Glucose-6-phosphate exchanger SLC37A4
Glucose-6-phosphate exchanger SLC37A4, also known as glucose-6-phosphate translocase, is an enzyme that in humans is encoded by the SLC37A4 gene.[5][6][7]
| SLC37A4 | |||||||||||||||||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Identifiers | |||||||||||||||||||||||||||||||||||||||||||||||||||
| Aliases | SLC37A4, G6PT1, G6PT2, G6PT3, GSD1b, GSD1c, GSD1d, TRG-19, TRG19, PRO0685, solute carrier family 37 member 4, CDG2W | ||||||||||||||||||||||||||||||||||||||||||||||||||
| External IDs | OMIM: 602671 MGI: 1316650 HomoloGene: 37482 GeneCards: SLC37A4 | ||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||
| Wikidata | |||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||




It consists of three subunits, each of which are vital components of the multi-enzyme Glucose-6-Phosphatase Complex (G6Pase). This important enzyme complex is located within the membrane of the endoplasmic reticulum, and catalyzes the terminal reactions in both glycogenolysis and gluconeogenesis.[8] The G6Pase complex is most abundant in liver tissue, but also present in kidney cells, small intestine, pancreatic islets and at a lower concentration in the gallbladder.[9][10] The G6Pase complex is highly involved in the regulation of homeostasis and blood glucose levels. Within this framework of glucose regulation, the translocase components are responsible for transporting the substrates and products across the endoplasmic reticulum membrane, resulting in the release of free glucose into the bloodstream.[8]
Structure
Glucose-6-phosphate translocase is a transmembrane protein providing a selective channel between the endoplasmic reticulum lumen and the cytosol. The enzyme is made up of three separate transporting subunits referred to as G6PT1 (subunit 1), G6PT2 (subunit 2) and G6PT3 (subunit 3). While the hydrolyzing component of the G6Pase complex is located on the side of the membrane on which it acts, namely facing the lumen, the translocases are all integral membrane proteins in order to perform their function as cross-membrane transporters. The translocases are spatially located on either side of the active site of the hydrolyzing component within the membrane, which allows the greatest speed and facility of the reaction.[11]
Mechanism
Each of the translocase subunits performs a specific function in the transport of substrates and products, and finally release of glucose (which will eventually reach the bloodstream), as a step in glycogenolysis or gluconeogenesis. G6PT1 transports Glucose-6-Phosphate from the cytosol into the lumen of the endoplasmic reticulum, where it is hydrolyzed by the catalytic subunit of G6Pase. After hydrolysis, glucose and inorganic phosphate are transported back into the cytosol by G6PT2 and G6PT3, respectively.[12] While the exact chemistry of the enzyme remains unknown, studies have shown that the mechanism of the enzyme complex is highly dependent upon the membrane structure. For instance, the Michaelis Constant of the enzyme for glucose-6-phosphate decreases significantly upon membrane disruption.[13] The originally proposed mechanism of the G6Pase system involved a relatively unspecific hydrolase, suggesting that G6PT1 alone provides the high specificity for the overall reaction by selective transport into the lumen, where hydrolysis occurs. Supporting evidence for this proposed reaction includes the marked decrease in substrate specificity of hydrolysis upon membrane degradation.[13]
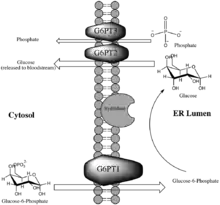
Figure 1 illustrates the role of G6P-Translocase within the G6Pase complex.
Inhibitors
Many inhibitors of glucose-6-phosphate translocase of novel, semi-synthetic or natural origin are known and of medical importance. Genetic algorithms for synthesizing novel inhibitors of G6PT1 have been developed and utilized in drug discovery.[14] Inhibitors of G6PT1 are the most studied as this subunit catalyzes the rate limiting step in glucose production through gluconeogenesis or glycogenolysis, and without its function these two processes could not occur. This inhibition holds great potential in drug development (discussed in "Medical and Disease Relevance"). Small-molecule inhibitors, such as mercaptopicolinic acid and diazobenzene sulfonate have some degree of inhibiting potential for G6PT1 but systematically lack specificity in inhibition, rendering them poor drug candidates.[15] Since the late 1990s, natural products have been increasingly studied as potent and specific inhibitors of G6PT1. Prominent examples of natural inhibitors include mumbaistatin and analogs, kodaistatin (harvested from extracts of Aspergillus terreus)[9] and chlorogenic acid.[16] Other natural product inhibitors of G6PT1 are found in the fungi Chaetomium carinthiacum, Bauhinia magalandra leaves, and streptomyces bacteria.[9][15]
Medical and disease relevance
1) Excessive activity of G6PT1 may contribute to the development of diabetes. Diabetes mellitus type 2 is a disease characterized by chronically elevated blood glucose levels, even when fasting.[17] The rapidly rising prevalence of type 2 diabetes, along with its strong correlation to heart disease and other health complications has rendered it an area of intense research with an urgent need for treatment options.[17] Studies monitoring blood glucose levels in rabbits revealed that the activity of G6Pase, and therefore G6PT1, is increased in specimens with diabetes. This strong correlation with diabetes type 2 makes the G6Pase complex, and G6PT1 in particular, an appealing drug target for control of blood glucose levels as its inhibition would directly prevent the release of free glucose into the bloodstream. It is possible that this mechanism of inhibition could be developed into a treatment for diabetes.[9]
2) The absence of a functional G6PT1 enzyme causes glycogen storage disease type Ib, commonly referred to as von Gierke disease, in humans. A common symptom of this disease is a build-up of glycogen in the liver and kidney causing enlargement of the organs.[16]
3) G6PT1 activity contributes to the survival of cells during hypoxia, which enables tumor cell growth and proliferation.[18]
See also
References
- ENSG00000281500 GRCh38: Ensembl release 89: ENSG00000137700, ENSG00000281500 - Ensembl, May 2017
- GRCm38: Ensembl release 89: ENSMUSG00000032114 - Ensembl, May 2017
- "Human PubMed Reference:". National Center for Biotechnology Information, U.S. National Library of Medicine.
- "Mouse PubMed Reference:". National Center for Biotechnology Information, U.S. National Library of Medicine.
- Gerin I, Veiga-da-Cunha M, Achouri Y, Collet JF, Van Schaftingen E (December 1997). "Sequence of a putative glucose 6-phosphate translocase, mutated in glycogen storage disease type Ib". FEBS Letters. 419 (2–3): 235–238. doi:10.1016/S0014-5793(97)01463-4. PMID 9428641. S2CID 31851796.
- Annabi B, Hiraiwa H, Mansfield BC, Lei KJ, Ubagai T, Polymeropoulos MH, et al. (February 1998). "The gene for glycogen-storage disease type 1b maps to chromosome 11q23". American Journal of Human Genetics. 62 (2): 400–405. doi:10.1086/301727. PMC 1376902. PMID 9463334.
- "Entrez Gene: SLC37A4 Solute carrier family 37 (glycerol-6-phosphate transporter), member 4".
- Parker JC, VanVolkenburg MA, Levy CB, Martin WH, Burk SH, Kwon Y, et al. (October 1998). "Plasma glucose levels are reduced in rats and mice treated with an inhibitor of glucose-6-phosphate translocase". Diabetes. 47 (10): 1630–1636. doi:10.2337/diabetes.47.10.1630. PMID 9753303.
- Parker, JC (2004). "Glucose-6-phosphatase inhibitors". Drugs of the Future. 29 (10): 1025–1033. doi:10.1358/dof.2004.029.10.863393.
- Hill A, Waddell ID, Hopwood D, Burchell A (May 1989). "The microsomal glucose-6-phosphatase enzyme of human gall-bladder". The Journal of Pathology. 158 (1): 53–56. doi:10.1002/path.1711580111. PMID 2547044. S2CID 2715249.
- van Schaftingen E, Gerin I (March 2002). "The glucose-6-phosphatase system". The Biochemical Journal. 362 (Pt 3): 513–532. doi:10.1042/0264-6021:3620513. PMC 1222414. PMID 11879177.
- Parker, JC (2001). "Glucose-6-phosphate translocase as a target for the design of antidiabetic agents". Drugs of the Future. 26 (7): 687–93. doi:10.1358/dof.2001.026.07.858712.
- Arion WJ, Wallin BK, Lange AJ, Ballas LM (February 1975). "On the involvement of a glucose 6-phosphate transport system in the function of microsomal glucose 6-phosphatase". Molecular and Cellular Biochemistry. 6 (2): 75–83. doi:10.1007/BF01732001. PMID 235736. S2CID 6980137.
- Bräuer S, Almstetter M, Antuch W, Behnke D, Taube R, Furer P, Hess S (Mar–Apr 2005). "Evolutionary chemistry approach toward finding novel inhibitors of the type 2 diabetes target glucose-6-phosphate translocase". Journal of Combinatorial Chemistry. 7 (2): 218–226. doi:10.1021/cc049867+. PMID 15762749.
- Lee TS, Das A, Khosla C (August 2007). "Structure-activity relationships of semisynthetic mumbaistatin analogs". Bioorganic & Medicinal Chemistry. 15 (15): 5207–5218. doi:10.1016/j.bmc.2007.05.019. PMID 17524653.
- Charkoudian, LK, et al. (April 2012). "Natural product inhibitors of glucose-6-phosphate translocase". Med. Chem. Commun. 3 (8): 926–31. doi:10.1039/C2MD20008B.
- American Diabetes Association (January 2012). "Standards of medical care in diabetes--2012". Diabetes Care. 35 (Suppl 1): S11–S63. doi:10.2337/dc12-s011. PMC 3632172. PMID 22187469.
- Tahanian E, Lord-Dufour S, Das A, Khosla C, Roy R, Annabi B (May 2010). "Inhibition of tubulogenesis and of carcinogen-mediated signaling in brain endothelial cells highlight the antiangiogenic properties of a mumbaistatin analog". Chemical Biology & Drug Design. 75 (5): 481–488. doi:10.1111/j.1747-0285.2010.00961.x. PMID 20486934. S2CID 205913311.
Further reading
- Chou JY, Matern D, Mansfield BC, Chen YT (March 2002). "Type I glycogen storage diseases: disorders of the glucose-6-phosphatase complex". Current Molecular Medicine. 2 (2): 121–143. doi:10.2174/1566524024605798. PMID 11949931.
- Narisawa K, Igarashi Y, Otomo H, Tada K (August 1978). "A new variant of glycogen storage disease type I probably due to a defect in the glucose-6-phosphate transport system". Biochemical and Biophysical Research Communications. 83 (4): 1360–1364. doi:10.1016/0006-291X(78)91371-2. PMID 212064.
- Fenske CD, Jeffery S, Weber JL, Houlston RS, Leonard JV, Lee PJ (April 1998). "Localisation of the gene for glycogen storage disease type 1c by homozygosity mapping to 11q". Journal of Medical Genetics. 35 (4): 269–272. doi:10.1136/jmg.35.4.269. PMC 1051271. PMID 9598717.
- Kure S, Suzuki Y, Matsubara Y, Sakamoto O, Shintaku H, Isshiki G, et al. (July 1998). "Molecular analysis of glycogen storage disease type Ib: identification of a prevalent mutation among Japanese patients and assignment of a putative glucose-6-phosphate translocase gene to chromosome 11". Biochemical and Biophysical Research Communications. 248 (2): 426–431. doi:10.1006/bbrc.1998.8985. PMID 9675154.
- Veiga-da-Cunha M, Gerin I, Chen YT, de Barsy T, de Lonlay P, Dionisi-Vici C, et al. (October 1998). "A gene on chromosome 11q23 coding for a putative glucose- 6-phosphate translocase is mutated in glycogen-storage disease types Ib and Ic". American Journal of Human Genetics. 63 (4): 976–983. doi:10.1086/302068. PMC 1377500. PMID 9758626.
- Marcolongo P, Barone V, Priori G, Pirola B, Giglio S, Biasucci G, et al. (October 1998). "Structure and mutation analysis of the glycogen storage disease type 1b gene". FEBS Letters. 436 (2): 247–250. doi:10.1016/S0014-5793(98)01129-6. PMID 9781688. S2CID 21464560.
- Lin B, Annabi B, Hiraiwa H, Pan CJ, Chou JY (November 1998). "Cloning and characterization of cDNAs encoding a candidate glycogen storage disease type 1b protein in rodents". The Journal of Biological Chemistry. 273 (48): 31656–31660. doi:10.1074/jbc.273.48.31656. PMID 9822626.
- Ihara K, Kuromaru R, Hara T (October 1998). "Genomic structure of the human glucose 6-phosphate translocase gene and novel mutations in the gene of a Japanese patient with glycogen storage disease type Ib". Human Genetics. 103 (4): 493–496. doi:10.1007/s004390050856. PMID 9856496. S2CID 26410800.
- Ihara K, Takabayashi A, Terasaki K, Hara T (1999). "Assignment1 of glucose 6-phosphate translocase (G6PT1) to human chromosome band 11q23.3 by in situ hybridization". Cytogenetics and Cell Genetics. 83 (1–2): 50–51. doi:10.1159/000015167. PMID 9925924. S2CID 35304470.
- Gerin I, Veiga-da-Cunha M, Noël G, Van Schaftingen E (February 1999). "Structure of the gene mutated in glycogen storage disease type Ib". Gene. 227 (2): 189–195. doi:10.1016/S0378-1119(98)00614-3. PMID 10023055.
- Hiraiwa H, Pan CJ, Lin B, Moses SW, Chou JY (February 1999). "Inactivation of the glucose 6-phosphate transporter causes glycogen storage disease type 1b". The Journal of Biological Chemistry. 274 (9): 5532–5536. doi:10.1074/jbc.274.9.5532. PMID 10026167.
- Pan CJ, Lin B, Chou JY (May 1999). "Transmembrane topology of human glucose 6-phosphate transporter". The Journal of Biological Chemistry. 274 (20): 13865–13869. doi:10.1074/jbc.274.20.13865. PMID 10318794.
- Janecke AR, Bosshard NU, Mayatepek E, Schulze A, Gitzelmann R, Burchell A, et al. (March 1999). "Molecular diagnosis of type 1c glycogen storage disease". Human Genetics. 104 (3): 275–277. doi:10.1007/s004390050948. PMID 10323254. S2CID 42696446.
- Hou DC, Kure S, Suzuki Y, Hasegawa Y, Hara Y, Inoue T, et al. (September 1999). "Glycogen storage disease type Ib: structural and mutational analysis of the microsomal glucose-6-phosphate transporter gene". American Journal of Medical Genetics. 86 (3): 253–257. doi:10.1002/(SICI)1096-8628(19990917)86:3<253::AID-AJMG11>3.0.CO;2-7. PMID 10482875.
- Veiga-da-Cunha M, Gerin I, Chen YT, Lee PJ, Leonard JV, Maire I, et al. (September 1999). "The putative glucose 6-phosphate translocase gene is mutated in essentially all cases of glycogen storage disease type I non-a". European Journal of Human Genetics. 7 (6): 717–723. doi:10.1038/sj.ejhg.5200366. PMID 10482962.
- Galli L, Orrico A, Marcolongo P, Fulceri R, Burchell A, Melis D, et al. (October 1999). "Mutations in the glucose-6-phosphate transporter (G6PT) gene in patients with glycogen storage diseases type 1b and 1c". FEBS Letters. 459 (2): 255–258. doi:10.1016/S0014-5793(99)01248-X. PMID 10518030. S2CID 41229244.
- Lam CW, Chan KY, Tong SF, Chan BY, Chan YT, Chan YW (July 2000). "A novel missense mutation (P191L) in the glucose-6-phosphate translocase gene identified in a Chinese family with glycogen storage disease 1b". Human Mutation. 16 (1): 94. doi:10.1002/1098-1004(200007)16:1<94::AID-HUMU26>3.0.CO;2-Q. PMID 10874322. S2CID 20061331.
- Santer R, Rischewski J, Block G, Kinner M, Wendel U, Schaub J, Schneppenheim R (August 2000). "Molecular analysis in glycogen storage disease 1 non-A: DHPLC detection of the highly prevalent exon 8 mutations of the G6PT1 gene in German patients". Human Mutation. 16 (2): 177. doi:10.1002/1098-1004(200008)16:2<177::AID-HUMU13>3.0.CO;2-8. PMID 10923042.