Zebrafish
The zebrafish (Danio rerio) is a freshwater fish belonging to the minnow family (Cyprinidae) of the order Cypriniformes. Native to India[2] and South Asia, it is a popular aquarium fish, frequently sold under the trade name zebra danio[3] (and thus often called a "tropical fish" although both tropical and subtropical). It is also found in private ponds.
| Danio rerio | |
|---|---|
 | |
| An adult female zebrafish | |
| Scientific classification | |
| Domain: | Eukaryota |
| Kingdom: | Animalia |
| Phylum: | Chordata |
| Class: | Actinopterygii |
| Order: | Cypriniformes |
| Family: | Cyprinidae |
| Subfamily: | Danioninae |
| Genus: | Danio |
| Species: | D. rerio |
| Binomial name | |
| Danio rerio (F. Hamilton, 1822) | |
| Synonyms | |
| |

The zebrafish is an important and widely used vertebrate model organism in scientific research. Zebrafish has been used for biomedicine and developmental biology. The species is used for studies, such as neurobehavioral phenomena. It is also used for psychological reasons such as abuse, cognitive, and affective disorders. The species are used to study and observe behavioral research.
Taxonomy
The zebrafish is a derived member of the genus Brachydanio, of the family Cyprinidae. It has a sister-group relationship with Danio aesculapii.[4] Zebrafish are also closely related to the genus Devario, as demonstrated by a phylogenetic tree of close species.[5]
Distribution
Range
The zebrafish is native to freshwater habitats in South Asia where it is found in India, Pakistan, Bangladesh, Nepal and Bhutan.[1][6][7][8] The northern limit is in the South Himalayas, ranging from the Sutlej river basin in the Pakistan–India border region to the state of Arunachal Pradesh in northeast Indian.[1][7] Its range is concentrated in the Ganges and Brahmaputra River basins, and the species was first described from Kosi River (lower Ganges basin) of India. Its range further south is more local, with scattered records from the Western and Eastern Ghats regions.[8][9][10] It has frequently been said to occur in Myanmar (Burma), but this is entirely based on pre-1930 records and likely refers to close relatives only described later, notably Danio kyathit.[8][11][12][13] Likewise, old records from Sri Lanka are highly questionable and remain unconfirmed.[11]
Zebrafish have been introduced to California, Connecticut, Florida and New Mexico in the United States, presumably by deliberate release by aquarists or by escape from fish farms. The New Mexico population had been extirpated by 2003 and it is unclear if the others survive, as the last published records were decades ago.[14] Elsewhere the species has been introduced to Colombia and Malaysia.[7][15]
Habitats
Zebrafish typically inhabit moderately flowing to stagnant clear water of quite shallow depth in streams, canals, ditches, oxbow lakes, ponds and rice paddies.[8][9][15][16] There is usually some vegetation, either submerged or overhanging from the banks, and the bottom is sandy, muddy or silty, often mixed with pebbles or gravel. In surveys of zebrafish locations throughout much of its Bangladeshi and Indian distribution, the water had a near-neutral to somewhat basic pH and mostly ranged from 16.5 to 34 °C (61.7–93.2 °F) in temperature.[8][9][17] One unusually cold site was only 12.3 °C (54.1 °F) and another unusually warm site was 38.6 °C (101.5 °F), but the zebrafish still appeared healthy. The unusually cold temperature was at one of the highest known zebrafish locations at 1,576 m (5,171 ft) above sea level, although the species has been recorded to 1,795 m (5,889 ft).[8][9]
Description
The zebrafish is named for the five uniform, pigmented, horizontal, blue stripes on the side of the body, which are reminiscent of a zebra's stripes, and which extend to the end of the caudal fin. Its shape is fusiform and laterally compressed, with its mouth directed upwards. The male is torpedo-shaped, with gold stripes between the blue stripes; the female has a larger, whitish belly and silver stripes instead of gold. Adult females exhibit a small genital papilla in front of the anal fin origin. The zebrafish can reach up to 4–5 cm (1.6–2.0 in) in length,[12] although they typically are 1.8–3.7 cm (0.7–1.5 in) in the wild with some variations depending on location.[9] Its lifespan in captivity is around two to three years, although in ideal conditions, this may be extended to over five years.[16][18] In the wild it is typically an annual species.[1]
Psychology
In 2015, a study was published about zebrafishes' capacity for episodic memory. The individuals showed a capacity to remember context with respect to objects, locations and occasions (what, when, where). Episodic memory is a capacity of explicit memory systems, typically associated with conscious experience.[19]
The Mauthner cells integrate a wide array of sensory stimuli to produce the escape reflex. Those stimuli are found to include the lateral line signals by McHenry et al. 2009 and visual signals consistent with looming objects by Temizer et al. 2015, Dunn et al. 2016, and Yao et al. 2016.[20]
Reproduction
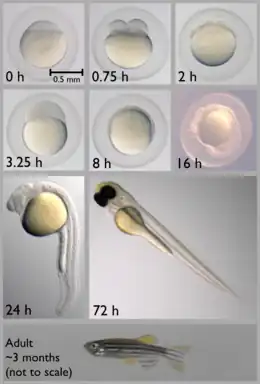
The approximate generation time for Danio rerio is three months. A male must be present for ovulation and spawning to occur. Zebrafish are asynchronous spawners[21] and under optimal conditions (such as food availability and favorable water parameters) can spawn successfully frequently, even on a daily basis.[22] Females are able to spawn at intervals of two to three days, laying hundreds of eggs in each clutch. Upon release, embryonic development begins; in absence of sperm, growth stops after the first few cell divisions. Fertilized eggs almost immediately become transparent, a characteristic that makes D. rerio a convenient research model species.[16] Sex determination of common laboratory strains was shown to be a complex genetic trait, rather than to follow a simple ZW or XY system.[23]
The zebrafish embryo develops rapidly, with precursors to all major organs appearing within 36 hours of fertilization. The embryo begins as a yolk with a single enormous cell on top (see image, 0 h panel), which divides into two (0.75 h panel) and continues dividing until there are thousands of small cells (3.25 h panel). The cells then migrate down the sides of the yolk (8 h panel) and begin forming a head and tail (16 h panel). The tail then grows and separates from the body (24 h panel). The yolk shrinks over time because the fish uses it for food as it matures during the first few days (72 h panel). After a few months, the adult fish reaches reproductive maturity (bottom panel).
To encourage the fish to spawn, some researchers use a fish tank with a sliding bottom insert, which reduces the depth of the pool to simulate the shore of a river. Zebrafish spawn best in the morning due to their Circadian rhythms. Researchers have been able to collect 10,000 embryos in 10 minutes using this method.[24] In particular, one pair of adult fish is capable of laying 200–300 eggs in one morning in approximately 5 to 10 at time.[25] Male zebrafish are furthermore known to respond to more pronounced markings on females, i.e., "good stripes", but in a group, males will mate with whichever females they can find. What attracts females is not currently understood. The presence of plants, even plastic plants, also apparently encourages spawning.[24]
Exposure to environmentally relevant concentrations of diisononyl phthalate (DINP), commonly used in a large variety of plastic items, disrupt the endocannabinoid system and thereby affect reproduction in a sex-specific manner.[26]
Feeding
Zebrafish are omnivorous, primarily eating zooplankton, phytoplankton, insects and insect larvae, although they can eat a variety of other foods, such as worms and small crustaceans, if their preferred food sources are not readily available.[16]
In research, adult zebrafish are often fed with brine shrimp, or paramecia.[27]
In the aquarium
Zebrafish are hardy fish and considered good for beginner aquarists. Their enduring popularity can be attributed to their playful disposition,[28] as well as their rapid breeding, aesthetics, cheap price and broad availability. They also do well in schools or shoals of six or more, and interact well with other fish species in the aquarium. However, they are susceptible to Oodinium or velvet disease, microsporidia (Pseudoloma neurophilia), and Mycobacterium species. Given the opportunity, adults eat hatchlings, which may be protected by separating the two groups with a net, breeding box or separate tank. In captivity, zebrafish live approximately forty-two months. Some captive zebrafish can develop a curved spine.[29]
The zebra danio was also used to make genetically modified fish and were the first species to be sold as GloFish (fluorescent colored fish).
Strains
In late 2003, transgenic zebrafish that express green, red, and yellow fluorescent proteins became commercially available in the United States. The fluorescent strains are tradenamed GloFish; other cultivated varieties include "golden", "sandy", "longfin" and "leopard".
The leopard danio, previously known as Danio frankei, is a spotted colour morph of the zebrafish which arose due to a pigment mutation.[30] Xanthistic forms of both the zebra and leopard pattern, along with long-finned strains, have been obtained via selective breeding programs for the aquarium trade.[31]
Various transgenic and mutant strains of zebrafish were stored at the China Zebrafish Resource Center (CZRC), a non-profit organization, which was jointly supported by the Ministry of Science and Technology of China and the Chinese Academy of Sciences.
Wild-type strains
The Zebrafish Information Network (ZFIN) provides up-to-date information about current known wild-type (WT) strains of D. rerio, some of which are listed below.[32]
- AB (AB)
- AB/C32 (AB/C32)
- AB/TL (AB/TL)
- AB/Tuebingen (AB/TU)
- C32 (C32)
- Cologne (KOLN)
- Darjeeling (DAR)
- Ekkwill (EKW)
- HK/AB (HK/AB)
- HK/Sing (HK/SING)
- Hong Kong (HK)
- India (IND)
- Indonesia (INDO)
- Nadia (NA)
- RIKEN WT (RW)
- Singapore (SING)
- SJA (SJA)
- SJD (SJD)
- SJD/C32 (SJD/C32)
- Tuebingen (TU)
- Tupfel long fin (TL)
- Tupfel long fin nacre (TLN)
- WIK (WIK)
- WIK/AB (WIK/AB)
Hybrids
Hybrids between different Danio species may be fertile: for example, between D. rerio and D. nigrofasciatus.[5]
Scientific research


D. rerio is a common and useful scientific model organism for studies of vertebrate development and gene function. Its use as a laboratory animal was pioneered by the American molecular biologist George Streisinger and his colleagues at the University of Oregon in the 1970s and 1980s; Streisinger's zebrafish clones were among the earliest successful vertebrate clones created.[33] Its importance has been consolidated by successful large-scale forward genetic screens (commonly referred to as the Tübingen/Boston screens). The fish has a dedicated online database of genetic, genomic, and developmental information, the Zebrafish Information Network (ZFIN). The Zebrafish International Resource Center (ZIRC) is a genetic resource repository with 29,250 alleles available for distribution to the research community. D. rerio is also one of the few fish species to have been sent into space.
Research with D. rerio has yielded advances in the fields of developmental biology, oncology,[34] toxicology,[25][35][36] reproductive studies, teratology, genetics, neurobiology, environmental sciences, stem cell research, regenerative medicine,[37][38] muscular dystrophies[39] and evolutionary theory.[5]
Model characteristics
As a model biological system, the zebrafish possesses numerous advantages for scientists. Its genome has been fully sequenced, and it has well-understood, easily observable and testable developmental behaviors. Its embryonic development is very rapid, and its embryos are relatively large, robust, and transparent, and able to develop outside their mother.[40] Furthermore, well-characterized mutant strains are readily available.
Other advantages include the species' nearly constant size during early development, which enables simple staining techniques to be used, and the fact that its two-celled embryo can be fused into a single cell to create a homozygous embryo. The zebrafish is also demonstrably similar to mammalian models and humans in toxicity testing, and exhibits a diurnal sleep cycle with similarities to mammalian sleep behavior.[41] However, zebrafish are not a universally ideal research model; there are a number of disadvantages to their scientific use, such as the absence of a standard diet[42] and the presence of small but important differences between zebrafish and mammals in the roles of some genes related to human disorders.[43][44]
Regeneration
Zebrafish have the ability to regenerate their heart and lateral line hair cells during their larval stages.[45][46] The cardiac regenerative process likely involves signaling pathways such as Notch and Wnt; hemodynamic changes in the damaged heart are sensed by ventricular endothelial cells and their associated cardiac cilia by way of the mechanosensitive ion channel TRPV4, subsequently facilitating the Notch signaling pathway via KLF2 and activating various downstream effectors such as BMP-2 and HER2/neu.[47] In 2011, the British Heart Foundation ran an advertising campaign publicising its intention to study the applicability of this ability to humans, stating that it aimed to raise £50 million in research funding.[48][49]
Zebrafish have also been found to regenerate photoreceptor cells and retinal neurons following injury, which has been shown to be mediated by the dedifferentiation and proliferation of Müller glia.[50] Researchers frequently amputate the dorsal and ventral tail fins and analyze their regrowth to test for mutations. It has been found that histone demethylation occurs at the site of the amputation, switching the zebrafish's cells to an "active", regenerative, stem cell-like state.[51][52] In 2012, Australian scientists published a study revealing that zebrafish use a specialised protein, known as fibroblast growth factor, to ensure their spinal cords heal without glial scarring after injury.[53][54] In addition, hair cells of the posterior lateral line have also been found to regenerate following damage or developmental disruption.[46][55] Study of gene expression during regeneration has allowed for the identification of several important signaling pathways involved in the process, such as Wnt signaling and Fibroblast growth factor.[55][56]
In probing disorders of the nervous system, including neurodegenerative diseases, movement disorders, psychiatric disorders and deafness, researchers are using the zebrafish to understand how the genetic defects underlying these conditions cause functional abnormalities in the human brain, spinal cord and sensory organs.[57][58][59][60] Researchers have also studied the zebrafish to gain new insights into the complexities of human musculoskeletal diseases, such as muscular dystrophy.[61] Another focus of zebrafish research is to understand how a gene called Hedgehog, a biological signal that underlies a number of human cancers, controls cell growth.
Background genetics
Inbred strains and traditional outbred stocks have not been developed for laboratory zebrafish, and the genetic variability of wild-type lines among institutions may contribute to the replication crisis in biomedical research.[62] Genetic differences in wild-type lines among populations maintained at different research institutions have been demonstrated using both Single-nucleotide polymorphisms[63] and microsatellite analysis.[64]
Gene expression
Due to their fast and short life cycles and relatively large clutch sizes, D. rerio or zebrafish are a useful model for genetic studies. A common reverse genetics technique is to reduce gene expression or modify splicing using Morpholino antisense technology. Morpholino oligonucleotides (MO) are stable, synthetic macromolecules that contain the same bases as DNA or RNA; by binding to complementary RNA sequences, they can reduce the expression of specific genes or block other processes from occurring on RNA. MO can be injected into one cell of an embryo after the 32-cell stage, reducing gene expression in only cells descended from that cell. However, cells in the early embryo (less than 32 cells) are interpermeable to large molecules,[65][66] allowing diffusion between cells. Guidelines for using Morpholinos in zebrafish describe appropriate control strategies.[67] Morpholinos are commonly microinjected in 500pL directly into 1-2 cell stage zebrafish embryos. The morpholino is able to integrate into most cells of the embryo.[68]
A known problem with gene knockdowns is that, because the genome underwent a duplication after the divergence of ray-finned fishes and lobe-finned fishes, it is not always easy to silence the activity of one of the two gene paralogs reliably due to complementation by the other paralog.[69] Despite the complications of the zebrafish genome, a number of commercially available global platforms exist for analysis of both gene expression by microarrays and promoter regulation using ChIP-on-chip.[70]
Genome sequencing
The Wellcome Trust Sanger Institute started the zebrafish genome sequencing project in 2001, and the full genome sequence of the Tuebingen reference strain is publicly available at the National Center for Biotechnology Information (NCBI)'s Zebrafish Genome Page. The zebrafish reference genome sequence is annotated as part of the Ensembl project, and is maintained by the Genome Reference Consortium.[71]
In 2009, researchers at the Institute of Genomics and Integrative Biology in Delhi, India, announced the sequencing of the genome of a wild zebrafish strain, containing an estimated 1.7 billion genetic letters.[72][73] The genome of the wild zebrafish was sequenced at 39-fold coverage. Comparative analysis with the zebrafish reference genome revealed over 5 million single nucleotide variations and over 1.6 million insertion deletion variations. The zebrafish reference genome sequence of 1.4GB and over 26,000 protein coding genes was published by Kerstin Howe et al. in 2013.[74]
Mitochondrial DNA
In October 2001, researchers from the University of Oklahoma published D. rerio's complete mitochondrial DNA sequence.[75] Its length is 16,596 base pairs. This is within 100 base pairs of other related species of fish, and it is notably only 18 pairs longer than the goldfish (Carassius auratus) and 21 longer than the carp (Cyprinus carpio). Its gene order and content are identical to the common vertebrate form of mitochondrial DNA. It contains 13 protein-coding genes and a noncoding control region containing the origin of replication for the heavy strand. In between a grouping of five tRNA genes, a sequence resembling vertebrate origin of light strand replication is found. It is difficult to draw evolutionary conclusions because it is difficult to determine whether base pair changes have adaptive significance via comparisons with other vertebrates' nucleotide sequences.[75]
Developmental genetics
T-boxes and homeoboxes are vital in Danio similarly to other vertebrates.[76][77] The Bruce et al. team are known for this area, and in Bruce et al. 2003 & Bruce et al. 2005 uncover the role of two of these elements in oocytes of this species.[76][77] By interfering via a dominant nonfunctional allele and a morpholino they find the T-box transcription activator Eomesodermin and its target mtx2 – a transcription factor – are vital to epiboly.[76][77] (In Bruce et al. 2003 they failed to support the possibility that Eomesodermin behaves like Vegt.[76] Neither they nor anyone else has been able to locate any mutation which – in the mother – will prevent initiation of the mesoderm or endoderm development processes in this species.)[76]
Pigmentation genes
In 1999, the nacre mutation was identified in the zebrafish ortholog of the mammalian MITF transcription factor.[78] Mutations in human MITF result in eye defects and loss of pigment, a type of Waardenburg Syndrome. In December 2005, a study of the golden strain identified the gene responsible for its unusual pigmentation as SLC24A5, a solute carrier that appeared to be required for melanin production, and confirmed its function with a Morpholino knockdown. The orthologous gene was then characterized in humans and a one base pair difference was found to strongly segregate fair-skinned Europeans and dark-skinned Africans.[79] Zebrafish with the nacre mutation have since been bred with fish with a roy orbison (roy) mutation to make Casper strain fish that have no melanophores or iridophores, and are transparent into adulthood. These fish are characterized by uniformly pigmented eyes and translucent skin.[80][81]
Transgenesis
Transgenesis is a popular approach to study the function of genes in zebrafish. Construction of transgenic zebrafish is rather easy by a method using the Tol2 transposon system. Tol2 element which encodes a gene for a fully functional transposase capable of catalyzing transposition in the zebrafish germ lineage. Tol2 is the only natural DNA transposable element in vertebrates from which an autonomous member has been identified.[82][83] Examples include the artificial interaction produced between LEF1 and Catenin beta-1/β-catenin/CTNNB1. Dorsky et al. 2002 investigated the developmental role of Wnt by transgenically expressing a Lef1/β-catenin reporter.[84]
There are well-established protocols for editing zebrafish genes using CRISPR-Cas9[85] and this tool has been used to generate genetically modified models.
Transparent adult bodies
In 2008, researchers at Boston Children's Hospital developed a new strain of zebrafish, named Casper, whose adult bodies had transparent skin.[80] This allows for detailed visualization of cellular activity, circulation, metastasis and many other phenomena.[80] In 2019 researchers published a crossing of a prkdc-/- and a IL2rga-/- strain that produced transparent, immunodeficient offspring, lacking natural killer cells as well as B- and T-cells. This strain can be adapted to 37 °C (99 °F) warm water and the absence of an immune system makes the use of patient derived xenografts possible.[86] In January 2013, Japanese scientists genetically modified a transparent zebrafish specimen to produce a visible glow during periods of intense brain activity.[87]
In January 2007, Chinese researchers at Fudan University genetically modified zebrafish to detect oestrogen pollution in lakes and rivers, which is linked to male infertility. The researchers cloned oestrogen-sensitive genes and injected them into the fertile eggs of zebrafish. The modified fish turned green if placed into water that was polluted by oestrogen.[88]
RNA splicing
In 2015, researchers at Brown University discovered that 10% of zebrafish genes do not need to rely on the U2AF2 protein to initiate RNA splicing. These genes have the DNA base pairs AC and TG as repeated sequences at the ends of each intron. On the 3'ss (3' splicing site), the base pairs adenine and cytosine alternate and repeat, and on the 5'ss (5' splicing site), their complements thymine and guanine alternate and repeat as well. They found that there was less reliance on U2AF2 protein than in humans, in which the protein is required for the splicing process to occur. The pattern of repeating base pairs around introns that alters RNA secondary structure was found in other teleosts, but not in tetrapods. This indicates that an evolutionary change in tetrapods may have led to humans relying on the U2AF2 protein for RNA splicing while these genes in zebrafish undergo splicing regardless of the presence of the protein.[89]
Orthology
D. rerio has three transferrins, all of which cluster closely with other vertebrates.[90]
Inbreeding depression
When close relatives mate, progeny may exhibit the detrimental effects of inbreeding depression. Inbreeding depression is predominantly caused by the homozygous expression of recessive deleterious alleles.[91] For zebrafish, inbreeding depression might be expected to be more severe in stressful environments, including those caused by anthropogenic pollution. Exposure of zebrafish to environmental stress induced by the chemical clotrimazole, an imidazole fungicide used in agriculture and in veterinary and human medicine, amplified the effects of inbreeding on key reproductive traits.[92] Embryo viability was significantly reduced in inbred exposed fish and there was a tendency for inbred males to sire fewer offspring.
Aquaculture research
Zebrafish are common models for research into fish farming, including pathogens[93][94][95] and parasites[93][95] causing yield loss and/or spread to adjacent wild populations.
This usefulness is less than it might be due to Danio's taxonomic distance from the most common aquaculture species.[94] Because the most common are salmonids and cod in the Protacanthopterygii and sea bass, sea bream, tilapia, and flatfish, in the Percomorpha, zebrafish results may not be perfectly applicable.[94] Various other models – Goldfish (Carassius auratus), Medaka (Oryzias latipes), Stickleback (Gasterosteus aculeatus), Roach (Rutilus rutilus), Pufferfish (Takifugu rubripes), Swordtail (Xiphophorus hellerii) – are less used normally but would be closer to particular target species.[95]
The only exception are the Carp (including Grass Carp, Ctenopharyngodon idella)[94] and Milkfish (Chanos chanos)[95] which are quite close, both being in the Cyprinidae. However it should also be noted that Danio consistently proves to be a useful model for mammals in many cases and there is dramatically more genetic distance between them than between Danio and any farmed fish.[94]
Neurochemistry
In a glucocorticoid receptor-defective mutant with reduced exploratory behavior, fluoxetine rescued the normal exploratory behavior.[96] This demonstrates relationships between glucocorticoids, fluoxetine, and exploration in this fish.[96]
Drug discovery and development
.jpg.webp)
The zebrafish and zebrafish larva is a suitable model organism for drug discovery and development. As a vertebrate with 70% genetic homology with humans,[74] it can be predictive of human health and disease, while its small size and fast development facilitates experiments on a larger and quicker scale than with more traditional in vivo studies, including the development of higher-throughput, automated investigative tools.[97][98] As demonstrated through ongoing research programmes, the zebrafish model enables researchers not only to identify genes that might underlie human disease, but also to develop novel therapeutic agents in drug discovery programmes.[99] Zebrafish embryos have proven to be a rapid, cost-efficient, and reliable teratology assay model.[100]
Drug screens
Drug screens in zebrafish can be used to identify novel classes of compounds with biological effects, or to repurpose existing drugs for novel uses; an example of the latter would be a screen which found that a commonly used statin (rosuvastatin) can suppress the growth of prostate cancer.[101] To date, 65 small-molecule screens have been carried out and at least one has led to clinical trials.[102] Within these screens, many technical challenges remain to be resolved, including differing rates of drug absorption resulting in levels of internal exposure that cannot be extrapolated from the water concentration, and high levels of natural variation between individual animals.[102]
Toxico- or pharmacokinetics
To understand drug effects, the internal drug exposure is essential, as this drives the pharmacological effect. Translating experimental results from zebrafish to higher vertebrates (like humans) requires concentration-effect relationships, which can be derived from pharmacokinetic and pharmacodynamic analysis.[103] Because of its small size, however, it is very challenging to quantify the internal drug exposure. Traditionally multiple blood samples would be drawn to characterize the drug concentration profile over time, but this technique remains to be developed. To date, only a single pharmacokinetic model for paracetamol has been developed in zebrafish larvae.[104]
Computational data analysis
Using smart data analysis methods, pathophysiological and pharmacological processes can be understood and subsequently translated to higher vertebrates, including humans.[103][105] An example is the use of systems pharmacology, which is the integration of systems biology and pharmacometrics. Systems biology characterizes (part of) an organism by a mathematical description of all relevant processes. These can be for example different signal transduction pathways that upon a specific signal lead to a certain response. By quantifying these processes, their behaviour in healthy and diseased situation can be understood and predicted. Pharmacometrics uses data from preclinical experiments and clinical trials to characterize the pharmacological processes that are underlying the relation between the drug dose and its response or clinical outcome. These can be for example the drug absorption in or clearance from the body, or its interaction with the target to achieve a certain effect. By quantifying these processes, their behaviour after different doses or in different patients can be understood and predicted to new doses or patients. By integrating these two fields, systems pharmacology has the potential to improve the understanding of the interaction of the drug with the biological system by mathematical quantification and subsequent prediction to new situations, like new drugs or new organisms or patients. Using these computational methods, the previously mentioned analysis of paracetamol internal exposure in zebrafish larvae showed reasonable correlation between paracetamol clearance in zebrafish with that of higher vertebrates, including humans.[104]
Medical research
Cancer
Zebrafish have been used to make several transgenic models of cancer, including melanoma, leukemia, pancreatic cancer and hepatocellular carcinoma.[106][107] Zebrafish expressing mutated forms of either the BRAF or NRAS oncogenes develop melanoma when placed onto a p53 deficient background. Histologically, these tumors strongly resemble the human disease, are fully transplantable, and exhibit large-scale genomic alterations. The BRAF melanoma model was utilized as a platform for two screens published in March 2011 in the journal Nature. In one study, the model was used as a tool to understand the functional importance of genes known to be amplified and overexpressed in human melanoma.[108] One gene, SETDB1, markedly accelerated tumor formation in the zebrafish system, demonstrating its importance as a new melanoma oncogene. This was particularly significant because SETDB1 is known to be involved in the epigenetic regulation that is increasingly appreciated to be central to tumor cell biology.
In another study, an effort was made to therapeutically target the genetic program present in the tumor's origin neural crest cell using a chemical screening approach.[109] This revealed that an inhibition of the DHODH protein (by a small molecule called leflunomide) prevented development of the neural crest stem cells which ultimately give rise to melanoma via interference with the process of transcriptional elongation. Because this approach would aim to target the "identity" of the melanoma cell rather than a single genetic mutation, leflunomide may have utility in treating human melanoma.[110]
Cardiovascular disease
In cardiovascular research, the zebrafish has been used to model human myocardial infarction model. The zebrafish heart completely regenerates after about 2 months of injury without any scar formation.[111] Zebrafish is also used as a model for blood clotting, blood vessel development, and congenital heart and kidney disease.[112]
Immune system
In programmes of research into acute inflammation, a major underpinning process in many diseases, researchers have established a zebrafish model of inflammation, and its resolution. This approach allows detailed study of the genetic controls of inflammation and the possibility of identifying potential new drugs.[113]
Zebrafish has been extensively used as a model organism to study vertebrate innate immunity. The innate immune system is capable of phagocytic activity by 28 to 30 h postfertilization (hpf)[114] while adaptive immunity is not functionally mature until at least 4 weeks postfertilization.[115]
Infectious diseases
As the immune system is relatively conserved between zebrafish and humans, many human infectious diseases can be modeled in zebrafish.[116][117][118][119] The transparent early life stages are well suited for in vivo imaging and genetic dissection of host-pathogen interactions.[120][121][122][123] Zebrafish models for a wide range of bacterial, viral and parasitic pathogens have already been established; for example, the zebrafish model for tuberculosis provides fundamental insights into the mechanisms of pathogenesis of mycobacteria.[124][125][126][127] Furthermore, robotic technology has been developed for high-throughput antimicrobial drug screening using zebrafish infection models.[128][129]
Repairing retinal damage

Another notable characteristic of the zebrafish is that it possesses four types of cone cell, with ultraviolet-sensitive cells supplementing the red, green and blue cone cell subtypes found in humans. Zebrafish can thus observe a very wide spectrum of colours. The species is also studied to better understand the development of the retina; in particular, how the cone cells of the retina become arranged into the so-called 'cone mosaic'. Zebrafish, in addition to certain other teleost fish, are particularly noted for having extreme precision of cone cell arrangement.[130]
This study of the zebrafish's retinal characteristics has also extrapolated into medical enquiry. In 2007, researchers at University College London grew a type of zebrafish adult stem cell found in the eyes of fish and mammals that develops into neurons in the retina. These could be injected into the eye to treat diseases that damage retinal neurons—nearly every disease of the eye, including macular degeneration, glaucoma, and diabetes-related blindness. The researchers studied Müller glial cells in the eyes of humans aged from 18 months to 91 years, and were able to develop them into all types of retinal neurons. They were also able to grow them easily in the lab. The stem cells successfully migrated into diseased rats' retinas, and took on the characteristics of the surrounding neurons. The team stated that they intended to develop the same approach in humans.[131][132]
Muscular dystrophies
Muscular dystrophies (MD) are a heterogeneous group of genetic disorders that cause muscle weakness, abnormal contractions and muscle wasting, often leading to premature death. Zebrafish is widely used as model organism to study muscular dystrophies.[39] For example, the sapje (sap) mutant is the zebrafish orthologue of human Duchenne muscular dystrophy (DMD).[133] The Machuca-Tzili and co-workers applied zebrafish to determine the role of alternative splicing factor, MBNL, in myotonic dystrophy type 1 (DM1) pathogenesis.[134] More recently, Todd et al. described a new zebrafish model designed to explore the impact of CUG repeat expression during early development in DM1 disease.[135] Zebrafish is also an excellent animal model to study congenital muscular dystrophies including CMD Type 1 A (CMD 1A) caused by mutation in the human laminin α2 (LAMA2) gene.[136] The zebrafish, because of its advantages discussed above, and in particular the ability of zebrafish embryos to absorb chemicals, has become a model of choice in screening and testing new drugs against muscular dystrophies.[137]
Bone physiology and pathology
Zebrafish have been used as model organisms for bone metabolism, tissue turnover, and resorbing activity. These processes are largely evolutionary conserved. They have been used to study osteogenesis (bone formation), evaluating differentiation, matrix deposition activity, and cross-talk of skeletal cells, to create and isolate mutants modeling human bone diseases, and test new chemical compounds for the ability to revert bone defects.[138][139] The larvae can be used to follow new (de novo) osteoblast formation during bone development. They start mineralising bone elements as early as 4 days post fertilisation. Recently, adult zebrafish are being used to study complex age related bone diseases such as osteoporosis and osteogenesis imperfecta.[140] The (elasmoid) scales of zebrafish function as a protective external layer and are little bony plates made by osteoblasts. These exoskeletal structures are formed by bone matrix depositing osteoblasts and are remodeled by osteoclasts. The scales also act as the main calcium storage of the fish. They can be cultured ex-vivo (kept alive outside of the organism) in a multi-well plate, which allows manipulation with drugs and even screening for new drugs that could change bone metabolism (between osteoblasts and osteoclasts).[140][141][142]
Diabetes
Zebrafish pancreas development is very homologous to mammals, such as mice. The signaling mechanisms and way the pancreas functions are very similar. The pancreas has an endocrine compartment, which contains a variety of cells. Pancreatic PP cells that produce polypeptides, and β-cells that produce insulin are two examples of those such cells. This structure of the pancreas, along with the glucose homeostasis system, are helpful in studying diseases, such as diabetes, that are related to the pancreas. Models for pancreas function, such as fluorescent staining of proteins, are useful in determining the processes of glucose homeostasis and the development of the pancreas. Glucose tolerance tests have been developed using zebrafish, and can now be used to test for glucose intolerance or diabetes in humans. The function of insulin are also being tested in zebrafish, which will further contribute to human medicine. The majority of work done surrounding knowledge on glucose homeostasis has come from work on zebrafish transferred to humans.[143]
Obesity
Zebrafish have been used as a model system to study obesity, with research into both genetic obesity and over-nutrition induced obesity. Obese zebrafish, similar to obese mammals, show dysregulation of lipid controlling metabolic pathways, which leads to weight gain without normal lipid metabolism.[143] Also like mammals, zebrafish store excess lipids in visceral, intramuscular, and subcutaneous adipose deposits. These reasons and others make zebrafish good models for studying obesity in humans and other species. Genetic obesity is usually studied in transgenic or mutated zebrafish with obesogenic genes. As an example, transgenic zebrafish with overexpressed AgRP, an endogenous melacortin antagonist, showed increased body weight and adipose deposition during growth.[143] Though zebrafish genes may not be the exact same as human genes, these tests could provide important insight into possible genetic causes and treatments for human genetic obesity.[143] Diet-induced obesity zebrafish models are useful, as diet can be modified from a very early age. High fat diets and general overfeeding diets both show rapid increases in adipose deposition, increased BMI, hepatosteatosis, and hypertriglyceridemia.[143] However, the normal fat, overfed specimens are still metabolically healthy, while high-fat diet specimens are not.[143] Understanding differences between types of feeding-induced obesity could prove useful in human treatment of obesity and related health conditions.[143]
Environmental toxicology
Zebrafish have been used as a model system in environmental toxicology studies.[25]
Epilepsy
Zebrafish have been used as a model system to study epilepsy. Mammalian seizures can be recapitulated molecularly, behaviorally, and electrophysiologically, using a fraction of the resources required for experiments in mammals.[144]
See also
- Japanese rice fish or medaka, another fish used for genetic, developmental, and biomedical research
- List of freshwater aquarium fish species
- Denison barb
References
- Vishwanath, W. (2010). "Danio rerio". IUCN Red List of Threatened Species. 2010: e.T166487A6219667. doi:10.2305/IUCN.UK.2010-4.RLTS.T166487A6219667.en. Retrieved 19 November 2021.
- Whiteley, Andrew R.; Bhat, Anuradha; Martins, Emilia P.; Mayden, Richard L.; Arunachalam, M.; Uusi‐Heikkilä, Silva; Ahmed, A. T. A.; Shrestha, Jiwan; Clark, Matthew; Stemple, Derek; Bernatchez, Louis (2011). "Population genomics of wild and laboratory zebrafish ( Danio rerio )". Molecular Ecology. 20 (20): 4259–4276. doi:10.1111/j.1365-294X.2011.05272.x. PMC 3627301. PMID 21923777.
- "Breeding Zebrafish (Zebra danios) - The fish doctor". www.thefishdoctor.co.uk. 16 November 2013.
- McCluskey BM, Postlethwait JH (March 2015). "Phylogeny of zebrafish, a "model species", within Danio, a "model genus"". Molecular Biology and Evolution. 32 (3): 635–652. doi:10.1093/molbev/msu325. PMC 4327152. PMID 25415969.
- Parichy DM (September 2006). "Evolution of danio pigment pattern development". Heredity. 97 (3): 200–210. doi:10.1038/sj.hdy.6800867. PMID 16835593.
- Petr, T. (1999). "Coldwater fish and fisheries in Bhutan". FAO. Retrieved 28 March 2019.
- Pritchard, V.L. (January 2001). Behavior and morphogy of the zebrafish, Danio rerio (PhD thesis). University of Leeds.
- Engeszer RE, Patterson LB, Rao AA, Parichy DM (2007). "Zebrafish in the wild: a review of natural history and new notes from the field". Zebrafish. 4 (1): 21–40. doi:10.1089/zeb.2006.9997. PMID 18041940. S2CID 34342799.
- Arunachalam M, Raja M, Vijayakumar C, Malaiammal P, Mayden RL (March 2013). "Natural history of zebrafish (Danio rerio) in India". Zebrafish. 10 (1): 1–14. doi:10.1089/zeb.2012.0803. PMID 23590398.
- "Zebrafish in the Natural Environment". University of Otago. Retrieved 28 March 2019.
- Spence, R. (December 2006), The behavior and ecology of the zebrafish, Danio rerio, University of Leicester
- "Brachydanio rerio (Hamilton, 1822)". SeriouslyFish. Retrieved 28 March 2019.
- Fang F (1998). "Danio kyathit, a new species of cyprinid species from Myitkyina, northern Myanmar". Ichthyological Exploration of Freshwaters. 8 (3): 273–280.
- "Danio rerio". Nonindigenous Aquatic Species. United States Geological Survey. June 14, 2013. Archived from the original on August 4, 2009. Retrieved July 3, 2013.
- Froese, Rainer; Pauly, Daniel (eds.) (2019). "Danio rerio" in FishBase. March 2019 version.
- Spence R, Gerlach G, Lawrence C, Smith C (February 2008). "The behaviour and ecology of the zebrafish, Danio rerio". Biological Reviews of the Cambridge Philosophical Society. 83 (1): 13–34. doi:10.1111/j.1469-185X.2007.00030.x. hdl:2381/27758. PMID 18093234. S2CID 18044956.
- Spence R, Fatema MK, Reichard M, Huq KA, Wahab MA, Ahmed ZF, Smith C (2006). "The distribution and habitat preferences of the zebrafish in Bangladesh". Journal of Fish Biology. 69 (5): 1435–1448. doi:10.1111/j.1095-8649.2006.01206.x.
- Gerhard GS, Kauffman EJ, Wang X, Stewart R, Moore JL, Kasales CJ, et al. (2002). "Life spans and senescent phenotypes in two strains of Zebrafish (Danio rerio)". Experimental Gerontology. 37 (8–9): 1055–1068. doi:10.1016/s0531-5565(02)00088-8. PMID 12213556. S2CID 25092240.
- Hamilton TJ, Myggland A, Duperreault E, May Z, Gallup J, Powell RA, et al. (November 2016). "Episodic-like memory in zebrafish". Animal Cognition. 19 (6): 1071–1079. doi:10.1007/s10071-016-1014-1. PMID 27421709. S2CID 2552608.
- Orger MB, de Polavieja GG (2017-07-25). "Zebrafish Behavior: Opportunities and Challenges". Annual Review of Neuroscience. 40 (1): 125–147. doi:10.1146/annurev-neuro-071714-033857. PMID 28375767. S2CID 46739494.
- Selman K, Wallace RA, Sarka A, Qi X (November 1993). "Stages of oocyte development in the zebrafish, Brachydanio rerio". Journal of Morphology. 218 (2): 203–224. doi:10.1002/jmor.1052180209. PMID 29865471. S2CID 46930941.
- Aleström P, D'Angelo L, Midtlyng PJ, Schorderet DF, Schulte-Merker S, Sohm F, Warner S (June 2020). "Zebrafish: Housing and husbandry recommendations". Laboratory Animals. 54 (3): 213–224. doi:10.1177/0023677219869037. PMC 7301644. PMID 31510859.
- Bradley K, Breyer P, Melville D, Broman K, Knapik E, Smith JR (Jun 2011). "A SNP-Based Linkage Map for Zebrafish Reveals Sex Determination Loci". G3 (Bethesda). 1 (1): 3–9. doi:10.1534/g3.111.000190. PMC 3178105. PMID 21949597. S2CID 11161125.
- Dockser A (January 13, 2012). "Birds Do It, Bees Do It, Even Zebrafish Do It—Just Too Little". Wall Street Journal. Retrieved February 11, 2012.
- Hill AJ, Teraoka H, Heideman W, Peterson RE (July 2005). "Zebrafish as a model vertebrate for investigating chemical toxicity". Toxicological Sciences. 86 (1): 6–19. doi:10.1093/toxsci/kfi110. PMID 15703261.
- Forner-Piquer I, Santangeli S, Maradonna F, Rabbito A, Piscitelli F, Habibi HR, et al. (October 2018). "Disruption of the gonadal endocannabinoid system in zebrafish exposed to diisononyl phthalate". Environmental Pollution. 241: 1–8. doi:10.1016/j.envpol.2018.05.007. PMID 29793103. S2CID 44120848.
- Westerfield M (2007). The Zebrafish Book: A Guide for the Laboratory Use of Zebrafish (Danio Rerio). University of Oregon Press.
- Gerhard GS, Cheng KC (December 2002). "A call to fins! Zebrafish as a gerontological model". Aging Cell. 1 (2): 104–111. doi:10.1046/j.1474-9728.2002.00012.x. PMID 12882339.
- "Zebrafish - Danio rerio - Details - Encyclopedia of Life". Encyclopedia of Life.
- Watanabe M, Iwashita M, Ishii M, Kurachi Y, Kawakami A, Kondo S, Okada N (September 2006). "Spot pattern of leopard Danio is caused by mutation in the zebrafish connexin41.8 gene". EMBO Reports. 7 (9): 893–897. doi:10.1038/sj.embor.7400757. PMC 1559663. PMID 16845369.
- Mills D (1993). Eyewitness Handbook: Aquarium Fish. Harper Collins. ISBN 978-0-7322-5012-6.
- "ZFIN: Wild-Type Lines: Summary Listing". zfin.org. Retrieved 2019-06-17.
- "In Memory of George Streisinger, "Founding Father" of Zebrafish Developmental and Genetic Research". University of Oregon. Archived from the original on September 29, 2015. Retrieved September 23, 2015.
- Xiang J, Yang H, Che C, Zou H, Yang H, Wei Y, et al. (2009). Isalan M (ed.). "Identifying tumor cell growth inhibitors by combinatorial chemistry and zebrafish assays". PLOS ONE. 4 (2): e4361. Bibcode:2009PLoSO...4.4361X. doi:10.1371/journal.pone.0004361. PMC 2633036. PMID 19194508.
- Bugel SM, Tanguay RL, Planchart A (September 2014). "Zebrafish: A marvel of high-throughput biology for 21st century toxicology". Current Environmental Health Reports. 1 (4): 341–352. doi:10.1007/s40572-014-0029-5. PMC 4321749. PMID 25678986.
- Dubińska-Magiera M, Daczewska M, Lewicka A, Migocka-Patrzałek M, Niedbalska-Tarnowska J, Jagla K (November 2016). "Zebrafish: A Model for the Study of Toxicants Affecting Muscle Development and Function". International Journal of Molecular Sciences. 17 (11): 1941. doi:10.3390/ijms17111941. PMC 5133936. PMID 27869769.
- Major RJ, Poss KD (2007). "Zebrafish Heart Regeneration as a Model for Cardiac Tissue Repair". Drug Discovery Today: Disease Models. 4 (4): 219–225. doi:10.1016/j.ddmod.2007.09.002. PMC 2597874. PMID 19081827.
- "Adult Stem Cell Research Avoids Ethical Concerns". Voice of America. 19 May 2010. Retrieved 21 June 2013.
- Plantié E, Migocka-Patrzałek M, Daczewska M, Jagla K (April 2015). "Model organisms in the fight against muscular dystrophy: lessons from drosophila and Zebrafish". Molecules. 20 (4): 6237–6253. doi:10.3390/molecules20046237. PMC 6272363. PMID 25859781.
- Dahm R (2006). "The Zebrafish Exposed". American Scientist. 94 (5): 446–53. doi:10.1511/2006.61.446. Archived from the original on 2017-04-18. Retrieved 2012-11-15.
- Jones R (October 2007). "Let sleeping zebrafish lie: a new model for sleep studies". PLOS Biology. 5 (10): e281. doi:10.1371/journal.pbio.0050281. PMC 2020498. PMID 20076649.
- Penglase S, Moren M, Hamre K (November 2012). "Lab animals: Standardize the diet for zebrafish model". Nature. 491 (7424): 333. Bibcode:2012Natur.491..333P. doi:10.1038/491333a. PMID 23151568.
- Jurynec MJ, Xia R, Mackrill JJ, Gunther D, Crawford T, Flanigan KM, et al. (August 2008). "Selenoprotein N is required for ryanodine receptor calcium release channel activity in human and zebrafish muscle". Proceedings of the National Academy of Sciences of the United States of America. 105 (34): 12485–12490. Bibcode:2008PNAS..10512485J. doi:10.1073/pnas.0806015105. PMC 2527938. PMID 18713863.
- Rederstorff M, Castets P, Arbogast S, Lainé J, Vassilopoulos S, Beuvin M, et al. (2011). "Increased muscle stress-sensitivity induced by selenoprotein N inactivation in mouse: a mammalian model for SEPN1-related myopathy". PLOS ONE. 6 (8): e23094. Bibcode:2011PLoSO...623094R. doi:10.1371/journal.pone.0023094. PMC 3152547. PMID 21858002.
- Wade, Nicholas (March 24, 2010). "Research Offers Clue Into How Hearts Can Regenerate in Some Species". The New York Times. Archived from the original on 2022-01-01.
- Lush ME, Piotrowski T (October 2014). "Sensory hair cell regeneration in the zebrafish lateral line". Developmental Dynamics. 243 (10): 1187–1202. doi:10.1002/dvdy.24167. PMC 4177345. PMID 25045019.
- Teske, Christopher. "An Evolving Role for Notch Signaling in Heart Regeneration of the Zebrafish Danio rerio". Researchgate.com. Retrieved 4 October 2022.
- "Mending Broken Hearts (2011) British Heart Foundation TV ad". British Heart Foundation via YouTube. January 31, 2011. Archived from the original on 2021-11-17. Retrieved November 15, 2012.
- "British Heart Foundation – The science behind the appeal". Bhf.org.uk. February 16, 2007. Archived from the original on 10 March 2012. Retrieved November 15, 2012.
- Bernardos RL, Barthel LK, Meyers JR, Raymond PA (June 2007). "Late-stage neuronal progenitors in the retina are radial Müller glia that function as retinal stem cells". The Journal of Neuroscience. 27 (26): 7028–7040. doi:10.1523/JNEUROSCI.1624-07.2007. PMC 6672216. PMID 17596452.
- Stewart S, Tsun ZY, Izpisua Belmonte JC (November 2009). "A histone demethylase is necessary for regeneration in zebrafish". Proceedings of the National Academy of Sciences of the United States of America. 106 (47): 19889–19894. Bibcode:2009PNAS..10619889S. doi:10.1073/pnas.0904132106. JSTOR 25593294. PMC 2785262. PMID 19897725.
- "Organ Regeneration In Zebrafish: Unraveling The Mechanisms". ScienceDaily.
- Goldshmit Y, Sztal TE, Jusuf PR, Hall TE, Nguyen-Chi M, Currie PD (May 2012). "Fgf-dependent glial cell bridges facilitate spinal cord regeneration in zebrafish". The Journal of Neuroscience. 32 (22): 7477–7492. doi:10.1523/JNEUROSCI.0758-12.2012. PMC 6703582. PMID 22649227.
- "Study Reveals Secret of Zebrafish | Biochemistry | Sci-News.com". Breaking Science News | Sci-News.com.
- Head JR, Gacioch L, Pennisi M, Meyers JR (July 2013). "Activation of canonical Wnt/β-catenin signaling stimulates proliferation in neuromasts in the zebrafish posterior lateral line". Developmental Dynamics. 242 (7): 832–846. doi:10.1002/dvdy.23973. PMID 23606225.
- Steiner AB, Kim T, Cabot V, Hudspeth AJ (April 2014). "Dynamic gene expression by putative hair-cell progenitors during regeneration in the zebrafish lateral line". Proceedings of the National Academy of Sciences of the United States of America. 111 (14): E1393–E1401. Bibcode:2014PNAS..111E1393S. doi:10.1073/pnas.1318692111. PMC 3986164. PMID 24706895.
- Kizil C (January 2018). "Mechanisms of Pathology-Induced Neural Stem Cell Plasticity and Neural Regeneration in Adult Zebrafish Brain". Current Pathobiology Reports. 6 (1): 71–77. doi:10.1007/s40139-018-0158-x. PMC 5978899. PMID 29938129.
- Cosacak MI, Bhattarai P, Reinhardt S, Petzold A, Dahl A, Zhang Y, Kizil C (April 2019). "Single-Cell Transcriptomics Analyses of Neural Stem Cell Heterogeneity and Contextual Plasticity in a Zebrafish Brain Model of Amyloid Toxicity". Cell Reports. 27 (4): 1307–1318.e3. doi:10.1016/j.celrep.2019.03.090. PMID 31018142.
- Bhattarai P, Cosacak MI, Mashkaryan V, Demir S, Popova SD, Govindarajan N, et al. (January 2020). "Neuron-glia interaction through Serotonin-BDNF-NGFR axis enables regenerative neurogenesis in Alzheimer's model of adult zebrafish brain". PLOS Biology. 18 (1): e3000585. doi:10.1371/journal.pbio.3000585. PMC 6964913. PMID 31905199.
- Xi Y, Noble S, Ekker M (June 2011). "Modeling neurodegeneration in zebrafish". Current Neurology and Neuroscience Reports. 11 (3): 274–282. doi:10.1007/s11910-011-0182-2. PMC 3075402. PMID 21271309.
- Bassett DI, Currie PD (October 2003). "The zebrafish as a model for muscular dystrophy and congenital myopathy". Human Molecular Genetics. 12 (Spec No 2): R265–R270. doi:10.1093/hmg/ddg279. PMID 14504264.
- Crim MJ, Lawrence C (January 2021). "A fish is not a mouse: understanding differences in background genetics is critical for reproducibility". Lab Animal. 50 (1): 19–25. doi:10.1038/s41684-020-00683-x. ISSN 0093-7355. PMID 33268901. S2CID 227259359.
- Whiteley AR, Bhat A, Martins EP, Mayden RL, Arunachalam M, Uusi-Heikkilä S, et al. (October 2011). "Population genomics of wild and laboratory zebrafish (Danio rerio)". Molecular Ecology. 20 (20): 4259–4276. doi:10.1111/j.1365-294X.2011.05272.x. PMC 3627301. PMID 21923777.
- Coe TS, Hamilton PB, Griffiths AM, Hodgson DJ, Wahab MA, Tyler CR (January 2009). "Genetic variation in strains of zebrafish (Danio rerio) and the implications for ecotoxicology studies". Ecotoxicology. 18 (1): 144–150. doi:10.1007/s10646-008-0267-0. PMID 18795247. S2CID 18370151.
- Kimmel CB, Law RD (March 1985). "Cell lineage of zebrafish blastomeres. I. Cleavage pattern and cytoplasmic bridges between cells". Developmental Biology. 108 (1): 78–85. doi:10.1016/0012-1606(85)90010-7. PMID 3972182.
- Kimmel CB, Law RD (March 1985). "Cell lineage of zebrafish blastomeres. III. Clonal analyses of the blastula and gastrula stages". Developmental Biology. 108 (1): 94–101. doi:10.1016/0012-1606(85)90012-0. PMID 3972184.
- Stainier DY, Raz E, Lawson ND, Ekker SC, Burdine RD, Eisen JS, et al. (October 2017). "Guidelines for morpholino use in zebrafish". PLOS Genetics. 13 (10): e1007000. doi:10.1371/journal.pgen.1007000. PMC 5648102. PMID 29049395.
- Rosen JN, Sweeney MF, Mably JD (March 2009). "Microinjection of zebrafish embryos to analyze gene function". Journal of Visualized Experiments (25). doi:10.3791/1115. PMC 2762901. PMID 19274045.
- Leong IU, Lan CC, Skinner JR, Shelling AN, Love DR (2012). "In vivo testing of microRNA-mediated gene knockdown in zebrafish". Journal of Biomedicine & Biotechnology. Hindawi. 2012: 350352. doi:10.1155/2012/350352. PMC 3303736. PMID 22500088.
- Tan PK, Downey TJ, Spitznagel EL, Xu P, Fu D, Dimitrov DS, et al. (October 2003). "Evaluation of gene expression measurements from commercial microarray platforms". Nucleic Acids Research. 31 (19): 5676–5684. doi:10.1093/nar/gkg763. PMC 206463. PMID 14500831.
- "Genome Reference Consortium". GRC. Retrieved October 23, 2012.
- "Decoding the Genome Mystery". Indian Express. July 5, 2009. Retrieved February 5, 2013.
- FishMap Zv8. Institute of Genomics and Integrative Biology (IGIB). Retrieved June 7, 2012.
- Howe K, Clark MD, Torroja CF, Torrance J, Berthelot C, Muffato M, et al. (April 2013). "The zebrafish reference genome sequence and its relationship to the human genome". Nature. 496 (7446): 498–503. Bibcode:2013Natur.496..498H. doi:10.1038/nature12111. PMC 3703927. PMID 23594743.
- Broughton RE, Milam JE, Roe BA (November 2001). "The complete sequence of the zebrafish (Danio rerio) mitochondrial genome and evolutionary patterns in vertebrate mitochondrial DNA". Genome Research. 11 (11): 1958–1967. doi:10.1101/gr.156801. PMC 311132. PMID 11691861.
- Schier, Alexander F.; Talbot, William S. (2005-12-01). "Molecular Genetics of Axis Formation in Zebrafish". Annual Review of Genetics. Annual Reviews. 39 (1): 561–613. doi:10.1146/annurev.genet.37.110801.143752. ISSN 0066-4197. PMID 16285872.
- Naiche, L.A.; Harrelson, Zachary; Kelly, Robert G.; Papaioannou, Virginia E. (2005-12-01). "T-Box Genes in Vertebrate Development". Annual Review of Genetics. Annual Reviews. 39 (1): 219–239. doi:10.1146/annurev.genet.39.073003.105925. ISSN 0066-4197. PMID 16285859.
- Lister JA, Robertson CP, Lepage T, Johnson SL, Raible DW (September 1999). "nacre encodes a zebrafish microphthalmia-related protein that regulates neural-crest-derived pigment cell fate". Development. 126 (17): 3757–3767. doi:10.1242/dev.126.17.3757. PMID 10433906.
- Lamason RL, Mohideen MA, Mest JR, Wong AC, Norton HL, Aros MC, et al. (December 2005). "SLC24A5, a putative cation exchanger, affects pigmentation in zebrafish and humans". Science. 310 (5755): 1782–1786. Bibcode:2005Sci...310.1782L. doi:10.1126/science.1116238. PMID 16357253. S2CID 2245002.
- White RM, Sessa A, Burke C, Bowman T, LeBlanc J, Ceol C, et al. (February 2008). "Transparent adult zebrafish as a tool for in vivo transplantation analysis". Cell Stem Cell. 2 (2): 183–189. doi:10.1016/j.stem.2007.11.002. PMC 2292119. PMID 18371439.
- Jeanna Bryner (February 6, 2008). "Scientists Create See-Through Fish, Watch Cancer Grow". livescience.com.
- Kawakami K, Takeda H, Kawakami N, Kobayashi M, Matsuda N, Mishina M (July 2004). "A transposon-mediated gene trap approach identifies developmentally regulated genes in zebrafish". Developmental Cell. 7 (1): 133–144. doi:10.1016/j.devcel.2004.06.005. PMID 15239961.
- Parinov S, Kondrichin I, Korzh V, Emelyanov A (October 2004). "Tol2 transposon-mediated enhancer trap to identify developmentally regulated zebrafish genes in vivo". Developmental Dynamics. 231 (2): 449–459. doi:10.1002/dvdy.20157. PMID 15366023.
- Barolo S, Posakony JW (May 2002). "Three habits of highly effective signaling pathways: principles of transcriptional control by developmental cell signaling". Genes & Development. Cold Spring Harbor Laboratory Press & The Genetics Society. 16 (10): 1167–1181. doi:10.1101/gad.976502. PMID 12023297. S2CID 14376483.
- Vejnar, Charles E.; Moreno-Mateos, Miguel A.; Cifuentes, Daniel; Bazzini, Ariel A.; Giraldez, Antonio J. (October 2016). "Optimized CRISPR–Cas9 System for Genome Editing in Zebrafish". Cold Spring Harbor Protocols. 2016 (10): pdb.prot086850. doi:10.1101/pdb.prot086850. ISSN 1940-3402. PMID 27698232.
- Yan C, Brunson DC, Tang Q, Do D, Iftimia NA, Moore JC, et al. (June 2019). "Visualizing Engrafted Human Cancer and Therapy Responses in Immunodeficient Zebrafish". Cell. 177 (7): 1903–1914.e14. doi:10.1016/j.cell.2019.04.004. PMC 6570580. PMID 31031007.
- "Researchers Capture A Zebrafish's Thought Process On Video". Popular Science. January 31, 2013. Retrieved February 4, 2013.
- "Fudan scientists turn fish into estrogen alerts". Xinhua. January 12, 2007. Retrieved November 15, 2012.
- Lin CL, Taggart AJ, Lim KH, Cygan KJ, Ferraris L, Creton R, et al. (January 2016). "RNA structure replaces the need for U2AF2 in splicing". Genome Research. 26 (1): 12–23. doi:10.1101/gr.181008.114. PMC 4691745. PMID 26566657.
- Gabaldón T, Koonin EV (May 2013). "Functional and evolutionary implications of gene orthology". Nature Reviews. Genetics. 14 (5): 360–366. doi:10.1038/nrg3456. PMC 5877793. PMID 23552219.
- Charlesworth D, Willis JH (November 2009). "The genetics of inbreeding depression". Nature Reviews. Genetics. 10 (11): 783–796. doi:10.1038/nrg2664. PMID 19834483. S2CID 771357.
- Bickley LK, Brown AR, Hosken DJ, Hamilton PB, Le Page G, Paull GC, et al. (February 2013). "Interactive effects of inbreeding and endocrine disruption on reproduction in a model laboratory fish". Evolutionary Applications. 6 (2): 279–289. doi:10.1111/j.1752-4571.2012.00288.x. PMC 3689353. PMID 23798977.
- Llewellyn, Martin S.; Boutin, Sébastien; Hoseinifar, Seyed Hossein; Derome, Nicolas (2014-06-02). "Teleost microbiomes: the state of the art in their characterization, manipulation and importance in aquaculture and fisheries". Frontiers in Microbiology. Frontiers. 5: 207. doi:10.3389/fmicb.2014.00207. ISSN 1664-302X. PMC 4040438. PMID 24917852. S2CID 13050990.
- Dahm, Ralf; Geisler, Robert (2006-04-25). "Learning from Small Fry: The Zebrafish as a Genetic Model Organism for Aquaculture Fish Species". Marine Biotechnology. European Society for Marine Biotechnology (ESMB) + Japanese Society for Marine Biotechnology (JSMB) + Australia New Zealand Marine Biotechnology Society (ANZMBS) (Springer). 8 (4): 329–345. doi:10.1007/s10126-006-5139-0. ISSN 1436-2228. PMID 16670967. S2CID 23994075.
- Ribas, Laia; Piferrer, Francesc (2013-07-31). "The zebrafish (Danio rerio) as a model organism, with emphasis on applications for finfish aquaculture research". Reviews in Aquaculture. Wiley. 6 (4): 209–240. doi:10.1111/raq.12041. ISSN 1753-5123. S2CID 84107971.
-
- • McCammon, Jasmine M.; Sive, Hazel (2015). "Addressing the Genetics of Human Mental Health Disorders in Model Organisms". Annual Review of Genomics and Human Genetics. Annual Reviews. 16 (1): 173–197. doi:10.1146/annurev-genom-090314-050048. ISSN 1527-8204. PMID 26002061. S2CID 19597664.
- • Ziv, L.; Muto, A.; Schoonheim, P. J.; Meijsing, S. H.; Strasser, D.; Ingraham, H. A.; Schaaf, M. J. M.; Yamamoto, K. R.; Baier, H. (2012). "An affective disorder in zebrafish with mutation of the glucocorticoid receptor". Molecular Psychiatry. Nature Portfolio/Macmillan Publishers Limited. 18 (6): 681–691. doi:10.1038/mp.2012.64. ISSN 1359-4184. PMC 4065652. PMID 22641177. S2CID 11962425. NIHMSID: NIHMS368312.
- Martin WK, Tennant AH, Conolly RB, Prince K, Stevens JS, DeMarini DM, et al. (January 2019). "High-Throughput Video Processing of Heart Rate Responses in Multiple Wild-type Embryonic Zebrafish per Imaging Field". Scientific Reports. 9 (1): 145. Bibcode:2019NatSR...9..145M. doi:10.1038/s41598-018-35949-5. PMC 6333808. PMID 30644404.
- Teixidó E, Kießling TR, Krupp E, Quevedo C, Muriana A, Scholz S (February 2019). "Automated Morphological Feature Assessment for Zebrafish Embryo Developmental Toxicity Screens". Toxicological Sciences. 167 (2): 438–449. doi:10.1093/toxsci/kfy250. PMC 6358258. PMID 30295906.
- "Fish for Science". University of Sheffield. 2011. Retrieved March 19, 2011.
- Brannen KC, Panzica-Kelly JM, Danberry TL, Augustine-Rauch KA (February 2010). "Development of a zebrafish embryo teratogenicity assay and quantitative prediction model". Birth Defects Research Part B: Developmental and Reproductive Toxicology. 89 (1): 66–77. doi:10.1002/bdrb.20223. PMID 20166227.
- Rennekamp AJ, Peterson RT (February 2015). "15 years of zebrafish chemical screening". Current Opinion in Chemical Biology. 24: 58–70. doi:10.1016/j.cbpa.2014.10.025. PMC 4339096. PMID 25461724.
- MacRae CA, Peterson RT (October 2015). "Zebrafish as tools for drug discovery". Nature Reviews. Drug Discovery. 14 (10): 721–731. doi:10.1038/nrd4627. PMID 26361349. S2CID 1979653.
- Van Wijk RC, Krekels EH, Hankemeier T, Spaink HP, Van der Graaf PH (2017). "Systems pharmacology of hepatic metabolism in zebrafish larvae". Drug Discovery Today: Disease Models. 22: 27–34. doi:10.1016/j.ddmod.2017.04.003.
- Kantae V, Krekels EH, Ordas A, González O, van Wijk RC, Harms AC, et al. (December 2016). "Pharmacokinetic Modeling of Paracetamol Uptake and Clearance in Zebrafish Larvae: Expanding the Allometric Scale in Vertebrates with Five Orders of Magnitude". Zebrafish. 13 (6): 504–510. doi:10.1089/zeb.2016.1313. PMC 5124745. PMID 27632065.
- Schulthess P, van Wijk RC, Krekels EH, Yates JW, Spaink HP, van der Graaf PH (May 2018). "Outside-In Systems Pharmacology Combines Innovative Computational Methods With High-Throughput Whole Vertebrate Studies". CPT: Pharmacometrics & Systems Pharmacology. 7 (5): 285–287. doi:10.1002/psp4.12297. PMC 5980533. PMID 29693322.
- Liu S, Leach SD (2011). "Zebrafish models for cancer". Annual Review of Pathology. 6: 71–93. doi:10.1146/annurev-pathol-011110-130330. PMID 21261518.
- "Zebrafish model of human melanoma reveals new cancer gene". Science Daily. March 23, 2011. Retrieved April 28, 2014.
- Ceol CJ, Houvras Y, Jane-Valbuena J, Bilodeau S, Orlando DA, Battisti V, et al. (March 2011). "The histone methyltransferase SETDB1 is recurrently amplified in melanoma and accelerates its onset". Nature. 471 (7339): 513–517. Bibcode:2011Natur.471..513C. doi:10.1038/nature09806. PMC 3348545. PMID 21430779.
- White RM, Cech J, Ratanasirintrawoot S, Lin CY, Rahl PB, Burke CJ, et al. (March 2011). "DHODH modulates transcriptional elongation in the neural crest and melanoma". Nature. 471 (7339): 518–522. Bibcode:2011Natur.471..518W. doi:10.1038/nature09882. PMC 3759979. PMID 21430780.
- "Arthritis Drug Could Help Beat Melanoma Skin Cancer, Study Finds". Sciencedaily. March 24, 2011. Retrieved November 15, 2012.
- Chablais, Fabian; Veit, Julia; Rainer, Gregor; Jaźwińska, Anna (2011). "The zebrafish heart regenerates after cryoinjury-induced myocardial infarction". BMC Developmental Biology. 11: 21. doi:10.1186/1471-213X-11-21. PMC 3078894. PMID 21473762.
- Drummond IA (February 2005). "Kidney development and disease in the zebrafish". Journal of the American Society of Nephrology. 16 (2): 299–304. doi:10.1681/ASN.2004090754. PMID 15647335. S2CID 25428361.
- "Investigating inflammatory disease using zebrafish". Fish For Science. Archived from the original on January 9, 2013. Retrieved November 15, 2012.
- Le Guyader D, Redd MJ, Colucci-Guyon E, Murayama E, Kissa K, Briolat V, et al. (January 2008). "Origins and unconventional behavior of neutrophils in developing zebrafish". Blood. 111 (1): 132–141. doi:10.1182/blood-2007-06-095398. PMID 17875807. S2CID 8853409.
- Novoa B, Figueras A (2012-01-01). "Zebrafish: Model for the Study of Inflammation and the Innate Immune Response to Infectious Diseases". In Lambris JD, Hajishengallis G (eds.). Current Topics in Innate Immunity II. Advances in Experimental Medicine and Biology. Vol. 946. Springer New York. pp. 253–275. doi:10.1007/978-1-4614-0106-3_15. hdl:10261/44975. ISBN 9781461401056. PMID 21948373. S2CID 6914876.
- Meeker ND, Trede NS (2008). "Immunology and zebrafish: spawning new models of human disease". Developmental and Comparative Immunology. 32 (7): 745–757. doi:10.1016/j.dci.2007.11.011. PMID 18222541.
- Renshaw SA, Trede NS (January 2012). "A model 450 million years in the making: zebrafish and vertebrate immunity". Disease Models & Mechanisms. 5 (1): 38–47. doi:10.1242/dmm.007138. PMC 3255542. PMID 22228790.
- Meijer AH, Spaink HP (June 2011). "Host-pathogen interactions made transparent with the zebrafish model". Current Drug Targets. 12 (7): 1000–1017. doi:10.2174/138945011795677809. PMC 3319919. PMID 21366518.
- van der Vaart M, Spaink HP, Meijer AH (2012). "Pathogen recognition and activation of the innate immune response in zebrafish". Advances in Hematology. 2012: 159807. doi:10.1155/2012/159807. PMC 3395205. PMID 22811714.
- Benard EL, van der Sar AM, Ellett F, Lieschke GJ, Spaink HP, Meijer AH (March 2012). "Infection of zebrafish embryos with intracellular bacterial pathogens". Journal of Visualized Experiments (61). doi:10.3791/3781. PMC 3415172. PMID 22453760.
- Meijer AH, van der Vaart M, Spaink HP (January 2014). "Real-time imaging and genetic dissection of host-microbe interactions in zebrafish". Cellular Microbiology. 16 (1): 39–49. doi:10.1111/cmi.12236. PMID 24188444.
- Torraca V, Masud S, Spaink HP, Meijer AH (July 2014). "Macrophage-pathogen interactions in infectious diseases: new therapeutic insights from the zebrafish host model". Disease Models & Mechanisms. 7 (7): 785–797. doi:10.1242/dmm.015594. PMC 4073269. PMID 24973749.
- Levraud JP, Palha N, Langevin C, Boudinot P (September 2014). "Through the looking glass: witnessing host-virus interplay in zebrafish". Trends in Microbiology. 22 (9): 490–497. doi:10.1016/j.tim.2014.04.014. PMID 24865811.
- Ramakrishnan L (2013). "Looking within the Zebrafish to Understand the Tuberculous Granuloma". The New Paradigm of Immunity to Tuberculosis. Advances in Experimental Medicine and Biology. Vol. 783. pp. 251–66. doi:10.1007/978-1-4614-6111-1_13. ISBN 978-1-4614-6110-4. PMID 23468113.
- Ramakrishnan L (2013). "The zebrafish guide to tuberculosis immunity and treatment". Cold Spring Harbor Symposia on Quantitative Biology. 78: 179–192. doi:10.1101/sqb.2013.78.023283. PMID 24643219.
- Cronan MR, Tobin DM (July 2014). "Fit for consumption: zebrafish as a model for tuberculosis". Disease Models & Mechanisms. 7 (7): 777–784. doi:10.1242/dmm.016089. PMC 4073268. PMID 24973748.
- Meijer AH (March 2016). "Protection and pathology in TB: learning from the zebrafish model". Seminars in Immunopathology. 38 (2): 261–273. doi:10.1007/s00281-015-0522-4. PMC 4779130. PMID 26324465.
- Spaink HP, Cui C, Wiweger MI, Jansen HJ, Veneman WJ, Marín-Juez R, et al. (August 2013). "Robotic injection of zebrafish embryos for high-throughput screening in disease models". Methods. 62 (3): 246–254. doi:10.1016/j.ymeth.2013.06.002. PMID 23769806.
- Veneman WJ, Marín-Juez R, de Sonneville J, Ordas A, Jong-Raadsen S, Meijer AH, Spaink HP (June 2014). "Establishment and optimization of a high throughput setup to study Staphylococcus epidermidis and Mycobacterium marinum infection as a model for drug discovery". Journal of Visualized Experiments. 88 (88): e51649. doi:10.3791/51649. PMC 4206090. PMID 24998295.
- Allison WT, Barthel LK, Skebo KM, Takechi M, Kawamura S, Raymond PA (October 2010). "Ontogeny of cone photoreceptor mosaics in zebrafish". The Journal of Comparative Neurology. 518 (20): 4182–4195. doi:10.1002/cne.22447. PMC 3376642. PMID 20878782.
- Lawrence JM, Singhal S, Bhatia B, Keegan DJ, Reh TA, Luthert PJ, et al. (August 2007). "MIO-M1 cells and similar muller glial cell lines derived from adult human retina exhibit neural stem cell characteristics". Stem Cells. 25 (8): 2033–2043. doi:10.1634/stemcells.2006-0724. PMID 17525239.
- "Zebra fish may point way to cure for blindness". The China Post. August 3, 2007. Archived from the original on 2012-09-10.
- Kunkel LM, Bachrach E, Bennett RR, Guyon J, Steffen L (May 2006). "Diagnosis and cell-based therapy for Duchenne muscular dystrophy in humans, mice, and zebrafish". Journal of Human Genetics. 51 (5): 397–406. doi:10.1007/s10038-006-0374-9. PMC 3518425. PMID 16583129.
- Machuca-Tzili LE, Buxton S, Thorpe A, Timson CM, Wigmore P, Luther PK, Brook JD (May 2011). "Zebrafish deficient for Muscleblind-like 2 exhibit features of myotonic dystrophy". Disease Models & Mechanisms. 4 (3): 381–392. doi:10.1242/dmm.004150. PMC 3097459. PMID 21303839.
- Todd PK, Ackall FY, Hur J, Sharma K, Paulson HL, Dowling JJ (January 2014). "Transcriptional changes and developmental abnormalities in a zebrafish model of myotonic dystrophy type 1". Disease Models & Mechanisms. 7 (1): 143–155. doi:10.1242/dmm.012427. PMC 3882056. PMID 24092878.
- Jones KJ, Morgan G, Johnston H, Tobias V, Ouvrier RA, Wilkinson I, North KN (October 2001). "The expanding phenotype of laminin alpha2 chain (merosin) abnormalities: case series and review". Journal of Medical Genetics. 38 (10): 649–657. doi:10.1136/jmg.38.10.649. PMC 1734735. PMID 11584042.
- Maves L (September 2014). "Recent advances using zebrafish animal models for muscle disease drug discovery". Expert Opinion on Drug Discovery. 9 (9): 1033–1045. doi:10.1517/17460441.2014.927435. PMC 4697731. PMID 24931439.
- Witten PE, Hansen A, Hall BK (December 2001). "Features of mono- and multinucleated bone resorbing cells of the zebrafish Danio rerio and their contribution to skeletal development, remodeling, and growth". Journal of Morphology. 250 (3): 197–207. doi:10.1002/jmor.1065. PMID 11746460. S2CID 33403358.
- Carnovali M, Banfi G, Mariotti M (2019). "Zebrafish Models of Human Skeletal Disorders: Embryo and Adult Swimming Together". BioMed Research International. 2019: 1253710. doi:10.1155/2019/1253710. PMC 6886339. PMID 31828085.
- Bergen DJ, Kague E, Hammond CL (2019). "Zebrafish as an Emerging Model for Osteoporosis: A Primary Testing Platform for Screening New Osteo-Active Compounds". Frontiers in Endocrinology. 10: 6. doi:10.3389/fendo.2019.00006. PMC 6361756. PMID 30761080.
- de Vrieze E, van Kessel MA, Peters HM, Spanings FA, Flik G, Metz JR (February 2014). "Prednisolone induces osteoporosis-like phenotype in regenerating zebrafish scales". Osteoporosis International. 25 (2): 567–578. doi:10.1007/s00198-013-2441-3. PMID 23903952. S2CID 21829206.
- de Vrieze E, Zethof J, Schulte-Merker S, Flik G, Metz JR (May 2015). "Identification of novel osteogenic compounds by an ex-vivo sp7:luciferase zebrafish scale assay". Bone. 74: 106–113. doi:10.1016/j.bone.2015.01.006. PMID 25600250.
- Zang L, Maddison LA, Chen W (20 August 2018). "Zebrafish as a Model for Obesity and Diabetes". Frontiers in Cell and Developmental Biology. 6 (91): 91. doi:10.3389/fcell.2018.00091. PMC 6110173. PMID 30177968.
- Cho SJ, Park E, Baker A, Reid AY (2020-09-10). "Age Bias in Zebrafish Models of Epilepsy: What Can We Learn From Old Fish?". Frontiers in Cell and Developmental Biology. 8: 573303. doi:10.3389/fcell.2020.573303. PMC 7511771. PMID 33015065.
Further reading
- Kalueff AV, Stewart AM, Kyzar EJ, Cachat J, Gebhardt M, Landsman S, et al. (International Zebrafish Neuroscience Research Consortium (ZNRC)) (2012). "Time to recognize zebrafish "affective" behavior". Behaviour. 149 (10/12): 1019–1036. JSTOR 41720603.
- Lambert DJ (1997). Freshwater Aquarium Fish. Edison, New Jersey: Chartwell Books. p. 19. ISBN 978-0-7858-0867-1.
- Sharpe S. "Zebra Danio". Your Guide to Freshwater Aquariums. Retrieved December 15, 2004.
- Kocher TD, Jeffery WR, Parichy DM, Peichel CL, Streelman JT, Thorgaard GH (2005). "Fish models for studying adaptive evolution and speciation". Zebrafish. 2 (3): 147–156. doi:10.1089/zeb.2005.2.147. PMID 18248189. S2CID 18940475.
- Bradbury J (May 2004). "Small fish, big science". PLOS Biology. 2 (5): E148. doi:10.1371/journal.pbio.0020148. PMC 406403. PMID 15138510.
- Westerfield M (2007). The Zebrafish Book: A guide for the laboratory use of zebrafish (Danio rerio) (5th ed.). Eugene, OR: University of Oregon Press. ASIN B003KFCWKS.
- Guttridge N (2012). "Targeted gene modification can rewrite zebrafish DNA". Nature. doi:10.1038/nature.2012.11463. S2CID 87708919.
- "A Point Of View: Fly, Fish, Mouse and Worm". BBC News. June 14, 2013. Retrieved June 15, 2013.
External links
- British Association of Zebrafish Husbandry Archived 2018-05-19 at the Wayback Machine
- International Zebrafish Society (IZFS)
- European Society for Fish Models in Biology and Medicine (EuFishBioMed)
- The Zebrafish Information Network (ZFIN)
- The Zebrafish International Resource Center (ZIRC)
- The European Zebrafish Resource Center (EZRC)
- The China Zebrafish Resource Center (CZRC)
- The Zebrafish Genome Sequencing Project at the Wellcome Trust Sanger Institute
- FishMap: The Zebrafish Community Genomics Browser at the Institute of Genomics and Integrative Biology (IGIB)
- WebHome Zebrafish GenomeWiki Beta Preview at the IGIB
- Genome sequencing initiative at the IGIB
- Danio rerio at Danios.info
- Sanger Institute Zebrafish Mutation Resource
- Zebrafish genome via Ensembl
- FishforScience.com – using zebrafish for medical research
- FishForPharma
- Breeding Zebrafish
- View the danRer10 genome assembly in the UCSC Genome Browser.