Diel vertical migration
Diel vertical migration (DVM), also known as diurnal vertical migration, is a pattern of movement used by some organisms, such as copepods, living in the ocean and in lakes. The word "diel" (IPA: /ˈdaɪ.əl/, /ˈdiː.əl/) comes from Latin: diēs, lit. 'day', and means a 24-hour period. The migration occurs when organisms move up to the uppermost layer of the sea at night and return to the bottom of the daylight zone of the oceans or to the dense, bottom layer of lakes during the day.[2] It is important to the functioning of deep-sea food webs and the biologically driven sequestration of carbon.[3]

and the ocean surface – animation by NASA [1]
In terms of biomass, it is the largest synchronous migration in the world.[4][2] It is not restricted to any one taxon as examples are known from crustaceans (copepods),[5] molluscs (squid),[6] and ray-finned fishes (trout).[7]
The phenomenon may be advantageous for a number of reasons, most typically to access food and avoid predators.[8] It is triggered by various stimuli, the most prominent being response to changes in light intensity,[8] though evidence suggests that biological clocks are an underlying stimulus as well.[9] While this mass migration is generally nocturnal, with the animals ascending from the depths at nightfall and descending at sunrise, the timing can be altered in response to the different cues and stimuli that trigger it. Some unusual events impact vertical migration: DVM can be absent during the midnight sun in Arctic regions[10][11] and vertical migration can occur suddenly during a solar eclipse.[12] The phenomenon also demonstrates cloud-driven variations.[13]
The common swift is an exception among birds in that it ascends and descends into high altitudes at dusk and dawn, similar to the vertical migration of aquatic lifeforms.
Discovery
The phenomenon was first documented by French naturalist Georges Cuvier in 1817. He noted that daphnia, a type of plankton, appeared and disappeared according to a diurnal pattern.[4][14]
During World War II the U.S. Navy was taking sonar readings of the ocean when they discovered the deep scattering layer (DSL). While performing sound propagation experiments, the University of California's Division of War Research (UCDWR) consistently had results of the echo-sounder that showed a distinct reverberation that they attributed to mid-water layer scattering agents. At the time, there was speculation that these readings may be attributed to enemy submarines.[15]
Martin W. Johnson of Scripps Institution of Oceanography proposed a possible explanation. Working with the UCDWR, the Scripps researchers were able to confirm that the observed reverberations from the echo-sounder were in fact related to the diel vertical migration of marine animals. The DSL was caused by large, dense groupings of organisms, like zooplankton, that scattered the sonar to create a false or second bottom.[4][14][15]
Once scientists started to do more research on what was causing the DSL, it was discovered that a large range of organisms were vertically migrating. Most types of plankton and some types of nekton have exhibited some type of vertical migration, although it is not always diel. These migrations may have substantial effects on mesopredators and apex predators by modulating the concentration and accessibility of their prey (e.g., impacts on the foraging behavior of pinnipeds[16]).
Types of vertical migration
Diel
This is the most common form of vertical migration. Organisms migrate on a daily basis through different depths in the water column. Migration usually occurs between shallow surface waters of the epipelagic zone and deeper mesopelagic zone of the ocean or hypolimnion zone of lakes.[2] There are three recognized types of diel vertical migration:
Nocturnal vertical migration
In the most common form, nocturnal vertical migration, organisms ascend to the surface around dusk, remaining at the surface for the night, then migrating to depth again around dawn.[8]
Reverse migration
Reverse migration occurs with organisms ascending to the surface at sunrise and remaining high in the water column throughout the day until descending with the setting sun. [8]
Twilight diel vertical migration
Twilight diel vertical migration involves two separate migrations in a single 24-hour period, with the first ascent at dusk followed by a descent at midnight, often known as the "midnight sink". The second ascent to the surface and descent to the depths occurs at sunrise.[8]
Seasonal
Organisms are found at different depths depending on what season it is.[17] Seasonal changes to the environment may influence changes to migration patterns. Normal diel vertical migration occurs in species of foraminifera throughout the year in the polar regions; however, during the midnight sun, no differential light cues exist so they remain at the surface to feed upon the abundant phytoplankton, or to facilitate photosynthesis by their symbionts.[11] This is not true for all species at all times, however. Zooplankton have been observed to resynchronize their migrations with the light of the moon during periods when the sun is not visible, and to stay in deeper waters when the moon is full.[4]
Larger seasonally-migrating zooplankton such as overwintering copepods have been shown to transport a substantial amount of carbon to the deep ocean through a process known as the lipid pump.[18] The lipid pump is a process that sequesters carbon (in the form of carbon-rich lipids) out of the surface ocean via the descent of copepods to the deep during autumn.[18] These copepods accumulate these lipids during late summer and autumn before descending to the deep to overwinter in response to reduced primary production and harsh conditions at the surface.[18][19] Furthermore, they rely on these lipid reserves that are metabolized for energy to survive through winter before ascending back to the surface in the spring, typically at the onset of a spring bloom.[18]
Ontogenetic
Organisms spend different stages of their life cycle at different depths.[20] There are often pronounced differences in migration patterns of adult female copepods, like Eurytemora affinis, which stay at depth with only a small upward movement at night, compared to the rest of its life stages which migrate over 10 meters. In addition, there is a trend seen in other copepods, like Acartia spp. that have an increasing amplitude of their DVM seen with their progressive life stages. This is possibly due to increasing body size of the copepods and the associated risk of visual predators, like fish, as being larger makes them more noticeable.[5]
Vertical migration stimuli
There are two different types of factors that are known to play a role in vertical migration, endogenous and exogenous. Endogenous factors originate from the organism itself; sex, age, size, biological rhythms, etc. Exogenous factors are environmental factors acting on the organism such as light, gravity, oxygen, temperature, predator-prey interactions, etc.[21]
Endogenous rhythm
Biological clocks are an ancient and adaptive sense of time innate to an organism that allows them to anticipate environmental changes and cycles so they are able to physiologically and behaviorally respond to the expected change.[9]
Evidence of circadian rhythms controlling DVM, metabolism, and even gene expression have been found in copepod species, Calanus finmarchicus. These copepods were shown to continue to exhibit these daily rhythms of vertical migration in the laboratory setting even in constant darkness, after being captured from an actively migrating wild population.[9]
An experiment was done at the Scripps Institution of Oceanography which kept organisms in column tanks with light/dark cycles. A few days later the light was changed to a constant low light and the organisms still displayed diel vertical migration. This suggests that some type of internal response was causing the migration.[22]
Clock gene expression
Many organisms, including the copepod C. finmarchicus, have genetic material devoted to maintaining their biological clock. The expression of these genes varies temporally with the expression significantly increasing following dawn and dusk at times of greatest vertical migration. These findings may indicate they work as a molecular stimulus for vertical migration.[9]
Body size
The relative body size of an organism has been found to affect DVM. Bull trout express daily and seasonal vertical migrations with smaller individuals always staying at a deeper layer than the larger individuals. This is most likely due to a predation risk, but is dependent on the individuals own size such that smaller animals may be more inclined to remain at depth.[7]
Light
"Light is the most common and critical cue for vertical migration".[8] However, as of 2010, there had not been sufficient research to determine which aspect of the light field was responsible.[8] As of 2020, research has suggested that both light intensity and spectral composition of light are important.[23]
Temperature
Organisms will migrate to a water depth with temperatures that best suit the organisms needs, for example some fish species migrate to warmer surface waters in order to aid digestion. Temperature changes can influence swimming behavior of some copepods. In the presence of a strong thermocline some zooplankton may be inclined to pass through it, and migrate to the surface waters, though this can be very variable even in a single species. The marine copepod, Calanus finmarchicus, will migrate through gradients with temperature differences of 6 °C over George's Bank; whereas, in the North Sea they are observed to remain below the gradient.[24]
Salinity
Changes in salinity may promote organism to seek out more suitable waters if they happen to be stenohaline or unequipped to handle regulating their osmotic pressure. Areas that are impacted by tidal cycles accompanied by salinity changes, estuaries for example, may see vertical migration in some species of zooplankton.[25] Salinity has also been proposed as a factor that regulates the biogeochemical impact of diel vertical migration.[26]
Pressure
Pressure changes have been found to produce differential responses that result in vertical migration. Many zooplankton will react to increased pressure with positive phototaxis, a negative geotaxis, and/or a kinetic response that results in ascending in the water column. Likewise, when there is a decrease in pressure, the zoo plankton respond by passively sinking or active downward swimming to descend in the water column.[25]
Predator kairomones
A predator might release a chemical cue which could cause its prey to vertically migrate away.[27] This may stimulate the prey to vertically migrate to avoid said predator. The introduction of a potential predator species, like a fish, to the habitat of diel vertical migrating zooplankton has been shown to influence the distribution patterns seen in their migration. For example, a study used Daphnia and a fish that was too small to prey of them (Lebistus reticulatus), found that with the introduction of the fish to the system the Daphnia remained below the thermocline, where the fish was not present. This demonstrates the effects of kairomones on Daphnia DVM.[24]
Tidal patterns
Some organisms have been found to move with the tidal cycle. A study looked at the abundance of a species of small shrimp, Acetes sibogae, and found that they tended to move further higher in the water column and in higher numbers during flood tides than during ebb tides experiences at the mouth of an estuary. It is possible that varying factors with the tides may be the true trigger for the migration rather than the movement of the water itself, like the salinity or minute pressure changes.[25]
Reasons for vertical migration
There are many hypotheses as to why organisms would vertically migrate, and several may be valid at any given time.[28]
Predator avoidance
The universality of DVM suggests that there is some powerful common factor behind it. The connection between available light and DVM has led researchers to theorize that organisms may stay in deeper, darker areas during the day to avoid being eaten by predators who depend on light to see and catch their prey. While the ocean's surface provides an abundance of food, it may be safest for many species to visit it at night.[4]
Light-dependent predation by fish is a common pressure that causes DVM behavior in zooplankton and krill. A given body of water may be viewed as a risk gradient whereby the surface layers are riskier to reside in during the day than deep water, and as such promotes varied longevity among zooplankton that settle at different daytime depths.[29] Indeed, in many instances it is advantageous for zooplankton to migrate to deep waters during the day to avoid predation and come up to the surface at night to feed. For example, the northern krill Meganyctiphanes norvegica undergoes diel vertical migration to avoid planktivorous fish.[30]
Patterns among migrators seem to support the predator avoidance theory. Migrators will stay in groups as they migrate, a behavior that may protect individuals within the group from being eaten. Groups of smaller, harder to see animals begin their upward migration before larger, easier to see species, consistent with the idea that detectability by visual predators is a key issue. Small creatures may start to migrate upwards as much as 20 minutes before the sun sets, while large conspicuous fish may wait as long as 80 minutes after the sun goes down. Species that are better able to avoid predators also tend to migrate before those with poorer swimming capabilities. Squid are a primary prey for Risso's dolphins (Grampus griseus), an air-breathing predator, but one that relies on acoustic rather than visual information to hunt. Squid delay their migration pattern by about 40 minutes when dolphins are about, lessening risk by feeding later and for a shorter time.[4][31]
Metabolic advantages
Another possibility is that predators can benefit from diel vertical migration as an energy conservation strategy. Studies indicate that male dogfish (Scyliorhinus canicula) follow a "hunt warm - rest cool" strategy that enables them to lower their daily energy costs. They remain in warm water only long enough to obtain food, and then return to cooler areas where their metabolism can operate more slowly. [31][32][33]
Alternatively, organisms feeding on the bottom in cold water during the day may migrate to surface waters at night in order to digest their meal at warmer temperatures.
Dispersal and transport
Organisms can use deep and shallow currents to find food patches or to maintain a geographical location.
Avoid UV damage
The sunlight can penetrate into the water column. If an organism, especially something small like a microbe, is too close to the surface the UV can damage them. So they would want to avoid getting too close to the surface, especially during daylight.[34][35]
Water transparency
A theory known as the “transparency-regulator hypothesis" predicts that "the relative roles of UV and visual predation pressure will vary systematically across a gradient of lake transparency."[34][35] In less transparent waters, where fish are present and more food is available, fish tend to be the main driver of DVM. In more transparent bodies of water, where fish are less numerous and food quality improves in deeper waters, UV light can travel farther, thus functioning as the main driver of DVM in such cases.[36]
Unusual events
Due to the particular types of stimuli and cues used to initiate vertical migration, anomalies can change the pattern drastically.
For example, the occurrence of midnight sun in the Arctic induces changes to planktonic life that would normally perform DVM with a 24-hour night and day cycle. In the summers of the Arctic the Earth's north pole is directed toward the sun creating longer days and at the high latitude continuous day light for more than 24-hours.[10] Species of foraminifera found in the ocean have been observed to cease their DVM pattern, and rather remain at the surface in favor of feeding on the phytoplankton.[37] For example Neogloboquadrina pachyderma, and for those species that contain symbionts, like Turborotalita quinqueloba, remain in sunlight to aid photosynthesis.[11] Changes in sea-ice and surface chlorophyll concentration are found to be stronger determinants of the vertical habitat of Arctic N. pachyderma.[37]
There is also evidence of changes to vertical migration patterns during solar eclipse events. In the moments that the sun is obscured during normal day light hours, there is a sudden dramatic decrease in light intensity. The decreased light intensity, replicates the typical lighting experienced at night time that stimulate the planktonic organisms to migrate. During an eclipse, some copepod species distribution is concentrated near the surface, for example Calanus finmarchicus displays a classic diurnal migration pattern but on a much shorter time scale during an eclipse.[12]
Importance for the biological pump
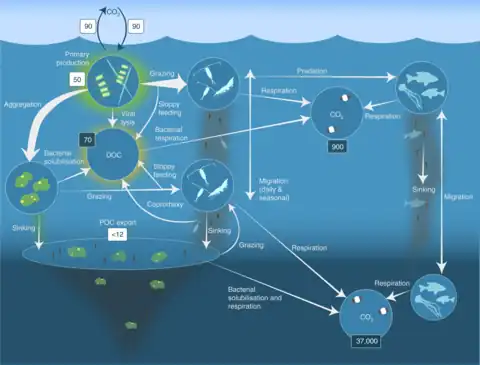
The biological pump is the conversion of CO2 and inorganic nutrients by plant photosynthesis into particulate organic matter in the euphotic zone and transference to the deeper ocean.[39] This is a major process in the ocean and without vertical migration it wouldn't be nearly as efficient. The deep ocean gets most of its nutrients from the higher water column when they sink down in the form of marine snow. This is made up of dead or dying animals and microbes, fecal matter, sand and other inorganic material.
Organisms migrate up to feed at night so when they migrate back to depth during the day they defecate large sinking fecal pellets.[39] Whilst some larger fecal pellets can sink quite fast, the speed that organisms move back to depth is still faster. At night organisms are in the top 100 metres of the water column, but during the day they move down to between 800 and 1000 meters. If organisms were to defecate at the surface it would take the fecal pellets days to reach the depth that they reach in a matter of hours. Therefore, by releasing fecal pellets at depth they have almost 1000 metres less to travel to get to the deep ocean. This is known as active transport. The organisms are playing a more active role in moving organic matter down to depths. Because a large majority of the deep sea, especially marine microbes, depends on nutrients falling down, the quicker they can reach the ocean floor the better.
Zooplankton and salps play a large role in the active transport of fecal pellets. 15–50% of zooplankton biomass is estimated to migrate, accounting for the transport of 5–45% of particulate organic nitrogen to depth.[39] Salps are large gelatinous plankton that can vertically migrate 800 meters and eat large amounts of food at the surface. They have a very long gut retention time, so fecal pellets usually are released at maximum depth. Salps are also known for having some of the largest fecal pellets. Because of this they have a very fast sinking rate, small detritus particles are known to aggregate on them. This makes them sink that much faster. As previously mentioned, the lipid pump represents a substantial flux of POC to the deep ocean in the form of lipids produced by large overwintering copepods.[18] Through overwintering, these lipids are transported to the deep in autumn and are metabolized at depths below the thermocline through winter before the copepods rise to the surface in the spring.[18] The metabolism of these lipids reduces this POC at depth while producing CO2 as a waste product, ultimately serving as a potentially significant contributor to oceanic carbon sequestration.[18] Although the flux of lipid carbon from the lipid pump has been reported to be comparable to the global POC flux from the biological pump, observational challenges with the lipid pump from deficient nutrient cycling,[40][41][42] and capture techniques have made it difficult to incorporate it into the global carbon export flux.[19][43][44] So while currently there is still much research being done on why organisms vertically migrate, it is clear that vertical migration plays a large role in the active transport of dissolved organic matter to depth.[45]
See also
References
- This twilight zone is dark, watery, and yes, also full of intrigue NASA Blog, 21 August 2018.
- "Diel Vertical Migration (DVM)". 2011-08-24. Retrieved 5 April 2012.
- Drazen, Jeffrey C.; Sutton, Tracey T. (3 January 2017). "Dining in the Deep: The Feeding Ecology of Deep-Sea Fishes". Annual Review of Marine Science. 9 (1): 337–366. Bibcode:2017ARMS....9..337D. doi:10.1146/annurev-marine-010816-060543. ISSN 1941-1405. PMID 27814034. Retrieved 21 December 2021.
- Seo, Hannah (3 December 2021). "The ups and downs of a great vertical migration". Knowable Magazine. doi:10.1146/knowable-110421-1. Retrieved 11 December 2021.
- P.B. Holliland, I. Ahlbeck, E. Westlund, S. Hansson (April 2012). "Ontogenetic and seasonal changes in diel vertical migration amplitude of the calanoid copepods Eurytemora affinis and Acartia spp. in a coastal area of the northern Baltic proper". Journal of Plankton Research. 34 (4): 298–307. doi:10.1093/plankt/fbs001.
{{cite journal}}: CS1 maint: multiple names: authors list (link) - R. Rosa, B.A. Seibel (July–August 2010). "Metabolic physiology of the Humboldt squid, Dosidicus gigas: Implications for vertical migration in a pronounced oxygen minimum zone". Progress in Oceanography. 86 (1–2): 72–80. Bibcode:2010PrOce..86...72R. doi:10.1016/j.pocean.2010.04.004.
- L.F.G. Gutowsky, P.M. Harrison, E.G. Martins, A. Leake, D.A. Patterson, M. Power, S.J. Cooke (August 2013). "Diel vertical migration hypotheses explain size-dependent behaviour in freshwater piscivore". Animal Behaviour. 86 (2): 365–373. doi:10.1016/j.anbehav.2013.05.027. S2CID 49544030.
{{cite journal}}: CS1 maint: multiple names: authors list (link) - B. Cisewski, V.H. Strass, M. Rhein, S. Kragefsky (January 2010). "Seasonal variation of diel vertical migration of zooplankton from ADCP backscatter time series data in the Lazarev Sea, Antarctica" (PDF). Deep Sea Research Part I: Oceanographic Research Papers. 57 (1): 78–94. Bibcode:2010DSRI...57...78C. doi:10.1016/j.dsr.2009.10.005.
{{cite journal}}: CS1 maint: multiple names: authors list (link) - N.S. Hafker, B. Meyer, K.S. Last, D.W. Pond, L. Huppe, M. Taschke (July 2017). "Circadian clock involvement in zooplankton diel vertical migration". Current Biology. 27 (14): 2194–2201. doi:10.1016/j.cub.2017.06.025. PMID 28712565.
{{cite journal}}: CS1 maint: multiple names: authors list (link) - Darnis, G.; Hobbs, L.; Geoffroy, M.; Grenvald, J. C.; Renaud, P. E.; Berge, J.; Cottier, F.; Kristiansen, S.; Daase, M.; Søreide, J. E.; Wold, A.; Morata, N.; Gabrielsen, T. (2017). "From polar night to midnight sun: Diel vertical migration, metabolism and biogeochemical role of zooplankton in a high Arctic fjord (Kongsfjorden, Svalbard)". Limnology and Oceanography. 62 (4): 1586–1605. Bibcode:2017LimOc..62.1586D. doi:10.1002/lno.10519. ISSN 1939-5590. S2CID 56252159.
- C. Manno, A.K. Pavlov (January 2014). "Living planktonic foraminifera in the Fram Strait (Arctic): absence of diel vertical migration during the midnight sun". Hydrobiologia. 721: 285–295. doi:10.1007/s10750-013-1669-4. S2CID 18320084.
- K. Sherman, K.A. Honey (May 1970). "Vertical movements of zooplankton during a solar eclipse". Nature. 227 (5263): 1156–1158. Bibcode:1970Natur.227.1156S. doi:10.1038/2271156a0. PMID 5451119. S2CID 4209244.
- Omand, Melissa M.; Steinberg, Deborah K.; Stamieszkin, Karen (10 August 2021). "Cloud shadows drive vertical migrations of deep-dwelling marine life". Proceedings of the National Academy of Sciences. 118 (32): e2022977118. Bibcode:2021PNAS..11822977O. doi:10.1073/pnas.2022977118. ISSN 0027-8424. PMC 8364114. PMID 34349017.
- Bandara, Kanchana; Varpe, Øystein; Wijewardene, Lishani; Tverberg, Vigdis; Eiane, Ketil (2021). "Two hundred years of zooplankton vertical migration research". Biological Reviews. 96 (4): 1547–1589. doi:10.1111/brv.12715. hdl:10037/22091. ISSN 1469-185X. PMID 33942990. S2CID 233722666. Retrieved 11 December 2021.
- Hill, M.N. (2005). Physical Oceanography. Harvard University Press. p. 499.
- Horning, M.; Trillmich, F. (1999). "Lunar cycles in diel prey migrations exert a stronger effect on the diving of juveniles than adult Galapagos fur seals". Proceedings of the Royal Society B. 266 (1424): 1127–1132. doi:10.1098/rspb.1999.0753. PMC 1689955. PMID 10406130.
- Visser, Andre; Sigrun Jonasdottir (1999). "Lipids, buoyancy and the seasonal vertical migration of Calanus finmarchicus". Fisheries Oceanography. 8: 100–106. doi:10.1046/j.1365-2419.1999.00001.x.
- Jónasdóttir, Sigrún Huld; Visser, André W.; Richardson, Katherine; Heath, Michael R. (2015-09-29). "Seasonal copepod lipid pump promotes carbon sequestration in the deep North Atlantic". Proceedings of the National Academy of Sciences. 112 (39): 12122–12126. doi:10.1073/pnas.1512110112. ISSN 0027-8424. PMC 4593097. PMID 26338976.
- Visser, Andre W.; Grønning, Josephine; Jónasdóttir, Sigrún Huld (2017). "Calanus hyperboreus and the lipid pump". Limnology and Oceanography. 62 (3): 1155–1165. Bibcode:2017LimOc..62.1155V. doi:10.1002/lno.10492. ISSN 1939-5590. S2CID 51989153.
- Kobari, Toru; Tsutomu Ikeda (2001). "Octogenetic vertical migration and life cycle of Neocalanus plumchrus (Crustacea:Copepoda) in the Oyashio region, with notes on regional variations in body size". Journal of Plankton Research. 23 (3): 287–302. doi:10.1093/plankt/23.3.287.
- Richards, Shane; Hugh Possingham; John Noye (1996). "Diel vertical migration: modeling light-mediated mechanisms". Journal of Plankton Research. 18 (12): 2199–2222. doi:10.1093/plankt/18.12.2199.
- Enright, J.T.; W.M. Hammer (1967). "Vertical Diurnal Migration and Endogenous Rhythmicity". Science. 157 (3791): 937–941. Bibcode:1967Sci...157..937E. doi:10.1126/science.157.3791.937. JSTOR 1722121. PMID 17792830. S2CID 20415350.
- Prins, Antoine; Deleris, Paul; Hubas, Cédric; Jesus, Bruno (2020). "Effect of Light Intensity and Light Quality on Diatom Behavioral and Physiological Photoprotection". Frontiers in Marine Science. 7: 203. doi:10.3389/fmars.2020.00203. ISSN 2296-7745.
- Ringelberg, Joop (2010). Diel Vertical Migration of Zooplankton in Lakes and Oceans. London New York, USA: Springer Science. pp. 122–127. ISBN 978-90-481-3092-4.
- Barnes, Margaret (1993). Oceanography and Marine Biology, An Annual Review. Vol. 31. Boca Raton, Florida, USA: CRC Press. pp. 327–329.
- Darnis, G.; Hobbs, L.; Geoffroy, M.; Grenvald, J. C.; Renaud, P. E.; Berge, J.; Cottier, F.; Kristiansen, S.; Daase, M.; E. Søreide, J.; Wold, A.; Morata, N.; Gabrielsen, T. (2017). "From polar night to midnight sun: Diel vertical migration, metabolism and biogeochemical role of zooplankton in a high Arctic fjord (Kongsfjorden, Svalbard)". Limnology and Oceanography. 62 (4): 1586–1605. Bibcode:2017LimOc..62.1586D. doi:10.1002/lno.10519. ISSN 0024-3590.
- von Elert, Eric; Georg Pohnert (2000). "Diel Predator specificity of kairomones in diel vertical migration of Daphnia: a chemical approach". OIKOS. 88 (1): 119–128. doi:10.1034/j.1600-0706.2000.880114.x. ISSN 0030-1299.
- Kerfoot, WC (1985). "Adaptive value of vertical migration: Comments on the predation hypothesis and some alternatives". Contributions in Marine Science. 27: 91–113. Archived from the original on 2013-01-21.
- Dawidowicz, Piotr; Prędki, Piotr; Pietrzak, Barbara (2012-11-23). "Depth-selection behavior and longevity in Daphnia: an evolutionary test for the predation-avoidance hypothesis". Hydrobiologia. 715 (1): 87–91. doi:10.1007/s10750-012-1393-5. ISSN 0018-8158.
- Onsrud, M.S.R.; Kaartvedt, S.; Røstad, A.; Klevjer, T.A. (2004-01-01). "Vertical distribution and feeding patterns in fish foraging on the krill Meganyctiphanes norvegica". ICES Journal of Marine Science. 61 (8): 1278–1290. doi:10.1016/j.icesjms.2004.09.005. ISSN 1095-9289.
- Benoit-Bird, Kelly J.; Moline, Mark A. (2021). "Vertical migration timing illuminates the importance of visual and nonvisual predation pressure in the mesopelagic zone". Limnology and Oceanography. 66 (8): 3010–3019. Bibcode:2021LimOc..66.3010B. doi:10.1002/lno.11855. ISSN 1939-5590. S2CID 236232448. Retrieved 13 December 2021.
- Sims, David W.; Wearmouth, Victoria J.; Southall, Emily J.; Hill, Jacqueline M.; Moore, Pippa; Rawlinson, Kate; Hutchinson, Neil; Budd, Georgina C.; Righton, David; Metcalfe, Julian D.; Nash, Jon P.; Morritt, David (January 2006). "Hunt warm, rest cool: bioenergetic strategy underlying diel vertical migration of a benthic shark". Journal of Animal Ecology. 75 (1): 176–190. doi:10.1111/j.1365-2656.2005.01033.x. ISSN 0021-8790. PMID 16903055. S2CID 39013753.
- Sogard, Susan M.; Olla, Bori L. (1996). "Food deprivation affects vertical distribution and activity of a marine fish in a thermal gradient: potential energy-conserving mechanisms" (PDF). Marine Ecology Progress Series. 133: 43–55. Bibcode:1996MEPS..133...43S. doi:10.3354/meps133043. Retrieved 20 December 2021.
- Leach, Taylor H.; Williamson, Craig E.; Theodore, Nora; Fischer, Janet M.; Olson, Mark H. (3 August 2015). "The role of ultraviolet radiation in the diel vertical migration of zooplankton: an experimental test of the transparency-regulator hypothesis". Journal of Plankton Research. 37 (5): 886–896. doi:10.1093/plankt/fbv061. ISSN 0142-7873.
- Williamson, Craig E.; Fischer, Janet M.; Bollens, Stephen M.; Overholt, Erin P.; Breckenridge, Joanne K. (September 2011). "Toward a more comprehensive theory of zooplankton diel vertical migration: Integrating ultraviolet radiation and water transparency into the biotic paradigm" (PDF). Limnology and Oceanography. 56 (5): 1603–1623. Bibcode:2011LimOc..56.1603W. doi:10.4319/lo.2011.56.5.1603. S2CID 16088708. Retrieved 21 December 2021.
- Tiberti, Rocco; Iacobuzio, Rocco (2012-12-09). "Does the fish presence influence the diurnal vertical distribution of zooplankton in high transparency lakes?". Hydrobiologia. 709 (1): 27–39. doi:10.1007/s10750-012-1405-5. ISSN 0018-8158. S2CID 18757663.
- Greco, Mattia; Jonkers, Lukas; Kretschmer, Kerstin; Bijma, Jelle; Kucera, Michal (12 September 2019). "Depth habitat of the planktonic foraminifera Neogloboquadrina pachyderma in the northern high latitudes explained by sea-ice and chlorophyll concentrations". Biogeosciences. 16 (17): 3425–3437. Bibcode:2019BGeo...16.3425G. doi:10.5194/bg-16-3425-2019. ISSN 1726-4170. S2CID 203072987.
- Cavan, E.L., Belcher, A., Atkinson, A., Hill, S.L., Kawaguchi, S., McCormack, S., Meyer, B., Nicol, S., Ratnarajah, L., Schmidt, K. and Steinberg, D.K. (2019) "The importance of Antarctic krill in biogeochemical cycles". Nature communications, 10(1): 1–13. doi:10.1038/s41467-019-12668-7.
 Material was copied from this source, which is available under a Creative Commons Attribution 4.0 International License.
Material was copied from this source, which is available under a Creative Commons Attribution 4.0 International License. - Steinberg, Deborah; Sarah Goldthwait; Dennis Hansell (2002). "Zooplankton vertical migration and the active transport of dissolved organic and inorganic nitrogen in the Sargasso Sea". Deep-Sea Research Part I. 49 (8): 1445–1461. Bibcode:2002DSRI...49.1445S. CiteSeerX 10.1.1.391.7622. doi:10.1016/S0967-0637(02)00037-7. ISSN 0967-0637.
- Suttle, Curtis A. (October 2007). "Marine viruses — major players in the global ecosystem". Nature Reviews Microbiology. 5 (10): 801–812. doi:10.1038/nrmicro1750. ISSN 1740-1534. PMID 17853907. S2CID 4658457.
- Eppley, Richard W.; Peterson, Bruce J. (December 1979). "Particulate organic matter flux and planktonic new production in the deep ocean". Nature. 282 (5740): 677–680. Bibcode:1979Natur.282..677E. doi:10.1038/282677a0. ISSN 1476-4687. S2CID 42385900.
- Siegel, D. A.; Buesseler, K. O.; Doney, S. C.; Sailley, S. F.; Behrenfeld, M. J.; Boyd, P. W. (2014). "Global assessment of ocean carbon export by combining satellite observations and food-web models". Global Biogeochemical Cycles. 28 (3): 181–196. Bibcode:2014GBioC..28..181S. doi:10.1002/2013GB004743. hdl:1912/6668. ISSN 1944-9224. S2CID 43799803.
- Kobari, Toru; Steinberg, Deborah K.; Ueda, Ai; Tsuda, Atsushi; Silver, Mary W.; Kitamura, Minoru (2008-07-01). "Impacts of ontogenetically migrating copepods on downward carbon flux in the western subarctic Pacific Ocean". Deep Sea Research Part II: Topical Studies in Oceanography. Understanding the Ocean's Biological Pump:results from VERTIGO. 55 (14): 1648–1660. Bibcode:2008DSRII..55.1648K. doi:10.1016/j.dsr2.2008.04.016. ISSN 0967-0645.
- Bradford-Grieve, J. M.; Nodder, S. D.; Jillett, J. B.; Currie, K.; Lassey, K. R. (2001-09-01). "Potential contribution that the copepod Neocalanus tonsus makes to downward carbon flux in the Southern Ocean". Journal of Plankton Research. 23 (9): 963–975. doi:10.1093/plankt/23.9.963. ISSN 0142-7873.
- Wiebe, P.H; L.P. Madin; L.R. Haury; G.R. Harbison; L.M. Philbin (1979). "Diel Vertical Migration by Salpa aspera and its potential for large-scale particulate organic matter transport to the deep-sea". Marine Biology. 53 (3): 249–255. doi:10.1007/BF00952433. S2CID 85127670.
| Internal rhythms |  | |
|---|---|---|
| External cycles | ||
| Fields | ||
| See also |
| |
| About fish |  | ||||
|---|---|---|---|---|---|
| Anatomy and physiology | |||||
| Sensory systems | |||||
| Reproduction | |||||
| Locomotion | |||||
| Other behaviour | |||||
| By habitat | |||||
| Other types | |||||
| Commerce |
| ||||
| Major groups | |||||
| Lists |
| ||||
| |||||

| Biological swarming |  | |
|---|---|---|
| Animal migration | ||
| Swarm algorithms | ||
| Collective motion | ||
| Swarm robotics | ||
| Related topics | ||