Axo-axonic synapse
An axo-axonic synapse is a type of synapse, formed by one neuron projecting its axon terminals onto another neuron's axon.[1]
Axo-axonic synapses have been found and described more recently than the other more familiar types of synapses, such as axo-dendritic synapses and axo-somatic synapses. The spatio-temporal properties of neurons get altered by the type of synapse formed between neurons. Unlike the other types, the axo-axonic synapse does not contribute towards triggering an action potential in the postsynaptic neuron. Instead, it affects the probability of neurotransmitter release in the response to any action potential passing through the axon of the postsynaptic neuron.[2] Thus, axo-axonic synapses appear to be very important for the brain in achieving a specialized neural computation.
Axo-axonic synapses are found throughout the central nervous system, including in the hippocampus, cerebral cortex and striatum in mammals;[3][4][5] in the neuro-muscular junctions in crustaceans;[6][7] and in the visual circuitry in dipterans.[8] Axo-axonic synapses can induce either inhibitory or excitatory effects in the postsynaptic neuron.[6][7][9] A classic example of the role of axo-axonic synapses is causing inhibitory effects on motoneurons in the spinal-somatic reflex arc. This phenomenon is known as presynaptic-inhibition.[10][11]
Background
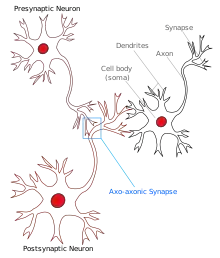
Complex interconnections of neurons form neural networks, which are responsible for various types of computation in the brain. Neurons receive inputs mainly through dendrites, which play a role in spatio-temporal computation, leading to the firing of an action potential which subsequently travels to synaptic terminals passing through axons.[12] Based on their locations, synapses can be classified into various kinds, such as axo-dendritic synapse, axo-somatic synapse, and axo-axonal synapse. The prefix here indicates the part of the presynaptic neuron (i.e., ‘axo-’ for axons), and the suffix represents the location where the synapse is formed on the postsynaptic neuron (i.e., ‘-dendritic’ for dendrites, ‘-somatic’ for cell body and ‘-axonic’ for synapses on axons).[13] Synapse location will govern the role of that synapse in a network of neurons. In axo-dendritic synapses, the presynaptic activity will affect the spatio-temporal computation in postsynaptic neurons by altering electrical potential in the dendritic branch. Whereas the axo-somatic synapse will affect the probability of firing an action potential in the postsynaptic neuron by causing inhibitory or excitatory effects directly at the cell body.[14]
Whereas the other types of synapses modulate postsynaptic neural activity, the axo-axonic synapses show subtle effects on the network-level neural information transfer. In such synapses, the activity in presynaptic neurons will not change the membrane potential (i.e., depolarize or hyperpolarize) of the cell body of postsynaptic neurons because presynaptic neurons project directly on the axons of the postsynaptic neurons. Thus, the axo-axonic synapse will mainly affect the probability of neurotransmitter vesicle release in response to an action potential firing in the postsynaptic neuron. Unlike other kinds of synapses, the axo-axonic synapse manipulates the effects of a postsynaptic neuron's firing on the neurons further downstream in the network.[2] Due to the mechanism of how axo-axonic synapses work, most of these synapses are inhibitory,[6] and yet a few show excitatory effects in postsynaptic neurons.[9]
History
The first direct evidence of the existence of axo-axonic synapses was provided by E. G. Gray in 1962. Gray produced electron microscopy photographs of axo-axonic synapses formed on the terminals of muscle afferents involved in the spinal somatic reflex arc in a cat's spinal cord slices.[15] Later, Gray coined the term ‘axo-axonic’ after getting photographic confirmation from as many as twelve axo-axonic synapses. Within the next two years, scientists found axo-axonic synapses in various other places in the nervous system in different animals, such as in the retina of cats and pigeons,[16] in the lateral geniculate nucleus of monkeys,[17] in the olfactory bulb of mice,[18] and in various lobes in the octopus brain.[19] This further confirmed the existence of axo-axonic synapses in the brain across animal phyla.
Prior to the discovery of axo-axonic synapses, physiologists predicted the possibility of such mechanisms as early as in year 1935, following their observations of electrophysiological recordings and quantal analysis of brain segments.[20] They had observed inhibitory responses in postsynaptic motoneurons in the slice preparation of the monosynaptic reflex arc. During simultaneous recordings from presynaptic and postsynaptic neurons, the physiologists could not make sense of the infrequent inhibition observed in the postsynaptic neuron, with no membrane potential changes in the presynaptic neuron. At that time, this phenomenon was known as “presynaptic inhibitory action”, the term proposed by Karl Frank in 1959 [14] and later well summarized by John Eccles in his book.[10] After Gray's finding of the axo-axonic synapse in 1962, scientists confirmed that this phenomenon was in fact due to the axo-axonic synapse present in the reflex arc.[10]
More recently, in 2006 researchers discovered the first evidence of excitatory effects caused by an axo-axonic synapse. They found that GABAergic neurons project onto the axons of pyramidal cells in the cerebral cortex to form axo-axonic synapse and elicit excitatory effects in cortical microcircuits.[9]
Function
Below are the brain locations where axo-axonic synapses are found in different animals.
Cerebellar cortex
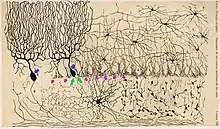
The axo-axonic synapse in the cerebellar cortex originally appeared in one of the drawings of Santiago Ramón y Cajal in his book published in 1909.[21] Later using electron microscopy, it was confirmed that the basket cell axon projects on the axon hillock of Purkinje cells in the cerebellar cortex in cats and other mammals, forming axo-axonic synapses.[5] The first electrophysiological characterization of an axo-axonic synapse formed on Purkinje cells was done in 1963, where the presynaptic basket cell axons were found to inhibit the terminal output of postsynaptic Purkinje cells through the axo-axonic synapse.[22] Network-level study revealed that the granule cells (a.k.a. the parallel fibers) which activated Purkinje cells, also activated the basket cells which subsequently inhibited the effect of Purkinje cells on the downstream network.[23]
Cerebral cortex
Axo-axonic synapses are found In the visual cortex (in V1 and V2) in mammals, and have been well studied in cats, rats and primates such as monkeys.[4][24][25][26][27] The synapse is formed on the initial segments of the axons of pyramidal cells in several layers in the visual cortex. The projecting neurons for these synapses come from various parts of the central nervous system and neocortex. Similarly, axo-axonic synapses are found in the motor cortex, in the subiculum and in the piriform cortex.[4] In the striate cortex, as the Golgi's method and electron microscopy revealed, as many as five axo-axonic synapses are formed onto a single pyramidal cell.[4] In the cerebral cortex, inhibitory axo-axonic synapses may play a widespread role in network level activity by enabling synchronized firing of pyramidal cells, essentially by modulating the threshold for output of these cells.[27][4] These synapses are also found on the initial segments of axons in pyramidal cells in the somatosensory cortex, and in the primary olfactory cortex which are found to be the inhibitory kind.[28][29] Studying the locations of axo-axonic synapses in the primary olfactory cortex, researchers have suggested that axo-axonic synapses may play a critical role in synchronizing oscillations in the piriform cortex (in the olfactory cortex), which aids olfaction.[30] The axo-axonic synapses are also found in the hippocampus. These synapses are found to be formed mainly on principal cells in stratum oriens and stratum pyramidale and rarely on stratum radiatum; they commonly receive projections from GABAergic local interneurons.[31] The horizontal interneurons show a laminar distribution of dendrites and are involved in axo-axonic synapses in the hippocampus, which get direct synaptic inputs from CA1 pyramidal cells.[3] Thus, in general, these studies indicate that axo-axonic synapses can provide a basic mechanism of information processing in the cerebral cortex.[32][30][31]
Basal ganglia
Microscopy studies in the striatum previously suggested rare occurrence of axo-axonic synapses in individual sections. Extrapolations from the topological data suggest much higher counts of such synapses in the striatum where the therapeutic role of the axo-axonic synapses in treating schizophrenia has been postulated previously.[33] In this study, authors examined 4,811 synapses in rat striatum sections, and 15 of them were found to be the axo-axonic synapses. These axo-axonic synapses are formed by dopaminergic inhibitory interneurons (on the presynaptic side) projecting onto the axons of glutamatergic cortico-striatal fibers in the rat striatum.[33]
Brainstem
Axo-axonic synapses are found in the spinal trigeminal nucleus in the brainstem.[34] Electron microscopy studies on the kitten brainstem quantified synaptogenesis of axo-axonic synapses in the spinal trigeminal nucleus at different development ages of the brain. Authors identified the synapses by counting vesicles released in the synaptic cleft, which can be observed in the micrographs. Axo-axonic contacts are shown to consistently increase throughout the development period, starting from the age of 3 hours to the age of 27 days in kittens. The highest rate of synaptogenesis is during the first 3 to 6 days, at the end of which, the kitten's spinal trigeminal nucleus will have nearly half of the axo-axonic synapses present in adult cats. Later, between 16 and 27 days of age, there is another surge of axo-axonic synaptogenesis.[34] Axo-axonic synapses are also observed in the solitary nucleus (also known as nucleus of the solitary tract) uniquely in the commissural portion in the neuroanatomical studies, which used 5-hydroxydopamine to label axo-axonic synapses. Axo-axonic synapses are formed on baroreceptor terminals by the presynaptic adrenergic fibers, and are proposed to play a role in baroreflex.[35]
Spinal Cord
Axo-axonic synapses are found in the mammalian spinal reflex arc[36][37][38] and in Substantia gelatinosa of Rolando (SGR).[39] In the spinal cord, axo-axonic synapses are formed on the terminals of sensory neurons with presynaptic inhibitory interneurons. These synapses are first studied using intracellular recordings from the spinal motoneurons in cats, and have been shown to cause presynaptic inhibition.[40] This seems to be a common mechanism in spinal cords, in which GABAergic interneurons inhibit presynaptic activity in sensory neurons and eventually control activity in motor neurons enabling selective control of muscles.[41] In efforts to quantify the occurrence of axo-axonic synapses in the SGR region in rats, 54 such synapses were found among the total 6,045 synapses examined. These 54 axo-axonic synapses were shown to have either agranular vesicles or large granular vesicles.[39]
Vestibular system
Axo-axonic synapses are found in the lateral vestibular nucleus in rats. Axo-axonic synapses are formed from the small axons of interneurons onto the axon terminals of large axons, which are upstream to the main dendritic stem.[42] Interestingly, the authors claimed that axo-axonic synapses, which are abundant in rats, are absent in the lateral vestibular nucleus in cats.[42] They note that the types of axon terminals identified and described in cats are all found in rats, but the reverse is not true because the axons forming the axo-axonic synapses are missing in cats. These synapses are proposed to enable complex neural computation for the vestibular reflex in rats.[42]
Hindbrain
Axo-axonic synapses are found in the mauthner cells in goldfish.[43][44] The axon hillock and initial axon segments of mauthner cells receive terminals from extremely fine unmyelinated fibers, which cover the axon hillock with helical projections. These helical projections around mauthner cells are also known as the axon cap. The difference between the axo-axonic synapses and other synapses on mauthner cells is that synapses on dendrites and soma receive myelinated fibers, while axons receive unmyelinated fibers.[43][44] Mauthner cells are big neurons which are involved in fast escape reflexes in fish. Thus, these axo-axonic synapses could selectively disable the escape network by controlling the effect of mauthner cells on the neural network further downstream. Studying the morphological variation of the axo-axonic synapses at the axon hillock in mauthner cells suggests that, evolutionarily, these synapses are more recent than the mauthner cells. Response to the startle can be mapped phylogenetically, which confirms that basal actinopterygian fish, with little to no axo-axonic synapses on mauthner cells, show worse escape response than fish with axo-axonic synapses.[45]
Neuromuscular junction
Inhibitory axo-axonic synapses are found in the crustacean neuromuscular junctions and have been widely studied in Crayfish.[6][7][46] Axo-axonic synapses are formed on the excitatory axons as a postsynaptic neuron by the motor neurons from the presynaptic side. Motor neurons, which is the common inhibitor in crab limb closers and limb accessory flexors, form axo-axonic synapses in addition to the neuromuscular junction with the muscles in crayfish.[46] These synapses were first observed in 1967,[6] when they were found to cause presynaptic inhibition in leg muscles of crayfish and crabs. Subsequent studies found that axo-axonic synapses showed varying numbers of occurrence based on the location of the leg muscles from the nervous system. For instance, proximal regions have thrice as many axo-axonic synapses than the central regions.[7] These synapses are proposed to function by limiting neurotransmitter release for controlled leg movements.[7]
Clinical Significance
An example of the physiological role of axo-axonic synapses, which are formed by GABAergic inhibitory interneurons to the axons of granule cells, is in eliciting spontaneous seizures, which is a key symptom of Intractable Epilepsy.[47] The presynaptic inhibitory interneurons, which can be labeled by cholecystokinin and GAT-1, are found to modulate the granule cells's spike output. The same cells subsequently project excitatory mossy fibers to pyramidal neurons in the hippocampal CA3 region.
One of the two leading theories for the pathoetiology of schizophrenia is the glutamate theory. Glutamate is a well studied neurotransmitter for its role in learning and memory, and also in the brain development during prenatal and childhood. Studies of rat striatum found inhibitory axo-axonic synapses formed on the glutamatergic cortico-striatal fibers.[33] They proposed that these axo-axonic synapses in the striatum could be responsible for inhibiting the glutamatergic neurons. Additionally, these dopaminergic synapses are also proposed to cause hyperdopaminergic activity and become neurotoxic for the postsynaptic glutamatergic neurons.[48] This mechanism is proposed to be a possible mechanism for glutamate dysfunction in observed schizophrenia.
Development
A study on the spinal cord in mice suggests that the sensory Ig/Caspr4 complex is involved in the formation of axo-axonic synapses on proprioceptive afferents. These synapses are formed through projection of GABAergic interneurons on sensory neurons, which is upstream to the motor neurons. In the axo-axonic synapse, expressing NB2 (Contactin5)/Caspr4 coreceptor complex in postsynaptic neurons along with expressing NrCAM/CHL1 in presynaptic interneurons results in the increased numbers of such synapses forming in the spinal cord.[49] Also, knocking out NB2 from the sensory neurons reduced the number of axo-axonic synapses from GABAergic interneurons, which suggests the necessity and the role of NB2 in synaptogenesis of axo-axonic type of synapses.[49][36]
References
- Palay SL (July 1956). "Synapses in the central nervous system". The Journal of Biophysical and Biochemical Cytology. 2 (4 Suppl): 193–202. doi:10.1083/jcb.2.4.193. PMC 2229686. PMID 13357542.
- Howard A, Tamas G, Soltesz I (June 2005). "Lighting the chandelier: new vistas for axo-axonic cells". Trends in Neurosciences. 28 (6): 310–6. doi:10.1016/j.tins.2005.04.004. PMID 15927687. S2CID 34853701.
- Buhl EH, Han ZS, Lörinczi Z, Stezhka VV, Karnup SV, Somogyi P (April 1994). "Physiological properties of anatomically identified axo-axonic cells in the rat hippocampus". Journal of Neurophysiology. 71 (4): 1289–307. doi:10.1152/jn.1994.71.4.1289. PMID 8035215.
- Somogyi P, Freund TF, Cowey A (1982-11-01). "The axo-axonic interneuron in the cerebral cortex of the rat, cat and monkey". Neuroscience. 7 (11): 2577–607. doi:10.1016/0306-4522(82)90086-0. PMID 7155343. S2CID 8693953.
- Gobel S (October 1971). "Axo-axonic septate junctions in the basket formations of the cat cerebellar cortex". The Journal of Cell Biology. 51 (1): 328–33. doi:10.1083/jcb.51.1.328. PMC 2108243. PMID 5165176.
- Atwood HL, Jones A (December 1967). "Presynaptic inhibition in crustacean muscle: axo-axonal synapse". Experientia. 23 (12): 1036–8. doi:10.1007/BF02136434. PMID 4294865. S2CID 27917444.
- Govind CK, Atwood HL, Pearce J (January 1995). "Inhibitory axoaxonal and neuromuscular synapses in the crayfish opener muscle: membrane definition and ultrastructure". The Journal of Comparative Neurology. 351 (3): 476–88. doi:10.1002/cne.903510313. PMID 7706554. S2CID 24022348.
- Cuntz H, Haag J, Forstner F, Segev I, Borst A (June 2007). "Robust coding of flow-field parameters by axo-axonal gap junctions between fly visual interneurons". Proceedings of the National Academy of Sciences of the United States of America. 104 (24): 10229–33. Bibcode:2007PNAS..10410229C. doi:10.1073/pnas.0703697104. PMC 1886000. PMID 17551009.
- Szabadics J, Varga C, Molnár G, Oláh S, Barzó P, Tamás G (January 2006). "Excitatory effect of GABAergic axo-axonic cells in cortical microcircuits". Science. 311 (5758): 233–5. Bibcode:2006Sci...311..233S. doi:10.1126/science.1121325. PMID 16410524. S2CID 40744562.
- Eccles JC (1961-12-01). "The mechanism of synaptic transmission". Ergebnisse der Physiologie, Biologischen Chemie und Experimentellen Pharmakologie. 51 (1): 299–430. doi:10.1007/BF02269100. PMID 13889060. S2CID 84195141.
- Alford S, Schwartz E (2009-01-01), "Presynaptic Inhibition", in Squire LR (ed.), Encyclopedia of Neuroscience, Academic Press, pp. 1001–1006, doi:10.1016/b978-008045046-9.00814-7, ISBN 978-0-08-045046-9
- Saladin KS (2011). Human anatomy (3rd ed.). New York: McGraw-Hill. ISBN 978-0-07-352560-0. OCLC 318191613.
- Principles of neural science. Kandel, Eric R., Schwartz, James H. (James Harris), 1932-2006., Jessell, Thomas M. (4th ed.). New York: McGraw-Hill, Health Professions Division. 2000. ISBN 0-8385-7701-6. OCLC 42073108.
{{cite book}}: CS1 maint: others (link) - Frank K (June 1959). "Basic Mechanisms of Synaptic Transmission in the Central Nervous System". IRE Transactions on Medical Electronics. ME-6 (2): 85–88. doi:10.1109/IRET-ME.1959.5007923.
- Gray EG (January 1962). "A morphological basis for pre-synaptic inhibition?". Nature. 193 (4810): 82–3. Bibcode:1962Natur.193...82G. doi:10.1038/193082a0. PMID 13901298. S2CID 4282950.
- Kidd M (April 1962). "Electron microscopy of the inner plexiform layer of the retina in the cat and the pigeon". Journal of Anatomy. 96 (Pt 2): 179–87. PMC 1244141. PMID 14455782.
- Colonnier M, Guillery RW (April 1964). "Synaptic organization in the lateral geniculate nucleus of the monkey". Zeitschrift für Zellforschung und Mikroskopische Anatomie. 62 (3): 333–55. doi:10.1007/BF00339284. PMID 14218147. S2CID 3173201.
- Hirata Y (February 1964). "Some Observations on the Fine Structure of the Synapses in the Olfactory Bulb of the Mouse, with Particular Reference to the Atypical Synaptic Configurations". Archivum Histologicum Japonicum = Nihon Soshikigaku Kiroku. 24 (3): 293–302. doi:10.1679/aohc1950.24.293. PMID 14133696.
- Gray EG, Young JZ (April 1964). "Electron Microscopy of Synaptic Structure of Octopus Brain". The Journal of Cell Biology. 21 (1): 87–103. doi:10.1083/jcb.21.1.87. PMC 2106419. PMID 14154498.
- Barron DH, Matthews BH (August 1935). "Intermittent conduction in the spinal cord". The Journal of Physiology. 85 (1): 73–103. doi:10.1113/jphysiol.1935.sp003303. PMC 1394492. PMID 16994699.
- Ramón y Cajal S (1909). Histologie du système nerveux de l'homme & des vertébrés. Paris: Maloine. doi:10.5962/bhl.title.48637.
- Andersen P, Eccles J, Voorhoeve PE (August 1963). "Inhibitory Synapses on Somas of Purkinje Cells in the Cerebellum". Nature. 199 (4894): 655–6. Bibcode:1963Natur.199..655A. doi:10.1038/199655a0. PMID 14074549. S2CID 39375166.
- Dizon MJ, Khodakhah K (July 2011). "The role of interneurons in shaping Purkinje cell responses in the cerebellar cortex". The Journal of Neuroscience. 31 (29): 10463–73. doi:10.1523/JNEUROSCI.1350-11.2011. PMC 3314287. PMID 21775592.
- Fairén A, Valverde F (December 1980). "A specialized type of neuron in the visual cortex of cat: a Golgi and electron microscope study of chandelier cells". The Journal of Comparative Neurology. 194 (4): 761–79. doi:10.1002/cne.901940405. PMID 7204642. S2CID 34356324.
- Lund JS, Boothe RG, Lund RD (November 1977). "Development of neurons in the visual cortex (area 17) of the monkey (Macaca nemestrina): a Golgi study from fetal day 127 to postnatal maturity". The Journal of Comparative Neurology. 176 (2): 149–88. doi:10.1002/cne.901760203. PMID 410850. S2CID 750242.
- Peters A, Proskauer CC, Ribak CE (April 1982). "Chandelier cells in rat visual cortex". The Journal of Comparative Neurology. 206 (4): 397–416. doi:10.1002/cne.902060408. PMID 7096634. S2CID 42805817.
- Somogyi P (November 1977). "A specific 'axo-axonal' interneuron in the visual cortex of the rat". Brain Research. 136 (2): 345–50. doi:10.1016/0006-8993(77)90808-3. PMID 922488. S2CID 7263036.
- Jones EG, Powell TP (September 1969). "Synapses on the axon hillocks and initial segments of pyramidal cell axons in the cerebral cortex". Journal of Cell Science. 5 (2): 495–507. doi:10.1242/jcs.5.2.495. PMID 5362338.
- Westrum LE (December 1975). "Electron microscopy of synaptic structures in olfactory cortex of early postnatal rats". Journal of Neurocytology. 4 (6): 713–32. doi:10.1007/BF01181632. PMID 1194932. S2CID 8710902.
- Wang X, Sun QQ (March 2012). "Characterization of axo-axonic synapses in the piriform cortex of Mus musculus". The Journal of Comparative Neurology. 520 (4): 832–47. doi:10.1002/cne.22792. PMC 3903392. PMID 22020781.
- Ganter P, Szücs P, Paulsen O, Somogyi P (2004). "Properties of horizontal axo-axonic cells in stratum oriens of the hippocampal CA1 area of rats in vitro". Hippocampus. 14 (2): 232–43. doi:10.1002/hipo.10170. PMID 15098728. S2CID 2993178.
- Peters A, Proskauer CC, Kaiserman-Abramof IR (December 1968). "The small pyramidal neuron of the rat cerebral cortex. The axon hillock and initial segment". The Journal of Cell Biology. 39 (3): 604–19. doi:10.1083/jcb.39.3.604. PMC 2107556. PMID 5699934.
- Kornhuber J, Kornhuber ME (1983). "Axo-axonic synapses in the rat striatum". European Neurology. 22 (6): 433–6. doi:10.1159/000115598. PMID 6662154.
- Dunn RC, Westrum LE, Dikmen SS (December 1977). "Axoaxonic synaptogenesis in neonatal kitten spinal trigeminal nucleus". Brain Research. 138 (3): 534–7. doi:10.1016/0006-8993(77)90689-8. PMID 616291. S2CID 27050299.
- Chiba T, Doba N (February 1976). "Catecholaminergic axo-axonic synapses in the nucleus of the tractus solitarius (pars commissuralis) of the cat: possible relation to presynaptic regulation of baroreceptor reflexes". Brain Research. 102 (2): 255–65. doi:10.1016/0006-8993(76)90881-7. PMID 1247885. S2CID 21966627.
- Goulding M, Bourane S, Garcia-Campmany L, Dalet A, Koch S (June 2014). "Inhibition downunder: an update from the spinal cord". Current Opinion in Neurobiology. SI: Inhibition: Synapses, Neurons and Circuits. 26: 161–6. doi:10.1016/j.conb.2014.03.006. PMC 4059017. PMID 24743058.
- Araki T, Otani T (September 1955). "Response of single motoneurons to direct stimulation in toad's spinal cord". Journal of Neurophysiology. 18 (5): 472–85. doi:10.1152/jn.1955.18.5.472. PMID 13252436.
- Coombs JS, Curtis DR, Eccles JC (December 1957). "The generation of impulses in motoneurones". The Journal of Physiology. 139 (2): 232–49. doi:10.1113/jphysiol.1957.sp005888. PMC 1358726. PMID 13492210.
- Zhu CG, Sandri C, Akert K (December 1981). "Morphological identification of axo-axonic and dendro-dendritic synapses in the rat substantia gelatinosa". Brain Research. 230 (1–2): 25–40. doi:10.1016/0006-8993(81)90389-9. PMID 7317779. S2CID 32854329.
- Conradi S (1968). "Axo-axonic synapses on cat spinal motoneurons". Acta Societatis Medicorum Upsaliensis. 73 (5–6): 239–42. PMID 5734435.
- Rudomin P (December 1990). "Presynaptic inhibition of muscle spindle and tendon organ afferents in the mammalian spinal cord". Trends in Neurosciences. 13 (12): 499–505. doi:10.1016/0166-2236(90)90084-N. PMID 1703681. S2CID 4050785.
- Sotelo C, Palay SL (February 1970). "The fine structure of the later vestibular nucleus in the rat. II. Synaptic organization". Brain Research. 18 (1): 93–115. doi:10.1016/0006-8993(70)90459-2. PMID 4313893.
- Robertson JD, Bodenheimer TS, Stage DE (October 1963). "The Ultrastructure of Mauthner Cell Synapses and Nodes in Goldfish Brains". The Journal of Cell Biology. 19 (1): 159–99. doi:10.1083/jcb.19.1.159. PMC 2106865. PMID 14069792.
- Nakajima Y (August 1974). "Fine structure of the synaptic endings on the Mauthner cell of the goldfish". The Journal of Comparative Neurology. 156 (4): 379–402. doi:10.1002/cne.901560402. PMID 4137668. S2CID 42394435.
- Bierman HS, Zottoli SJ, Hale ME (2009). "Evolution of the Mauthner axon cap". Brain, Behavior and Evolution. 73 (3): 174–87. doi:10.1159/000222562. PMID 19494486. S2CID 25637965.
- Pearce J, Govind CK (April 1993). "Reciprocal axo-axonal synapses between the common inhibitor and excitor motoneurons in crustacean limb muscles". Journal of Neurocytology. 22 (4): 259–65. doi:10.1007/BF01187124. PMID 8478645. S2CID 8541786.
- Sayin U, Osting S, Hagen J, Rutecki P, Sutula T (April 2003). "Spontaneous seizures and loss of axo-axonic and axo-somatic inhibition induced by repeated brief seizures in kindled rats". The Journal of Neuroscience. 23 (7): 2759–68. doi:10.1523/JNEUROSCI.23-07-02759.2003. PMC 6742074. PMID 12684462.
- Howes O, McCutcheon R, Stone J (February 2015). "Glutamate and dopamine in schizophrenia: an update for the 21st century". Journal of Psychopharmacology. 29 (2): 97–115. doi:10.1177/0269881114563634. PMC 4902122. PMID 25586400.
- Ashrafi S, Betley JN, Comer JD, Brenner-Morton S, Bar V, Shimoda Y, et al. (January 2014). "Neuronal Ig/Caspr recognition promotes the formation of axoaxonic synapses in mouse spinal cord". Neuron. 81 (1): 120–9. doi:10.1016/j.neuron.2013.10.060. PMC 3898991. PMID 24411736.
Further reading
- Alford S, Schwartz E (2009). "Presynaptic inhibition". Encyclopedia of Neuroscience. pp. 1001–1006. doi:10.1016/B978-008045046-9.00814-7. ISBN 9780080450469.
- Bennett MR (11 October 2001). History of the Synapse. CRC Press. ISBN 9781482284171.
- Eccles JC (22 October 2013). The Physiology of Synapses. Academic Press. ISBN 9781483226064.
- Kandel ER, ed. (2013). Principles of Neural Science (5th ed.). New York: McGraw-Hill.
- Roberts A, Bush BM, eds. (5 February 1981). Neurones without impulses: their significance for vertebrate and invertebrate nervous systems. Cambridge University Press.
- DeFelipe J, Jones EG (July 1992). "Santiago Ramón y Cajal and methods in neurohistology". Trends in Neurosciences. 15 (7): 237–46. doi:10.1016/0166-2236(92)90057-f. PMID 1381115. S2CID 44572164.
- Eccles JC (1961). "The mechanism of synaptic transmission". Ergebnisse der Physiologie, Biologischen Chemie und Experimentellen Pharmakologie. 51: 299–430. doi:10.1007/BF02269100. PMID 13889060. S2CID 84195141.
- Mozzachiodi R, Byrne JH (January 2010). "More than synaptic plasticity: role of nonsynaptic plasticity in learning and memory". Trends in Neurosciences. 33 (1): 17–26. doi:10.1016/j.tins.2009.10.001. PMC 2815214. PMID 19889466.
- Vitten H, Isaacson JS (September 2001). "Synaptic transmission: exciting times for presynaptic receptors". Current Biology. 11 (17): R695–7. doi:10.1016/s0960-9822(01)00411-0. PMID 11553342.
- Zhou ZJ, Lee S (September 2008). "Synaptic physiology of direction selectivity in the retina". The Journal of Physiology. 586 (18): 4371–6. doi:10.1113/jphysiol.2008.159020. PMC 2614022. PMID 18617561.