 421 (6921): 335–340. Bibcode:2003Natur.421..335X. doi:10.1038/nature01342. PMID 12540892. S2CID 1160118.
- Xu, X.; Zhang, F. (2005). "A new maniraptoran dinosaur from China with long feathers on the metatarsus". Naturwissenschaften. 92 (4): 173–177. Bibcode:2005NW.....92..173X. doi:10.1007/s00114-004-0604-y. PMID 15685441. S2CID 789908.
- Dececchi, T.A.; Larsson, H.C.E.; Habib, M.B. (2016). "The wings before the bird: an evaluation of flapping-based locomotory hypotheses in bird antecedents". PeerJ. 4: e2159. doi:10.7717/peerj.2159. PMC 4941780. PMID 27441115.
- Longrich, N.R.; Currie, P.J. (2009). "A microraptorine (Dinosauria–Dromaeosauridae) from the Late Cretaceous of North America". PNAS. 106 (13): 5002–7. Bibcode:2009PNAS..106.5002L. doi:10.1073/pnas.0811664106. PMC 2664043. PMID 19289829.
- Z., Csiki; Vremir, M.; Brusatte, S. L.; Norell, M. A. (2010). "An aberrant island-dwelling theropod dinosaur from the Late Cretaceous of Romania". Proceedings of the National Academy of Sciences of the United States of America. 107 (35): 15357–61. Bibcode:2010PNAS..10715357C. doi:10.1073/pnas.1006970107. PMC 2932599. PMID 20805514. Supporting Information
- Manning, P.L.; Payne, D.; Pennicott, J.; Barrett, P.M.; Ennos, R.A. (2006). "Dinosaur killer claws or climbing crampons?". Biology Letters. 2 (1): 110–112. doi:10.1098/rsbl.2005.0395. PMC 1617199. PMID 17148340.
- Bhullar, B.A.; et al. (2012). "Birds have paedomorphic dinosaur skulls". Nature. 487 (7406): 223–226. Bibcode:2012Natur.487..223B. doi:10.1038/nature11146. PMID 22722850. S2CID 4370675.
- Dinosaurs 'shrank' regularly to become birds
- "How Dinosaurs Shrank and Became Birds". Scientific American. Archived from the original on 2023-07-20.
- Turner, Alan H.; Pol, Diego; Clarke, Julia A.; Erickson, Gregory M.; Norell, M. (2007). "A basal dromaeosaurid and size evolution preceding avian flight". Science. 317 (5843): 1378–1381. Bibcode:2007Sci...317.1378T. doi:10.1126/science.1144066. PMID 17823350.
- Lefèvre, Ulysse; Cau, Andrea; Cincotta, Aude; Hu, Dongyu; Chinsamy, Anusuya; Escuillié, François; Godefroit, Pascal (2017). "A new Jurassic theropod from China documents a transitional step in the macrostructure of feathers". The Science of Nature. 104 (9–10): 74. Bibcode:2017SciNa.104...74L. doi:10.1007/s00114-017-1496-y. PMID 28831510. S2CID 32780661.
- Cau, Andrea; Beyrand, Vincent; Voeten, Dennis F. A. E.; Fernandez, Vincent; Tafforeau, Paul; Stein, Koen; Barsbold, Rinchen; Tsogtbaatar, Khishigjav; Currie, Philip J.; Godefroit, Pascal (2017-12-06). "Synchrotron scanning reveals amphibious ecomorphology in a new clade of bird-like dinosaurs". Nature. 552 (7685): 395–399. doi:10.1038/nature24679. ISSN 1476-4687.
- Sereno, P. C. (1997). "The origin and evolution of dinosaurs" (PDF). Annual Review of Earth and Planetary Sciences. 25: 435–489. Bibcode:1997AREPS..25..435S. doi:10.1146/annurev.earth.25.1.435.
- Sereno, P. C. (1998). "A rationale for phylogenetic definitions, with application to the higher level taxonomy of Dinosauria". Neues Jahrbuch für Geologie und Paläontologie. 210: 41–83. doi:10.1127/njgpa/210/1998/41.
- Agnolín, Federico L.; Novas, Fernando E. (2011). "Unenlagiid theropods: are they members of the Dromaeosauridae (Theropoda, Maniraptora)?". Anais da Academia Brasileira de Ciências. 83 (1): 117–162. doi:10.1590/S0001-37652011000100008. PMID 21437379.
- Godefroit, Pascal; Cau, Andrea; Hu, Dong-Yu; Escuillié, François; Wu, Wenhao; Dyke, Gareth (2013). "A Jurassic avialan dinosaur from China resolves the early phylogenetic history of birds". Nature. 498 (7454): 359–362. Bibcode:2013Natur.498..359G. doi:10.1038/nature12168. PMID 23719374. S2CID 4364892.
- Mortimer, M. (2012): The Theropod Database: Phylogeny of Theropoda. Retrieved 2013-AUG-15.
- Federico L. Agnolín and Fernando E. Novas (2013). Avian ancestors. A review of the phylogenetic relationships of the theropods Unenlagiidae, Microraptoria, Anchiornis and Scansoriopterygidae. SpringerBriefs in Earth System Sciences. pp. 1–96. doi:10.1007/978-94-007-5637-3. ISBN 978-94-007-5636-6. S2CID 199493087.
- Senter, P (2007). "A new look at the phylogeny of Coelurosauria (Dinosauria: Theropoda)". Journal of Systematic Palaeontology. 5 (4): 429–463. doi:10.1017/S1477201907002143. S2CID 83726237.
- Chatterjee, S. (2015). The rise of birds: 225 million years of evolution. Johns Hopkins University Press, 45-48.
- Claramunt, S.; Cracraft, J. (11 December 2015). "A new time tree reveals Earth history's imprint on the evolution of modern birds". Science Advances. 1 (11): e1501005. Bibcode:2015SciA....1E1005C. doi:10.1126/sciadv.1501005. PMC 4730849. PMID 26824065.
- Fountaine, Toby M.R.; Benton, Michael J.; Dyke, Gareth J.; Nudds, Robert L. (4 February 2005). "The quality of the fossil record of Mesozoic birds". Proc Biol Sci. 272 (1560): 289–294. doi:10.1098/rspb.2004.2923. PMC 1634967. PMID 15705554.
- Foth, Christian; Tischlinger, Helmut; Rauhut, Oliver W.M. (2 July 2014). "New specimen of Archaeopteryx provides insights into the evolution of pennaceous feathers". Nature. 511 (7507): 79–82. Bibcode:2014Natur.511...79F. doi:10.1038/nature13467. PMID 24990749. S2CID 4464659.
External links







.png.webp)


.jpg.webp)





| Anatomy | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Behaviour | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Evolution | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Fossil birds |
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Human interaction | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Lists | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
