Haplogroup N (mtDNA)
Haplogroup N is a human mitochondrial DNA (mtDNA) clade. A macrohaplogroup, its descendant lineages are distributed across many continents. Like its sibling macrohaplogroup M, macrohaplogroup N is a descendant of the haplogroup L3.
| Haplogroup N | |
|---|---|
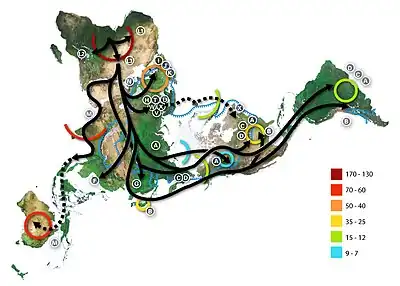 Ancient dispersal of haplogroup L3, its descendant M and N lineages, and other mtDNA clades. Numbers represent thousand years before present. | |
| Possible time of origin | ~55-70,000 YBP[1][2] or 50-65,000 YBP[3] |
| Possible place of origin | Asia[4][5][6][7][8] or East Africa[9][10][11] |
| Ancestor | L3 |
| Descendants | N1'5, N2, N8, N9, N10, N11, N13, N14, N21, N22, A, I, O, R, S, X, Y, W |
| Defining mutations | 8701, 9540, 10398, 10873, 15301[12] |
All mtDNA haplogroups found outside of Africa are descendants of either haplogroup N or its sibling haplogroup M. M and N are the signature maternal haplogroups that define the theory of the recent African origin of modern humans and subsequent early human migrations around the world. The global distribution of haplogroups N and M indicates that there was likely at least one major prehistoric migration of humans out of Africa, with both N and M later evolving outside the continent.[7]
Origins

There is widespread agreement in the scientific community concerning the African ancestry of haplogroup L3 (haplogroup N's parent clade).[13] However, whether or not the mutations which define haplogroup N itself first occurred within Asia or Africa has been a subject for ongoing discussion and study.[13]
The out of Africa hypothesis has gained generalized consensus. However, many specific questions remain unsettled. To know whether the two M and N macrohaplogroups that colonized Eurasia were already present in Africa before the exit is puzzling.
Torroni et al. 2006 state that Haplogroups M, N and R occurred somewhere between East Africa and the Persian Gulf.[14]
Also related to the origins of haplogroup N is whether ancestral haplogroups M, N and R were part of the same migration out of Africa, or whether Haplogroup N left Africa via the Northern route through the Levant, and M left Africa via Horn of Africa. This theory was suggested because haplogroup N is by far the predominant haplogroup in Western Eurasia, and haplogroup M is absent in Western Eurasia, but is predominant in India and is common in regions East of India. However, the mitochondrial DNA variation in isolated "relict" populations in southeast Asia and among Indigenous Australians supports the view that there was only a single dispersal from Africa. Southeast Asian populations and Indigenous Australians all possess deep rooted clades of both haplogroups M and N.[15] The distribution of the earliest branches within haplogroups M, N, and R across Eurasia and Oceania therefore supports a three-founder-mtDNA scenario and a single migration route out of Africa.[16] These findings also highlight the importance of Indian subcontinent in the early genetic history of human settlement and expansion.[17]
Asian origin hypothesis
The hypothesis of Asia as the place of origin of haplogroup N is supported by the following:
- Haplogroup N is found in all parts of the world but has low frequencies in Sub-Saharan Africa. According to a number of studies, the presence of Haplogroup N in Africa is most likely the result of back migration from Eurasia.[6]
- The oldest clades of macrohaplogroup N are found in Asia and Australia.
- It would be paradoxical that haplogroup N had traveled all the distance to Australia or New World yet failed to affect other populations within Africa besides North Africans and Horn Africans.
- The mitochondrial DNA variation in isolated "relict" populations in southeast Asia supports the view that there was only a single dispersal from Africa.[15] The distribution of the earliest branches within haplogroups M, N, and R across Eurasia and Oceania provides additional evidence for a three-founder-mtDNA scenario and a single migration route out of Africa.[16] These findings also highlight the importance of Indian subcontinent in the early genetic history of human settlement and expansion.[17] Therefore, N's history is similar to M and R which have their most probable origin in South Asia.
A study (Vai et al. 2019), finds a basal branch of maternal haplogroup N in early Neolithic North African remains from the Libyan site of Takarkori. The authors propose that N most likely split from L3 in the Arabian peninsula and later migrated back to North Africa, with its sister haplogroup M also likely splitting from L3 in the Middle East, but also suggest that N may have possibly diverged in North Africa, and state that more information is necessary to be certain.[3]
African origin hypothesis
According to Toomas Kivisild "the lack of L3 lineages other than M and N in India and among non-African mitochondria in general suggests that the earliest migration(s) of modern humans already carried these two mtDNA ancestors, via a departure route over the Horn of Africa.[9]
In 2019, a study by Vai et al. presented evidence of a basal branch of haplogroup N from the Neolithic Sahara. They suggest that N either diverged from haplogroup L3 in the Near East (possibly in the Arabian peninsula, following the exit of L3 from Africa), then back-migrated to North Africa, or that it instead may have originated in North Africa (having diverged from L3 there).[3]
Distribution
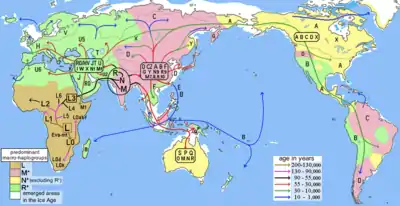
Haplogroup N is derived from the ancestral L3 macrohaplogroup, which represents the migration discussed in the theory of the recent African origin of modern humans. Haplogroup N is the ancestral haplogroup to almost all clades today distributed in Europe and Oceania, as well as many found in Asia and the Americas. It is believed to have arisen at a similar time to haplogroup M. Haplogroup N subclades like haplogroup U6 are also found at high to low frequencies in northwest and northeast Africa due to a back migration from Europe or Asia during the Paleolithic c. 46,000 ybp, the estimated age of the basal U6* clade.[18] Other haplogroups common in Western Eurasia, such as R0, J, and T, are also common in North Africa and parts of East Africa.
The haplogroup N descendant lineage U6 has been found among Iberomaurusian specimens at the Taforalt site, which date from the Epipaleolithic.[19] In Sub-Saharan Africa, several ancient samples of N have been found, the oldest so far being K1a dating back to about 2000 BC in Kenya.[20] Additionally, haplogroup N predominated among ancient Egyptian mummies excavated at the Abusir el-Meleq archaeological site in Middle Egypt, which date from the Pre-Ptolemaic/late New Kingdom, Ptolemaic, and Roman periods.[21]
Subgroups distribution
Haplogroup N's derived clades include the macro-haplogroup R and its descendants, and haplogroups A, I, S, W, X, and Y.
Rare unclassified haplogroup N* has been found among fossils belonging to the Cardial and Epicardial culture (Cardium pottery) and the Pre-Pottery Neolithic B.[22] A rare unclassified form of N has been also been reported in modern Algeria.[23]
- Haplogroup N1'5
- Haplogroup N1 – found in Africa .[24]
- Haplogroup N1b – found in Middle East, Egypt (Gurna),[25] Caucasus and Europe.
- N1a'c'd'e'I
- Haplogroup N1c – Northern Saudi Arabia, Turkey [8]
- N1a'd'e'I
- Haplogroup N1d – India
- N1a'e'I
- Haplogroup N1a – Arabian Peninsula and Northeast Africa.[8] Found also in Central Asia and Southern Siberia.[26] This branch is well attested in ancient people from various cultures of Neolithic Europe, from Hungary to Spain,[27] and among the earliest farmers of Anatolia.[28]
- N1e'I
- Haplogroup N5 – found in India.[16]
- Haplogroup N1 – found in Africa .[24]
- Haplogroup N2
- Haplogroup N2a – small clade found in West Europe.[31]
- Haplogroup W[32] – found in Western Eurasia and South Asia[33]
- Haplogroup N3 – all subgroups have so far only been found in Belarus
- Haplogroup N3a
- Haplogroup N3a1
- Haplogroup N3b
- Haplogroup N7 – all subgroups have so far only been found in Cambodia
- Haplogroup N7a
- Haplogroup N7a1
- Haplogroup N7a2
- Haplogroup N7b
- Haplogroup N7a
- Haplogroup N8 – found in China.[34]
- Haplogroup N9 – found in Far East.[24] [TMRCA 45,709.7 ± 7,931.5 ybp; CI=95%[35]]
- Haplogroup N9a [TMRCA 17,520.4 ± 4,389.8 ybp; CI=95%[35]]
- Haplogroup N9a12 – Khon Mueang (Pai District)[36][37]
- Haplogroup N9a-C16261T
- Haplogroup N9a-C16261T* – Vietnam (Kinh)[38][37]
- Haplogroup N9a-A4129G-A4913G-T12354C-A12612G-C12636T-T16311C!!! – Tashkurgan (Kyrgyz)[39][37]
- Haplogroup N9a1'3 [TMRCA 15,007.4 ± 6,060.1 ybp; CI=95%[35]]
- Haplogroup N9a1 – Chinese (Hakka in Taiwan, etc.), She, Tu, Uyghur, Tuvan, Mongolia, Khamnigan,[40] Korea,[41][40] Japan [TMRCA 9,200 (95% CI 7,100 <-> 11,600) ybp[37]]
- Haplogroup N9a1a – Chinese (Sichuan, Zhanjiang, etc.) [TMRCA 7,300 (95% CI 3,800 <-> 12,800) ybp[37]]
- Haplogroup N9a1b – Kyrgyz (Tashkurgan)[39][37]
- Haplogroup N9a1c – Vietnam (Tay people),[42] Thailand (Khon Mueang from Chiang Mai Province,[36] Lao Isan from Loei Province[36])
- Haplogroup N9a3 – China [TMRCA 11,500 (95% CI 7,500 <-> 16,800) ybp[37]]
- Haplogroup N9a3a – Japan, Korean (Seoul), Taiwan (incl. Paiwan), Thailand (Mon from Lopburi Province and Kanchanaburi Province[36]), China, Uyghur, Kyrgyz (Tashkurgan), Kazakhstan, Buryat, Russia (Belgorod, Chechen Republic, etc.), Ukraine, Moldova, Belarus, Lithuania, Poland, Czech (West Bohemia), Hungary, Austria, Germany [TMRCA 8,280.9 ± 5,124.4 ybp; CI=95%[35]]
- Haplogroup N9a1 – Chinese (Hakka in Taiwan, etc.), She, Tu, Uyghur, Tuvan, Mongolia, Khamnigan,[40] Korea,[41][40] Japan [TMRCA 9,200 (95% CI 7,100 <-> 11,600) ybp[37]]
- Haplogroup N9a2'4'5'11 [TMRCA 15,305.4 ± 4,022.6 ybp; CI=95%[35]]
- Haplogroup N9a2 – Japan, Korea,[41] China (Barghut in Hulunbuir, Uyghur, etc.) [TMRCA 10,700 (95% CI 8,200 <-> 13,800) ybp[37]]
- Haplogroup N9a2a – Japan, Korea, Uyghur [TMRCA 8,100 (95% CI 6,500 <-> 10,000) ybp[37]]
- Haplogroup N9a2a1 – Japan [TMRCA 4,200 (95% CI 1,850 <-> 8,400) ybp[37]]
- Haplogroup N9a2a2 – Japan, Korea, Volga-Ural region (Tatar)[40] [TMRCA 5,700 (95% CI 3,500 <-> 8,900) ybp[37]]
- Haplogroup N9a2a3 – Japan, Hulun-Buir region (Barghut)[40] [TMRCA 4,700 (95% CI 2,400 <-> 8,400) ybp[37]]
- Haplogroup N9a2a4 – Japan [TMRCA 2,800 (95% CI 600 <-> 7,900) ybp[37]]
- Haplogroup N9a2b – China
- Haplogroup N9a2c [TMRCA 7,200 (95% CI 3,600 <-> 12,700) ybp[37]]
- Haplogroup N9a2d – Japan, Korea[37] [TMRCA 5,200 (95% CI 1,800 <-> 12,000) ybp[37]]
- Haplogroup N9a2e – China
- Haplogroup N9a2a – Japan, Korea, Uyghur [TMRCA 8,100 (95% CI 6,500 <-> 10,000) ybp[37]]
- Haplogroup N9a4 – Malaysia [TMRCA 7,900 (95% CI 3,900 <-> 14,300) ybp[37]]
- Haplogroup N9a5 [TMRCA 8,700 (95% CI 4,700 <-> 15,000) ybp[37]]
- Haplogroup N9a5* – Korea
- Haplogroup N9a5a – Japan
- Haplogroup N9a5b – Japan [TMRCA 5,300 (95% CI 1,150 <-> 15,300) ybp[37]]
- Haplogroup N9a11 – Taiwan (Hakka, Minnan), Laos (Lao from Luang Prabang[36])
- Haplogroup N9a2 – Japan, Korea,[41] China (Barghut in Hulunbuir, Uyghur, etc.) [TMRCA 10,700 (95% CI 8,200 <-> 13,800) ybp[37]]
- Haplogroup N9a6 – Thailand (Phuan from Lopburi Province,[36] Khon Mueang from Lamphun Province,[36] Phutai from Sakon Nakhon Province,[36] Lawa from Mae Hong Son Province,[36] Soa from Sakon Nakhon Province[36]), Vietnam, Sumatra [TMRCA 11,972.5 ± 5,491.7 ybp; CI=95%[35]]
- Haplogroup N9a6a – Cambodia (Khmer), Malaysia (Bidayuh, Jehai, Temuan, Kensiu), Sumatra, Sundanese
- Haplogroup N9a6b – Malaysia (Seletar)
- Haplogroup N9a7 – Japan
- Haplogroup N9a8 – Japan, China, Buryat
- Haplogroup N9a9 – Chelkans (Biyka, Turochak), Tubalar (North-East Altai), Kyrgyz (Kyrgyzstan), China, Ukraine (Vinnytsia Oblast), Romania (10th century AD Dobruja)
- Haplogroup N9a10 – Thailand (Khon Mueang from Mae Hong Son Province, Chiang Mai Province, Lamphun Province, and Lampang Province,[36] Shan from Mae Hong Son Province,[36] Lao Isan from Loei Province,[36] Black Tai from Kanchanaburi Province,[36] Phuan from Sukhothai Province and Phichit Province,[36] Mon from Kanchanaburi Province[36]), Laos (Lao from Luang Prabang,[36] Hmong[38]), Vietnam (Tay Nung), China (incl. Han in Chongqing)
- Haplogroup N9a10a – China, Taiwan (Ami)
- Haplogroup N9a10a1 – Chinese (Suzhou)
- Haplogroup N9a10a2 – Philippines (Ivatan), Taiwan (Ami)
- Haplogroup N9a10a2a – Taiwan (Atayal, Tsou)
- Haplogroup N9a10b – China
- Haplogroup N9a10a – China, Taiwan (Ami)
- Haplogroup N9b – Japan, Udegey, Nanai, Korea[41] [TMRCA 14,885.6 ± 4,092.5 ybp; CI=95%[35]]
- Haplogroup Y[43] – found especially among Nivkhs, Ulchs, Nanais, Negidals, Ainus, and the population of Nias Island, with a moderate frequency among other Tungusic peoples, Koreans, Mongols, Koryaks, Itelmens, Chinese, Japanese, Tajiks, Island Southeast Asians (including Taiwanese aborigines), and some Turkic peoples[26] [TMRCA 24,576.4 ± 7,083.2 ybp; CI=95%[35]]
- Haplogroup Y1 – Korea, Taiwan (Minnan), Thailand (Iu Mien from Phayao Province[44][37]), Poland, Slovakia, Czech Republic [TMRCA 14,689.5 ± 5,264.3 ybp; CI=95%[35]]
- Haplogroup Y1a – Nivkh, Ulchi, Hezhen, Udegey, Even, Zabaikal Buryat, Mongolian, Daur, Korea, Han, Tibet, Ukraine[45] [TMRCA 7,467.5 ± 5,526.7 ybp; CI=95%[35]]
- Haplogroup Y1b – Volga Tatar [TMRCA 9,222.8 ± 4,967.0 ybp; CI=95%[35]]
- Haplogroup Y1c - Korea (especially Jeju Island), Khamnigan, Uyghur, Canada
- Haplogroup Y2 – Chinese, Japanese, Korean, Khamnigan, South Africa (Cape Coloured) [TMRCA 7,279.3 ± 2,894.5 ybp; CI=95%[35]]
- Haplogroup Y1 – Korea, Taiwan (Minnan), Thailand (Iu Mien from Phayao Province[44][37]), Poland, Slovakia, Czech Republic [TMRCA 14,689.5 ± 5,264.3 ybp; CI=95%[35]]
- Haplogroup N9a [TMRCA 17,520.4 ± 4,389.8 ybp; CI=95%[35]]
- Haplogroup N10 – found in China (Han from Shanghai, Jiangsu, Fujian, Guangdong, and Yunnan, Hani and Yi from Yunnan, She from Guizhou, Uzbek from Xinjiang) and Southeast Asia (Thailand, Indonesia, Vietnam, Malaysia).[34]
- Haplogroup N11 – Mainland China & Philippines: Han Chinese (Yunnan, Sichuan, and Hubei), Tibetan (Xizang), Dongxiang (Gansu), Oroqen (Inner Mongolia) and Mamanwa (Philippines).[34][47]
- Haplogroup O or N12- found among Indigenous Australians and the Floresians of Indonesia.
- Haplogroup N13 – Aboriginal Australians[48]
- Haplogroup N14 – Aboriginal Australians[49]
- Haplogroup N21 – Temuan, Semelai, Thailand, Khmer, ethnic Malays from Malaysia and Indonesia.[50]
- Haplogroup N22 – Southeast Asia, Bangladesh, India, Japan[51]
- Haplogroup A[52] – found in Central and East Asia, as well as among Native Americans.
- Haplogroup S[53] – extended among Aboriginal Australians.
- Haplogroup X[54] – found most often in Western Eurasia, but also present in the Americas.[24]
- Haplogroup X1 – found primarily in North Africa as well as in some populations of the Levant, notably among the Druze
- Haplogroup X2 – found in Western Eurasia, Siberia and among Native Americans
- Haplogroup R[55] – a very extended and diversified macro-haplogroup.
Subclades
Tree
This phylogenetic tree of haplogroup N subclades is based on the paper by Mannis van Oven and Manfred Kayser Updated comprehensive phylogenetic tree of global human mitochondrial DNA variation[12] and subsequent published research.
See also
|
Phylogenetic tree of human mitochondrial DNA (mtDNA) haplogroups | |||||||||||||||||||||||||||||||||||||||
| Mitochondrial Eve (L) | |||||||||||||||||||||||||||||||||||||||
| L0 | L1–6 | ||||||||||||||||||||||||||||||||||||||
| L1 | L2 | L3 | L4 | L5 | L6 | ||||||||||||||||||||||||||||||||||
| M | N | ||||||||||||||||||||||||||||||||||||||
| CZ | D | E | G | Q | O | A | S | R | I | W | X | Y | |||||||||||||||||||||||||||
| C | Z | B | F | R0 | pre-JT | P | U | ||||||||||||||||||||||||||||||||
| HV | JT | K | |||||||||||||||||||||||||||||||||||||
| H | V | J | T | ||||||||||||||||||||||||||||||||||||
References
- Soares, Pedro; Ermini, Luca; Thomson, Noel; Mormina, Maru; Rito, Teresa; Röhl, Arne; Salas, Antonio; Oppenheimer, Stephen; MacAulay, Vincent; Richards, Martin B. (2009). "Correcting for Purifying Selection: An Improved Human Mitochondrial Molecular Clock". The American Journal of Human Genetics. 84 (6): 740–59. doi:10.1016/j.ajhg.2009.05.001. PMC 2694979. PMID 19500773.
- Soares, P; Alshamali, F; Pereira, J. B; Fernandes, V; Silva, N. M; Afonso, C; Costa, M. D; Musilova, E; MacAulay, V; Richards, M. B; Cerny, V; Pereira, L (2011). "The Expansion of mtDNA Haplogroup L3 within and out of Africa". Molecular Biology and Evolution. 29 (3): 915–927. doi:10.1093/molbev/msr245. PMID 22096215.
- Vai S, Sarno S, Lari M, Luiselli D, Manzi G, Gallinaro M, Mataich S, Hübner A, Modi A, Pilli E, Tafuri MA, Caramelli D, di Lernia S (March 2019). "Ancestral mitochondrial N lineage from the Neolithic 'green' Sahara". Sci Rep. 9 (1): 3530. Bibcode:2019NatSR...9.3530V. doi:10.1038/s41598-019-39802-1. PMC 6401177. PMID 30837540.
- MacAulay, V.; Hill, C; Achilli, A; Rengo, C; Clarke, D; Meehan, W; Blackburn, J; Semino, O; et al. (2005). "Single, Rapid Coastal Settlement of Asia Revealed by Analysis of Complete Mitochondrial Genomes" (PDF). Science. 308 (5724): 1034–36. Bibcode:2005Sci...308.1034M. doi:10.1126/science.1109792. PMID 15890885. S2CID 31243109.
Haplogroup L3 (the African clade that gave rise to the two basal non-African clades, haplogroups M and N) is 84,000 years old, and haplogroups M and N themselves are almost identical in age at 63,000 years old, with haplogroup R diverging rapidly within haplogroup N 60,000 years ago.
- Richards, Martin; Bandelt, Hans-Jürgen; Kivisild, Toomas; Oppenheimer, Stephen (2006). "A Model for the Dispersal of Modern Humans out of Africa". In Bandelt, Hans-Jürgen; Macaulay, Vincent; Richards, Martin (eds.). Human Mitochondrial DNA and the Evolution of Homo sapiens. Nucleic Acids and Molecular Biology. Vol. 18. pp. 225–65. doi:10.1007/3-540-31789-9_10. ISBN 978-3-540-31788-3.
subclades. L3b d, L3e and L3f, for instance, are clearly of African origin, whereas haplogroup N is of apparently Eurasian origin
- Gonder, M. K.; Mortensen, H. M.; Reed, F. A.; De Sousa, A.; Tishkoff, S. A. (2006). "Whole-mtDNA Genome Sequence Analysis of Ancient African Lineages". Molecular Biology and Evolution. 24 (3): 757–68. doi:10.1093/molbev/msl209. PMID 17194802.
- Olivieri, A.; Achilli, A.; Pala, M.; Battaglia, V.; Fornarino, S.; Al-Zahery, N.; Scozzari, R.; Cruciani, F.; Behar, D. M.; Dugoujon, J.-M.; Coudray, C.; Santachiara-Benerecetti, A. S.; Semino, O.; Bandelt, H.-J.; Torroni, A. (2006). "The mtDNA Legacy of the Levantine Early Upper Palaeolithic in Africa". Science. 314 (5806): 1767–70. Bibcode:2006Sci...314.1767O. doi:10.1126/science.1135566. PMID 17170302. S2CID 3810151.
The scenario of a back-migration into Africa is supported by another feature of the mtDNA phylogeny. Haplogroup M's Eurasian sister clade, haplogroup N, which has a very similar age to M and no indication of an African origin
- Abu-Amero, Khaled K; Larruga, José M; Cabrera, Vicente M; González, Ana M (2008). "Mitochondrial DNA structure in the Arabian Peninsula". BMC Evolutionary Biology. 8: 45. doi:10.1186/1471-2148-8-45. PMC 2268671. PMID 18269758.
- Kivisild T, Rootsi S, Metspalu M, et al. (2003). "The genetic heritage of the earliest settlers persists both in Indian tribal and caste populations". American Journal of Human Genetics. 72 (2): 313–32. doi:10.1086/346068. PMC 379225. PMID 12536373."Also, the lack of L3 lineages other than M and N in India and among non-African mitochondria in general suggests that the earliest migration(s) of modern humans already carried these two mtDNA ancestors, via a departure route over the horn of Africa."
- Kivisild; et al. (2007). "Genetic Evidence of Modern Human Dispersals in South Asia". The Evolution and History of Human Populations in South Asia. ISBN 9781402055621.
- Osman, Maha M.; et al. "Mitochondrial HVRI and whole mitogenome sequence variations portray similar scenarios on the genetic structure and ancestry of northeast Africans" (PDF). Institute of Endemic Diseases. Meta Gene.
- van Oven M, Kayser M (2009). "Updated comprehensive phylogenetic tree of global human mitochondrial DNA variation". Human Mutation. 30 (2): E386–94. doi:10.1002/humu.20921. PMID 18853457. S2CID 27566749.
- González, Ana M; Larruga, José M; Abu-Amero, Khaled K; Shi, Yufei; Pestano, José; Cabrera, Vicente M (2007). "Mitochondrial lineage M1 traces an early human backflow to Africa". BMC Genomics. 8: 223. doi:10.1186/1471-2164-8-223. PMC 1945034. PMID 17620140.
- Torroni, A; Achilli, A; MacAulay, V; Richards, M; Bandelt, H (2006). "Harvesting the fruit of the human mtDNA tree". Trends in Genetics. 22 (6): 339–45. doi:10.1016/j.tig.2006.04.001. PMID 16678300.
- MacAulay, V.; Hill, C; Achilli, A; Rengo, C; Clarke, D; Meehan, W; Blackburn, J; Semino, O; et al. (2005). "Single, Rapid Coastal Settlement of Asia Revealed by Analysis of Complete Mitochondrial Genomes" (PDF). Science. 308 (5724): 1034–36. Bibcode:2005Sci...308.1034M. doi:10.1126/science.1109792. PMID 15890885. S2CID 31243109.
- Palanichamy MG, Sun C, Agrawal S, et al. (2004). "Phylogeny of mitochondrial DNA macrohaplogroup N in India, based on complete sequencing: implications for the peopling of South Asia". American Journal of Human Genetics. 75 (6): 966–78. doi:10.1086/425871. PMC 1182158. PMID 15467980.
- Maji, Suvendu; Krithika, S.; Vasulu, T. S. (2008). "Distribution of Mitochondrial DNA Macrohaplogroup N in India with Special Reference to Haplogroup R and its Sub-Haplogroup U" (PDF). International Journal of Human Genetics. 8 (1–2): 85–96. doi:10.1080/09723757.2008.11886022. S2CID 14231815.
- Hervella, M.; et al. (2016). "The mitogenome of a 35,000-year-old Homo sapiens from Europe supports a Paleolithic back-migration to Africa". Scientific Reports. 6: 25501. Bibcode:2016NatSR...625501H. doi:10.1038/srep25501. PMC 4872530. PMID 27195518.
- Bernard Secher; Rosa Fregel; José M Larruga; Vicente M Cabrera; Phillip Endicott; José J Pestano; Ana M González (2014). "The history of the North African mitochondrial DNA haplogroup U6 gene flow into the African, Eurasian and American continents". Scientific Reports. 14: 109. doi:10.1186/1471-2148-14-109. PMC 4062890. PMID 24885141.
- Prendergast, Mary E.; Lipson, Mark; Sawchuk, Elizabeth A.; Olalde, Iñigo; Ogola, Christine A.; Rohland, Nadin; Sirak, Kendra A.; Adamski, Nicole; Bernardos, Rebecca; Broomandkhoshbacht, Nasreen; Callan, Kimberly; Culleton, Brendan J.; Eccles, Laurie; Harper, Thomas K.; Lawson, Ann Marie; Mah, Matthew; Oppenheimer, Jonas; Stewardson, Kristin; Zalzala, Fatma; Ambrose, Stanley H.; Ayodo, George; Gates, Henry Louis; Gidna, Agness O.; Katongo, Maggie; Kwekason, Amandus; Mabulla, Audax Z. P.; Mudenda, George S.; Ndiema, Emmanuel K.; Nelson, Charles; Robertshaw, Peter; Kennett, Douglas J.; Manthi, Fredrick K.; Reich, David (5 July 2019). "Ancient DNA reveals a multistep spread of the first herders into sub-Saharan Africa". Science. 365 (6448): eaaw6275. Bibcode:2019Sci...365.6275P. doi:10.1126/science.aaw6275. PMC 6827346. PMID 31147405.
- Schuenemann, Verena J.; et al. (2017). "Ancient Egyptian mummy genomes suggest an increase of Sub-Saharan African ancestry in post-Roman periods". Nature Communications. 8: 15694. Bibcode:2017NatCo...815694S. doi:10.1038/ncomms15694. PMC 5459999. PMID 28556824.
- Fernández, Eva; et al. (2014). "Ancient DNA analysis of 8000 BC near eastern farmers supports an early neolithic pioneer maritime colonization of Mainland Europe through Cyprus and the Aegean Islands". PLOS Genetics. 10 (6): e1004401. doi:10.1371/journal.pgen.1004401. PMC 4046922. PMID 24901650.
- Bekada, Asmahan; Arauna, Lara R.; Deba, Tahria; Calafell, Francesc; Benhamamouch, Soraya; Comas, David; Kayser, Manfred (24 September 2015). "Genetic Heterogeneity in Algerian Human Populations". PLOS ONE. 10 (9): e0138453. Bibcode:2015PLoSO..1038453B. doi:10.1371/journal.pone.0138453. PMC 4581715. PMID 26402429.
- "Ian Logan's mtDNA site". Archived from the original on 2011-12-09. Retrieved 2009-07-17.
- A. Stevanovitch; A. Gilles; E. Bouzaid; R. Kefi; F. Paris; R. P. Gayraud; J. L. Spadoni; F. El-Chenawi; E. Béraud-Colomb (January 2004). "Mitochondrial DNA Sequence Diversity in a Sedentary Population from Egypt". Annals of Human Genetics. 68 (1): 23–39. doi:10.1046/j.1529-8817.2003.00057.x. PMID 14748828. S2CID 44901197.
- Derenko, M; Malyarchuk, B; Grzybowski, T; Denisova, G; Dambueva, I; Perkova, M; Dorzhu, C; Luzina, F; Lee, H; Vanecek, Tomas; Villems, Richard; Zakharov, Ilia (2007). "Phylogeographic Analysis of Mitochondrial DNA in Northern Asian Populations". The American Journal of Human Genetics. 81 (5): 1025–41. doi:10.1086/522933. PMC 2265662. PMID 17924343.
- Lipson, Mark; Szécsényi-Nagy, Anna; Mallick, Swapan; Pósa, Annamária; Stégmár, Balázs; Keerl, Victoria; Rohland, Nadin; Stewardson, Kristin; Ferry, Matthew; Michel, Megan; Oppenheimer, Jonas; Broomandkhoshbacht, Nasreen; Harney, Eadaoin; Nordenfelt, Susanne; Llamas, Bastien; Gusztáv Mende, Balázs; Köhler, Kitti; Oross, Krisztián; Bondár, Mária; Marton, Tibor; Osztás, Anett; Jakucs, János; Paluch, Tibor; Horváth, Ferenc; Csengeri, Piroska; Koós, Judit; Sebők, Katalin; Anders, Alexandra; Raczky, Pál; Regenye, Judit; Barna, Judit P.; Fábián, Szilvia; Serlegi, Gábor; Toldi, Zoltán; Gyöngyvér Nagy, Emese; Dani, János; Molnár, Erika; Pálfi, György; Márk, László; Melegh, Béla; Bánfai, Zsolt; Domboróczki, László; Fernández-Eraso, Javier; Antonio Mujika-Alustiza, José; Alonso Fernández, Carmen; Jiménez Echevarría, Javier; Bollongino, Ruth; Orschiedt, Jörg; Schierhold, Kerstin; Meller, Harald; Cooper, Alan; Burger, Joachim; Bánffy, Eszter; Alt, Kurt W.; Lalueza-Fox, Carles; Haak, Wolfgang; Reich, David (8 November 2017). "Parallel palaeogenomic transects reveal complex genetic history of early European farmers". Nature. 551 (7680): 368–372. Bibcode:2017Natur.551..368L. doi:10.1038/nature24476. PMC 5973800. PMID 29144465.
- Kılınç, Gülşah Merve; Omrak, Ayça; Özer, Füsun; Günther, Torsten; Büyükkarakaya, Ali Metin; Bıçakçı, Erhan; Baird, Douglas; Dönertaş, Handan Melike; Ghalichi, Ayshin; Yaka, Reyhan; Koptekin, Dilek; Açan, Sinan Can; Parvizi, Poorya; Krzewińska, Maja; Daskalaki, Evangelia A.; Yüncü, Eren; Dağtaş, Nihan Dilşad; Fairbairn, Andrew; Pearson, Jessica; Mustafaoğlu, Gökhan; Erdal, Yılmaz Selim; Çakan, Yasin Gökhan; Togan, İnci; Somel, Mehmet; Storå, Jan; Jakobsson, Mattias; Götherström, Anders (10 October 2016). "The Demographic Development of the First Farmers in Anatolia". Current Biology. 26 (19): 2659–2666. doi:10.1016/j.cub.2016.07.057. PMC 5069350. PMID 27498567. -
- Lippold, Sebastian; Xu, Hongyang; Ko, Albert; Li, Mingkun; Renaud, Gabriel; Butthof, Anne; Schröder, Roland; Stoneking, Mark (2014). "Human paternal and maternal demographic histories: insights from high-resolution Y chromosome and mtDNA sequences". Investigative Genetics. 5: 13. bioRxiv 10.1101/001792. doi:10.1186/2041-2223-5-13. PMC 4174254. PMID 25254093.
- "Haplogroups I & N". Ianlogan.co.uk. Retrieved 2010-07-29.
- Turchi, Chiara; Buscemi, Loredana; Previderè, Carlo; Grignani, Pierangela; Brandstätter, Anita; Achilli, Alessandro; Parson, Walther; Tagliabracci, Adriano; Ge.f.i., Group (2007). "Italian mitochondrial DNA database: results of a collaborative exercise and proficiency testing". International Journal of Legal Medicine. 122 (3): 199–204. doi:10.1007/s00414-007-0207-1. PMID 17952451. S2CID 11496367.
- "Haplogroup W". Ianlogan.co.uk. Retrieved 2010-07-29.
- Metspalu M, Kivisild T, Metspalu E, et al. (2004). "Most of the extant mtDNA boundaries in south and southwest Asia were likely shaped during the initial settlement of Eurasia by anatomically modern humans". BMC Genetics. 5: 26. doi:10.1186/1471-2156-5-26. PMC 516768. PMID 15339343.
- Kong, Qing-Peng et al. 2011, Large-Scale mtDNA Screening Reveals a Surprising Matrilineal Complexity in East Asia and Its Implications to the Peopling of the Region.
- Behar et al., 2012b
- Soanboon, Pattanawit; Nanakorn, Somsong; Kutanan, Wibhu (June 2016). "Determination of sex difference from fingerprint ridge density in northeastern Thai teenagers". Egyptian Journal of Forensic Sciences. 6 (2): 185–193. doi:10.1016/j.ejfs.2015.08.001.
- YFull Haplogroup YTree v6.05.11 at 25 September 2018.
- Brandão, Andreia; Eng, Ken Khong; Rito, Teresa; Cavadas, Bruno; Bulbeck, David; Gandini, Francesca; Pala, Maria; Mormina, Maru; Hudson, Bob; White, Joyce; Ko, Tsang-Ming; Saidin, Mokhtar; Zafarina, Zainuddin; Oppenheimer, Stephen; Richards, Martin B.; Pereira, Luísa; Soares, Pedro (2016). "Quantifying the legacy of the Chinese Neolithic on the maternal genetic heritage of Taiwan and Island Southeast Asia". Human Genetics. 135 (4): 363–376. doi:10.1007/s00439-016-1640-3. PMC 4796337. PMID 26875094.
- Peng, Min-Sheng; Xu, Weifang; Song, Jiao-Jiao; Chen, Xing; Sulaiman, Xierzhatijiang; Cai, Liuhong; Liu, He-Qun; Wu, Shi-Fang; Gao, Yun; Abdulloevich, Najmudinov Tojiddin; Afanasevna, Manilova Elena; Ibrohimovich, Khudoidodov Behruz; Chen, Xi; Yang, Wei-Kang; Wu, Miao; Li, Gui-Mei; Yang, Xing-Yan; Rakha, Allah; Yao, Yong-Gang; Upur, Halmurat; Zhang, Ya-Ping (29 November 2017). "Mitochondrial genomes uncover the maternal history of the Pamir populations". European Journal of Human Genetics. 26 (1): 124–136. doi:10.1038/s41431-017-0028-8. PMC 5839027. PMID 29187735.
- Derenko, Miroslava; Malyarchuk, Boris; Denisova, Galina; Perkova, Maria; Rogalla, Urszula; Grzybowski, Tomasz; Khusnutdinova, Elza; Dambueva, Irina; Zakharov, Ilia (21 February 2012). "Complete Mitochondrial DNA Analysis of Eastern Eurasian Haplogroups Rarely Found in Populations of Northern Asia and Eastern Europe". PLOS ONE. 7 (2): e32179. Bibcode:2012PLoSO...732179D. doi:10.1371/journal.pone.0032179. PMC 3283723. PMID 22363811.
- Hwan Young Lee, Ji-Eun Yoo, Myung Jin Park, Ukhee Chung, Chong-Youl Kim, and Kyoung-Jin Shin, "East Asian mtDNA haplogroup determination in Koreans: Haplogroup-level coding region SNP analysis and subhaplogroup-level control region sequence analysis." Electrophoresis (2006). DOI 10.1002/elps.200600151.
- Duong, Nguyen Thuy; Macholdt, Enrico; Ton, Nguyen Dang; Arias, Leonardo; Schröder, Roland; Van Phong, Nguyen; Thi Bich Thuy, Vo; Ha, Nguyen Hai; Thi Thu Hue, Huynh; Thi Xuan, Nguyen; Thi Phuong Oanh, Kim; Hien, Le Thi Thu; Hoang, Nguyen Huy; Pakendorf, Brigitte; Stoneking, Mark; Van Hai, Nong (3 August 2018). "Complete human mtDNA genome sequences from Vietnam and the phylogeography of Mainland Southeast Asia". Scientific Reports. 8 (1): 11651. Bibcode:2018NatSR...811651D. doi:10.1038/s41598-018-29989-0. PMC 6076260. PMID 30076323.
- "Haplogroup Y". Ianlogan.co.uk. 2009-09-22. Retrieved 2010-07-29.
- Wibhu Kutanan, Rasmi Shoocongdej, Metawee Srikummool, et al. (2020), "Cultural variation impacts paternal and maternal genetic lineages of the Hmong-Mien and Sino-Tibetan groups from Thailand." European Journal of Human Genetics. https://doi.org/10.1038/s41431-020-0693-x
- Y-DNA D Haplogroup Project at Family Tree DNA
- Hongbin Yao, Mengge Wang, Xing Zou, et al., "New insights into the fine-scale history of western-eastern admixture of the northwestern Chinese population in the Hexi Corridor via genome-wide genetic legacy." Mol Genet Genomics 2021 Mar 1. doi: 10.1007/s00438-021-01767-0.
- Gunnarsdóttir, Ellen et al. 2010, High-throughput sequencing of complete human mtDNA genomes from the Philippines
- "Hudjashov". Ianlogan.co.uk. Retrieved 2010-07-29.
- Pierson MJ, Martinez-Arias R, Holland BR, Gemmell NJ, Hurles ME, Penny D (2006). "Deciphering past human population movements in Oceania: provably optimal trees of 127 mtDNA genomes". Molecular Biology and Evolution. 23 (10): 1966–75. doi:10.1093/molbev/msl063. PMC 2674580. PMID 16855009.
- Hill, C.; Soares, P.; Mormina, M.; MacAulay, V.; Meehan, W.; Blackburn, J.; Clarke, D.; Raja, J. M.; Ismail, P.; Bulbeck, D.; Oppenheimer, S.; Richards, M. (2006). "Phylogeography and Ethnogenesis of Aboriginal Southeast Asians". Molecular Biology and Evolution. 23 (12): 2480–91. doi:10.1093/molbev/msl124. PMID 16982817.
- Rieux, Adrien; Eriksson, Anders; Li, Mingkun; et al. (2014). "Improved Calibration of the Human Mitochondrial Clock Using Ancient Genomes". Mol Biol Evol. 31 (10): 2780–92. doi:10.1093/molbev/msu222. PMC 4166928. PMID 25100861.
- "Haplogroup A". Ianlogan.co.uk. Retrieved 2010-07-29.
- "Haplogroup S". Ianlogan.co.uk. Retrieved 2010-07-29.
- "Haplogroup X". Ianlogan.co.uk. Retrieved 2010-07-29.
- "Haplogroup R*". Ianlogan.co.uk. Retrieved 2010-07-29.
External links
- Haplogroup N
- Mannis van Oven's – mtDNA subtree N
- Spread of Haplogroup N, from National Geographic
- Hudjashov, G.; Kivisild, T.; Underhill, P. A.; Endicott, P.; Sanchez, J. J.; Lin, A. A.; Shen, P.; Oefner, P.; Renfrew, C.; Villems, R.; Forster, P. (2007). "Revealing the prehistoric settlement of Australia by Y chromosome and mtDNA analysis". Proceedings of the National Academy of Sciences. 104 (21): 8726–30. Bibcode:2007PNAS..104.8726H. doi:10.1073/pnas.0702928104. PMC 1885570. PMID 17496137.
- Katherine Borges' The Haplogroup N mtDNA Study at Family Tree DNA
- General
- Ian Logan's Mitochondrial DNA Site