Glucosinolate
Glucosinolates are natural components of many pungent plants such as mustard, cabbage, and horseradish. The pungency of those plants is due to mustard oils produced from glucosinolates when the plant material is chewed, cut, or otherwise damaged. These natural chemicals most likely contribute to plant defence against pests and diseases, and impart a characteristic bitter flavor property to cruciferous vegetables.[1]
Occurrence
Glucosinolates occur as secondary metabolites of almost all plants of the order Brassicales. This includes the economically important family Brassicaceae as well as Capparaceae and Caricaceae. Outside of the Brassicales, the genera Drypetes[2] and Putranjiva in the family Putranjivaceae, are the only other known occurrence of glucosinolates. Glucosinolates occur in various edible plants such as cabbage (white cabbage, Chinese cabbage, broccoli), Brussels sprouts, watercress, horseradish, capers, and radishes where the breakdown products often contribute a significant part of the distinctive taste. The glucosinolates are also found in seeds of these plants.[1][3]
Chemistry
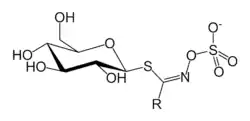
Glucosinolates constitute a natural class of organic compounds that contain sulfur and nitrogen and are derived from glucose and an amino acid. They are water-soluble anions and belong to the glucosides. Every glucosinolate contains a central carbon atom, which is bound to the sulfur atom of the thioglucose group, and via a nitrogen atom to a sulfate group (making a sulfated aldoxime). In addition, the central carbon is bound to a side group; different glucosinolates have different side groups, and it is variation in the side group that is responsible for the variation in the biological activities of these plant compounds. The essence of glucosinolate chemistry is their ability to convert into an isothiocyanate (a "mustard oil") upon hydrolysis of the thioglucoside bond by the enzyme myrosinase.[4]
The semisystematic naming of glucosinolates consists of the chemical name of the group "R" in the diagram followed by "glucosinolate", with or without a space. For example, allylglucosinolate and allyl glucosinolate refer to the same compound: both versions are found in the literature.[5] Isothiocyanates are conventionally written as two words.[4]
The following are some glucosinolates and their isothiocyanate products:[4]
- Allylglucosinolate (sinigrin) is the precursor of allyl isothiocyanate
- Benzylglucosinolate (glucotropaeolin) is the precursor of benzyl isothiocyanate
- Phenethylglucosinolate (gluconasturtiin) is the precursor of phenethyl isothiocyanate
- (R)-4-(methylsulfinyl)butylglucosinolate (glucoraphanin) is the precursor of (R)-4-(methylsulfinyl)butyl isothiocyanate (sulforaphane)
- (R)-2-hydroxybut-3-enylglucosinolate (progoitrin) is probably the precursor of (S)-2-hydroxybut-3-enyl isothiocyanate, which is expected to be unstable and immediately cyclize to form (S)-5-vinyloxazolidine-2-thione (goitrin)
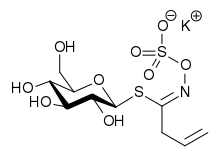
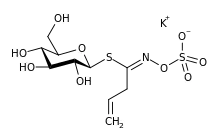
Sinigrin was first of the class to be isolated — in 1839 as its potassium salt.[6] Its chemical structure had been established by 1930, showing that it is a glucose derivative with β-D-glucopyranose configuration. It was unclear at that time whether the C=N bond was in the Z (or syn) form, with sulfur and oxygen substituents on the same side of the double bond, or the alternative E form in which they are on opposite sides. The matter was settled by X-ray crystallography in 1963.[7][8] It is now known that all natural glucosinolates are of Z form, although both forms can be made in the laboratory.[5] The "ate" ending in the naming of these compounds implies that they are anions at physiological pH and an early name for this allylglucosinolate was potassium myronate.[6] Care must be taken when discussing these compounds since some older publications do not make it clear whether they refer to the anion alone, its corresponding acid or the potassium salt.[5]
Biochemistry
Natural diversity from a few amino acids
About 132 different glucosinolates are known to occur naturally in plants. They are biosynthesized from amino acids: so-called aliphatic glucosinolates derived from mainly methionine, but also alanine, leucine, isoleucine, or valine. (Most glucosinolates are actually derived from chain-elongated homologues of these amino acids, e.g. glucoraphanin is derived from dihomomethionine, which is methionine chain-elongated twice.) Aromatic glucosinolates include indolic glucosinolates, such as glucobrassicin, derived from tryptophan and others from phenylalanine, its chain-elongated homologue homophenylalanine, and sinalbin derived from tyrosine.[4]
Biosynthetic pathway
Full details of the sequence of reactions that converts individual amino acids into the corresponding glucosinolate have been studied in the cress Arabidopsis thaliana.[9][5]
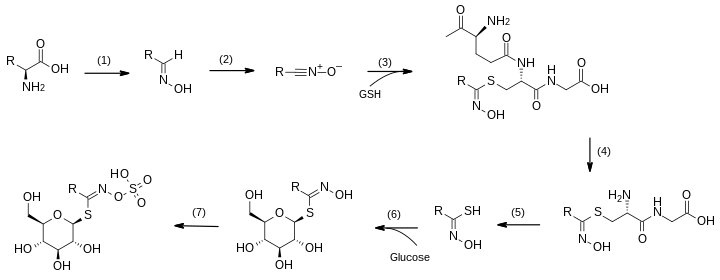
A sequence of seven enzyme-catalysed steps is used. The sulfur atom is incorporated from glutathione (GSH) and the sugar component is added to the resulting thiol derivative by a glycosyltransferase before the final sulfonation step.[10]
Enzymatic activation
The plants contain the enzyme myrosinase, which, in the presence of water, cleaves off the glucose group from a glucosinolate.[11] The remaining molecule then quickly converts to an isothiocyanate, a nitrile, or a thiocyanate; these are the active substances that serve as defense for the plant. Glucosinolates are also called mustard oil glycosides. The standard product of the reaction is the isothiocyanate (mustard oil); the other two products mainly occur in the presence of specialised plant proteins that alter the outcome of the reaction.[12]
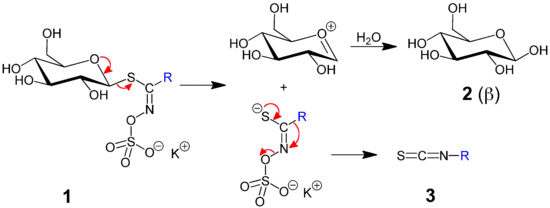
In the chemical reaction illustrated above, the red curved arrows in the left side of figure are simplified compared to reality, as the role of the enzyme myrosinase is not shown. However, the mechanism shown is fundamentally in accordance with the enzyme-catalyzed reaction.
In contrast, the reaction illustrated by red curved arrows at the right side of the figure, depicting the rearrangement of atoms resulting in the isothiocyanate, is expected to be non-enzymatic. This type of rearrangement can be named a Lossen rearrangement, or a Lossen-like rearrangement, since this name was first used for the analogous reaction leading to an organic isocyanate (R-N=C=O).
To prevent damage to the plant itself, the myrosinase and glucosinolates are stored in separate compartments of the cell or in different cells in the tissue, and come together only or mainly under conditions of physical injury (see Myrosinase).
Biological effects
Toxicity
The use of glucosinolate-containing crops as primary food source for animals can have negative effects if the concentration of glucosinolate is higher than what is acceptable for the animal in question, because some glucosinolates have been shown to have toxic effects (mainly as goitrogens and anti-thyroid agents) in livestock at high doses.[13] However, tolerance level to glucosinolates varies even within the same genus (e.g. Acomys cahirinus and Acomys russatus).[14]
Taste and eating behavior
The glucosinolate sinigrin, among others, was shown to be responsible for the bitterness of cooked cauliflower and Brussels sprouts.[1][15] Glucosinolates may alter animal eating behavior.[16]
Research
The isothiocyanates formed from glucosinolates are under laboratory research to assess the expression and activation of enzymes that metabolize xenobiotics, such as carcinogens.[17] Observational studies have been conducted to determine if consumption of cruciferous vegetables affects cancer risk in humans, but there is insufficient clinical evidence to indicate that consuming isothiocyanates in cruciferous vegetables is beneficial, according to a 2017 review.[17]
Insects
Glucosinolates and their products have a negative effect on many insects, resulting from a combination of deterrence and toxicity. In an attempt to apply this principle in an agronomic context, some glucosinolate-derived products can serve as antifeedants, i.e., natural pesticides.[18]
In contrast, the diamondback moth, a pest of cruciferous plants, may recognize the presence of glucosinolates, allowing it to identify the proper host plant.[19] Indeed, a characteristic, specialised insect fauna is found on glucosinolate-containing plants, including butterflies, such as large white, small white, and orange tip, but also certain aphids, moths, such as the southern armyworm, sawflies, and flea beetles. For instance, the large white butterfly deposits its eggs on these glucosinolate-containing plants, and the larvae survive even with high levels of glucosinolates and eat plant material containing glucosinolates.[20] The whites and orange tips all possess the so-called nitrile specifier protein, which diverts glucosinolate hydrolysis toward nitriles rather than reactive isothiocyanates.[21] In contrast, the diamondback moth possesses a completely different protein, glucosinolate sulfatase, which desulfates glucosinolates, thereby making them unfit for degradation to toxic products by myrosinase.[22]
Other kinds of insects (specialised sawflies and aphids) sequester glucosinolates.[23] In specialised aphids, but not in sawflies, a distinct animal myrosinase is found in muscle tissue, leading to degradation of sequestered glucosinolates upon aphid tissue destruction.[24] This diverse panel of biochemical solutions to the same plant chemical plays a key role in the evolution of plant-insect relationships.[25]
Induced production
Plants produce glucosinolates in response to the degree of herbivory being suffered. Their production in relation to atmospheric CO2 concentrations is complex: increased CO2 can give increased, decreased or unchanged production and there may be genetic variation within the Brassicales.[26][27]
See also
References
- Ishida M, Hara M, Fukino N, Kakizaki T, Morimitsu Y (May 2014). "Glucosinolate metabolism, functionality and breeding for the improvement of Brassicaceae vegetables". Breeding Science. 64 (1): 48–59. doi:10.1270/jsbbs.64.48. PMC 4031110. PMID 24987290.
- Rodman JE, Karol KG, Price RA, Sytsma KJ (1996). "Molecules, Morphology, and Dahlgren's Expanded Order Capparales". Systematic Botany. 21 (3): 289–307. doi:10.2307/2419660. JSTOR 2419660.
- Fahey, Jed W.; Zalcmann, Amy T.; Talalay, Paul (2001). "The chemical diversity and distribution of glucosinolates and isothiocyanates among plants". Phytochemistry. 56 (1): 5–51. doi:10.1016/S0031-9422(00)00316-2. PMID 11198818.
- Agerbirk N, Olsen CE (May 2012). "Glucosinolate structures in evolution". Phytochemistry. 77: 16–45. doi:10.1016/j.phytochem.2012.02.005. PMID 22405332.
- Blažević, Ivica; Montaut, Sabine; Burčul, Franko; Olsen, Carl Erik; Burow, Meike; Rollin, Patrick; Agerbirk, Niels (2020). "Glucosinolate structural diversity, identification, chemical synthesis and metabolism in plants". Phytochemistry. 169: 112100. doi:10.1016/j.phytochem.2019.112100. PMID 31771793. S2CID 208318505.
- Benn, M. H.; Ettlinger, M. G. (1965). "The synthesis of sinigrin". Chemical Communications (19): 445. doi:10.1039/C19650000445.
- Waser, Jürg; Watson, William H. (1963). "Crystal Structure of Sinigrin". Nature. 198 (4887): 1297–1298. doi:10.1038/1981297b0. S2CID 4187013.
- Marsh, R. E.; Waser, J. (1970). "Refinement of the crystal structure of sinigrin". Acta Crystallographica Section B Structural Crystallography and Crystal Chemistry. 26 (7): 1030–1037. doi:10.1107/S0567740870003539.
- Sønderby, Ida E.; Geu-Flores, Fernando; Halkier, Barbara A. (2010). "Biosynthesis of glucosinolates – gene discovery and beyond". Trends in Plant Science. 15 (5): 283–290. doi:10.1016/j.tplants.2010.02.005. PMID 20303821.
- Petersen, Annette; Crocoll, Christoph; Halkier, Barbara Ann (2019). "De novo production of benzyl glucosinolate in Escherichia coli". Metabolic Engineering. 54: 24–34. doi:10.1016/j.ymben.2019.02.004. PMID 30831267. S2CID 73475853.
- Bongoni R, Verkerk R, Steenbekkers B, Dekker M, Stieger M (September 2014). "Evaluation of different cooking conditions on broccoli (Brassica oleracea var. italica) to improve the nutritional value and consumer acceptance". Plant Foods for Human Nutrition. 69 (3): 228–234. doi:10.1007/s11130-014-0420-2. PMID 24853375. S2CID 35228794.
- Burow M, Bergner A, Gershenzon J, Wittstock U (January 2007). "Glucosinolate hydrolysis in Lepidium sativum--identification of the thiocyanate-forming protein". Plant Molecular Biology. 63 (1): 49–61. doi:10.1007/s11103-006-9071-5. PMID 17139450. S2CID 22955134.
- "Plants Poisonous to Livestock: Glucosinolates (Goitrogenic Glycosides)". Cornell University, Department of Animal Science. 10 September 2015. Retrieved 16 August 2018.
- Samuni-Blank M, Arad Z, Dearing MD, Gerchman Y, Karasov WH, Izhaki I (2013). "Friend or foe? Disparate plant–animal interactions of two congeneric rodents". Evolutionary Ecology. 27 (6): 1069–1080. doi:10.1007/s10682-013-9655-x. S2CID 280376.
- Van Doorn HE, Van der Kruk GC, van Holst GJ, Raaijmakers-Ruijs NC, Postma E, Groeneweg B, Jongen WH (1998). "The glucosinolates sinigrin and progoitrin are important determinants for taste preference and bitterness of Brussels sprouts". Journal of the Science of Food and Agriculture. 78: 30–38. doi:10.1002/(SICI)1097-0010(199809)78:1<30::AID-JSFA79>3.0.CO;2-N.
- Samuni-Blank M, Izhaki I, Dearing MD, Gerchman Y, Trabelcy B, Lotan A, Karasov WH, Arad Z (July 2012). "Intraspecific directed deterrence by the mustard oil bomb in a desert plant". Current Biology. 10 (22): 1218–1220. doi:10.1016/j.cub.2012.04.051. PMID 22704992.
- "Isothiocyanates". Micronutrient Information Center, Linus Pauling Institute, Oregon State University. 1 April 2017. Retrieved 26 June 2022.
- Furlan L, Bonetto C, Finotto A, Lazzeri L, Malaguti L, Patalano G, Parker W (2010). "The efficacy of biofumigant meals and plants to control wireworm populations". Industrial Crops and Products. 31 (2): 245–254. doi:10.1016/j.indcrop.2009.10.012.
- Badenes-Pérez FR, Reichelt M, Gershenzon J, Heckel DG (January 2011). "Phylloplane location of glucosinolates in Barbarea spp. (Brassicaceae) and misleading assessment of host suitability by a specialist herbivore". The New Phytologist. 189 (2): 549–556. doi:10.1111/j.1469-8137.2010.03486.x. PMID 21029103.
- David, W. A. L.; Gardiner, B. O. C. (1962). "Oviposition and the hatching of the eggs of Pieris brassicae (L.) in a laboratory culture". Bulletin of Entomological Research. 53: 91–109. doi:10.1017/S0007485300047982.
- Wittstock U, Agerbirk N, Stauber EJ, Olsen CE, Hippler M, Mitchell-Olds T, et al. (April 2004). "Successful herbivore attack due to metabolic diversion of a plant chemical defense". Proceedings of the National Academy of Sciences of the United States of America. 101 (14): 4859–4864. Bibcode:2004PNAS..101.4859W. doi:10.1073/pnas.0308007101. PMC 387339. PMID 15051878.
- Ratzka A, Vogel H, Kliebenstein DJ, Mitchell-Olds T, Kroymann J (August 2002). "Disarming the mustard oil bomb". Proceedings of the National Academy of Sciences of the United States of America. 99 (17): 11223–11228. Bibcode:2002PNAS...9911223R. doi:10.1073/pnas.172112899. PMC 123237. PMID 12161563.
- Müller C, Agerbirk N, Olsen CE, Boevé JL, Schaffner U, Brakefield PM (December 2001). "Sequestration of host plant glucosinolates in the defensive hemolymph of the sawfly Athalia rosae". Journal of Chemical Ecology. 27 (12): 2505–2516. doi:10.1023/A:1013631616141. PMID 11789955. S2CID 24529256.
- Bridges M, Jones AM, Bones AM, Hodgson C, Cole R, Bartlet E, et al. (January 2002). "Spatial organization of the glucosinolate-myrosinase system in brassica specialist aphids is similar to that of the host plant". Proceedings. Biological Sciences. 269 (1487): 187–191. doi:10.1098/rspb.2001.1861. PMC 1690872. PMID 11798435.
- Wheat CW, Vogel H, Wittstock U, Braby MF, Underwood D, Mitchell-Olds T (December 2007). "The genetic basis of a plant-insect coevolutionary key innovation". Proceedings of the National Academy of Sciences of the United States of America. 104 (51): 20427–20431. Bibcode:2007PNAS..10420427W. doi:10.1073/pnas.0706229104. PMC 2154447. PMID 18077380.
- Bidart-Bouzat, M. Gabriela; Imeh-Nathaniel, Adebobola (2008). "Global Change Effects on Plant Chemical Defenses against Insect Herbivores". Journal of Integrative Plant Biology. 50 (11): 1339–1354. doi:10.1111/j.1744-7909.2008.00751.x. PMID 19017122.
- Zavala JA, Nabity PD, DeLucia EH (7 January 2013). "An emerging understanding of mechanisms governing insect herbivory under elevated CO2". Annual Review of Entomology. Annual Reviews. 58 (1): 79–97. doi:10.1146/annurev-ento-120811-153544. PMID 22974069.
External links
- Glucosinolate metabolism pathways from MetaCyc