Hydrogen potassium ATPase
Gastric hydrogen potassium ATPase, also known as H+/K+ ATPase, is an enzyme which functions to acidify the stomach.[1] It is a member of the P-type ATPases, also known as E1-E2 ATPases due to its two states.[2]
| ATPase, H+/K+ exchanging, alpha polypeptide | |||||||
|---|---|---|---|---|---|---|---|
| Identifiers | |||||||
| Symbol | ATP4A | ||||||
| NCBI gene | 495 | ||||||
| HGNC | 819 | ||||||
| OMIM | 137216 | ||||||
| RefSeq | NM_000704 | ||||||
| UniProt | P20648 | ||||||
| Other data | |||||||
| EC number | 7.2.2.19 | ||||||
| Locus | Chr. 19 q13.1 | ||||||
| |||||||
| ATPase, H+/K+ exchanging, beta polypeptide | |||||||
|---|---|---|---|---|---|---|---|
| Identifiers | |||||||
| Symbol | ATP4B | ||||||
| NCBI gene | 496 | ||||||
| HGNC | 820 | ||||||
| OMIM | 137217 | ||||||
| RefSeq | NM_000705 | ||||||
| UniProt | P51164 | ||||||
| Other data | |||||||
| EC number | 7.2.2.19 | ||||||
| Locus | Chr. 13 q34 | ||||||
| |||||||
Biological function and location
The gastric hydrogen potassium ATPase or H+/K+ ATPase is the proton pump of the stomach. It exchanges potassium from the intestinal lumen with cytoplasmic hydronium[2] and is the enzyme primarily responsible for the acidification of the stomach contents and the activation of the digestive enzyme pepsin[3] (see gastric acid).
The H+/K+ ATPase is found in parietal cells, which are highly specialized epithelial cells located in the inner cell lining of the stomach called the gastric mucosa. Parietal cells possess an extensive secretory membrane system and the H+/K+ ATPase is the major protein constituent of these membranes. A small amount of H+/K+ ATPase is also found in the renal medulla.[2]
Genes and protein structure
The H+/K+ ATPase is a heterodimeric protein, the product of 2 genes. The gene ATP4A[4] encodes the H+/K+ ATPase α subunit, and is a ~1000-amino acid protein that contains the catalytic sites of the enzyme and forms the pore through the cell membrane that allows the transport of ions. Hydronium ions bind to two active sites present in the α subunit.[5] The α subunit also has a phosphorylation site (Asp385).[6] The gene ATP4B[7] encodes the β subunit of the H+/K+ ATPase, which is a ~300-amino acid protein with a 36-amino acid N-terminal cytoplasmic domain, a single transmembrane domain, and a highly glycosylated extracellular domain.
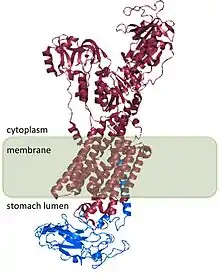
The H+/K+ ATPase β subunit stabilizes the H+/K+ ATPase α subunit and is required for function of the enzyme. The β subunit prevents the pump from running in reverse,[8] and it also appears to contain signals that direct the heterodimer to membrane destinations within the cell, although some of these signals are subordinate to signals found in H+/K+ ATPase α subunit.
The structure of H+/K+ ATPase has been determined for humans, dogs, hogs, rats, and rabbits and is 98% homologous across all species.[2]
Enzyme mechanism and activity
H+/K+ ATPase is a P2-type ATPase, a member of the eukaryotic class of P-type ATPases.[9] Like the Ca2+ and the Na+/K+ ATPases, the H+/K+ ATPase functions as an α, β protomer.[10] Unlike other eukaryotic ATPases, the H+/K+ ATPase is electroneutral, transporting one proton into the stomach lumen per potassium ion retrieved from the gastric lumen.[9] As an ion pump the H+/K+ ATPase is able to transport ions against a concentration gradient using energy derived from the hydrolysis of ATP. Like all P-type ATPases, a phosphate group is transferred from adenosine triphosphate (ATP) to the H+/K+ ATPase during the transport cycle. This phosphate transfer powers a conformational change in the enzyme that helps drive ion transport.
The hydrogen potassium ATPase is activated indirectly by gastrin that causes ECL cells to release histamine.[11] The histamine binds to H2 receptors on the parietal cell, activating a cAMP-dependent pathway which causes the enzyme to move from the cytoplasmic tubular membranes to deeply folded canaliculi of the stimulated parietal cell.[2] Once localized, the enzyme alternates between two conformations, E1 and E2, to transport ions across the membrane.

The E1 conformation binds a phosphate from ATP and hydronium ion on the cytoplasmic side. The enzyme then changes to the E2 conformation, allowing hydronium to be released in the lumen. The E2 conformation binds potassium, and reverts to the E1 conformation to release phosphate and K+ into the cytoplasm where another ATP can be hydrolyzed to repeat the cycle.[2] The β subunit prevents the E2-P conformation from reverting to the E1-P conformation, making proton pumping unidirectional.[8] The number of ions transported per ATP varies from 2H+/2K+ to 1H+/1K+depending on the pH of the stomach.[12]
Disease relevance and inhibition
Inhibiting the hydrogen potassium pump to decrease stomach acidity has been the most common method of treating diseases including gastroesophageal reflux disease (GERD/GORD) and peptic ulcer disease (PUD).[13] Reducing acidity alleviates disease symptoms but does not treat the actual cause of GERD (abnormal relaxation of the esophageal sphincter) or PUD (Helicobacter pylori and NSAIDs).[14]
Three drug classes have been used to inhibit H+/K+-ATPases. H2-receptor antagonists, like cimetidine (Tagamet), inhibit the signaling pathway that leads to activation of the ATPase. This type of inhibitor is effective in treating ulcers but does not prevent them from forming, and patients develop tolerance to them after about one week, leading to a 50% reduction in efficacy.[15] Proton pump inhibitors (PPIs) were later developed, starting with Timoprazole in 1975.[15] PPIs are acid-activated prodrugs that inhibit the hydrogen-potassium ATPase by binding covalently to active pumps.[16] Current PPIs like Omeprazole have a short half-life of approximately 90 minutes.[17] Acid pump antagonists (APAs) or potassium-competitive acid blockers (PCABs) are a third type of inhibitor that blocks acid secretion by binding to the K+ active site.[15] APAs provide faster inhibition than PPIs since they do not require acid activation. Revaprazan was the first APA used clinically in east Asia, and other APAs are being developed since they appear to provide better acid control in clinical trials.[17]
Inactivation of the proton pump can also lead to health problems. A study in mice by Krieg et al.[18] found that a mutation of the pump’s α-subunit led to achlorhydria, resulting in problems with iron absorption, leading to iron deficiency and anemia. The use of PPIs has not been correlated with an elevated risk of anemia, so the H+/K+-ATPase is thought to aid iron absorption but is not necessarily required.[18]
Current association of dementia and PPIs have been documented in Germany and in research articles denoting how Benzimidazole derivatives, Astemizole (AST) and Lansoprazole (LNS) interact with anomalous aggregates of tau protein (neurofibrillary tangles).[19][20][21] Current theories include the non-selective blockade of sodium-potassium pumps in the brain causing osmotic imbalances or swelling in the cells. [auth opinion] Interaction of PPIs with other drug affecting the sodium-potassium pump, e.g., digoxin, warfarin etc., has been well documented.[22] Memory has been associated with astrocytes and the alpha3 subunit of adenosine receptor found in hydrogen/Sodium-potassium pumps may be a focal point in dementia.[23][24][25] Chronic use of PPIs may cause down regulation of alpha3 subunit increasing damage to astrocytes.[26] Osteopetrosis via TCIRG1 gene has a strong association with pre-senile dementia.[27][28]
References
- Sakai, Hideki; Fujii, Takuto; Takeguchi, Noriaki (2016). "Chapter 13. Proton-Potassium (H+/K+) ATPases: Properties and Roles in Health and Diseases". In Astrid, Sigel; Helmut, Sigel; Roland K.O., Sigel (eds.). The Alkali Metal Ions: Their Role in Life. Metal Ions in Life Sciences. Vol. 16. Springer. pp. 459–483. doi:10.1007/978-3-319-21756-7_13. PMID 26860309.
- Shin, J. M.; Munson, K.; Vagin, O.; Sachs, G. (2009). "The gastric HK-ATPase: structure, function and inhibition". Pflügers Archiv: European Journal of Physiology. 457 (3): 609–622. doi:10.1007/s00424-008-0495-4. PMC 3079481. PMID 18536934.
- Berg, J. M.; Tymoczko, J. L.; Stryer, L. (2012). Biochemistry (7th ed.). New York: W.H. Freeman and Company.
- ATP4A ATPase H+/K+ transporting alpha subunit
- Chourasia, M.; Sastry, G. M.; Sastry. G. N. (2005). "Proton binding sites and conformational analysis of H+K+-ATPase". Biochemical and Biophysical Research Communications. 336 (3): 961–966. doi:10.1016/j.bbrc.2005.08.205. PMID 16157306.
- Scheirlinckx, F.; Raussens, V.; Ruysschaert, J.-M.; Goormaghtigh, E. (2004). "Conformational changes in gastric H+/K+-ATPase monitored by difference Fourier-transform infrared spectroscopy and hydrogen/deuterium exchange". Biochemical Journal (Pt 1 ed.). 382 (Pt 1): 121–129. doi:10.1042/BJ20040277. PMC 1133922. PMID 15096097.
- ATP4B ATPase H+/K+ transporting beta subunit
- Abe, K.; Tani, K.; Nishizawa, T.; Fujiyoshi, Y. (2009). "Inter-subunit interaction of gastric H+,K+-ATPase prevents reverse reaction of the transport cycle". The EMBO Journal. 28 (11): 1637–1643. doi:10.1038/emboj.2009.102. PMC 2693145. PMID 19387495.
- Shin, J. M.; Sachs, G. (2009). "Ion Motive ATPases: P-type ATPases". eLS. doi:10.1002/9780470015902.a0001379.pub2. ISBN 978-0470016176.
- Dach, I.; Olesen, C.; Signor, L.; Nissen, P.; le Maire, M.; Møller, J. V.; Ebel, C. (2012). "Active Detergent-solubilized H+,K+-ATPase Is a Monomer". The Journal of Biological Chemistry. 287 (50): 41963–41978. doi:10.1074/jbc.M112.398768. PMC 3516743. PMID 23055529.
- Prinz, C.; Kajimura, M.; Scott, D.; Helander, H.; Shin, J.; Besancon, M.; Bamberg, K.; Hersey, S.; Sachs, G. (1992). "Acid secretion and the H,K ATPase of stomach". The Yale Journal of Biology and Medicine. 65 (6): 577–596. PMC 2589780. PMID 1341065.
- Abe, K.; Tani, K.; Friedrich, T.; Fujiyoshi, Y. (2012). "Cryo-EM structure of gastric H+,K+-ATPase with a single occupied cation-binding site". Proceedings of the National Academy of Sciences of the United States of America. 109 (45): 18401–18406. Bibcode:2012PNAS..10918401A. doi:10.1073/pnas.1212294109. PMC 3494912. PMID 23091039.
- Shin, J. M.; Vagin, O.; Munson, K.; Kidd, M.; Modlin, I. M.; Sachs, G. (2008). "Molecular mechanisms in therapy of acid-related diseases". Cellular and Molecular Life Sciences. 65 (2): 264–281. doi:10.1007/s00018-007-7249-x. PMC 3081136. PMID 17928953.
- Yeomans, N. D. (2011). "The ulcer sleuths: The search for the cause of peptic ulcers". Journal of Gastroenterology and Hepatology. 26: 35–41. doi:10.1111/j.1440-1746.2010.06537.x. PMID 21199512. S2CID 42592868.
- Sachs, G.; Shin, J. M.; Vagin, O.; Lambrecht, N.; Yakubov, I.; Munson, K. (2007). "The Gastric H,K ATPase as a Drug Target: Past, Present and Future". Journal of Clinical Gastroenterology. 41 (Suppl 2): S226–S242. doi:10.1097/MCG.0b013e31803233b7. PMC 2860960. PMID 17575528.
- Shin J. M.; Sachs G. (2008). "Pharmacology of Proton Pump Inhibitors". Current Gastroenterology Reports. 10 (6): 528–534. doi:10.1007/s11894-008-0098-4. PMC 2855237. PMID 19006606.
- Shin, J. M.; Sachs, G. (2009). "Long lasting inhibitors of the gastric H,K-ATPase". Expert Review of Clinical Pharmacology. 2 (5): 461–468. doi:10.1586/ecp.09.33. PMC 2995460. PMID 21132072.
- Krieg L.; Milstein O.; Krebs, P.; Xia, Y.; Beutler B.; Du, X. (2011). "Mutation of the gastric hydrogen-potassium ATPase alpha subunit causes iron-deficiency anemia in mice". Blood. 118 (24): 6418–6425. doi:10.1182/blood-2011-04-350082. PMC 3236123. PMID 21976678.
- Gomm W, von Holt K, Thomé F, Broich K, Maier W, Fink A, Doblhammer G, Haenisch B (2016). "Association of Proton Pump Inhibitors With Risk of Dementia: A Pharmacoepidemiological Claims Data Analysis". JAMA Neurol. 73 (4): 410–416. doi:10.1001/jamaneurol.2015.4791. PMID 26882076.
- Rojo LE, Alzate-Morales J, Saavedra IN, Davies P, Maccioni RB (2010). "Selective interaction of lansoprazole and astemizole with tau polymers: potential new clinical use in diagnosis of Alzheimer's disease". J Alzheimers Dis. 19 (2): 573–89. doi:10.3233/JAD-2010-1262. PMC 2951486. PMID 20110603.
- Fawaz MV, Brooks AF, Rodnick ME, Carpenter GM, Shao X, Desmond TJ, Sherman P, Quesada CA, Hockley BG, Kilbourn MR, Albin RL, Frey KA, Scott PJ (2014). "High affinity radiopharmaceuticals based upon lansoprazole for PET imaging of aggregated tau in Alzheimer's disease and progressive supranuclear palsy: synthesis, preclinical evaluation, and lead selection". ACS Chem Neurosci. 5 (8): 718–30. doi:10.1021/cn500103u. PMC 4140593. PMID 24896980. Epub 2014 Jun 16.
- Trifirò G, Corrao S, Alacqua M, Moretti S, Tari M, Caputi AP, Arcoraci V (2006). "Interaction risk with proton pump inhibitors in general practice: significant disagreement between different drug-related information sources". Br J Clin Pharmacol. 62 (5): 582–90. doi:10.1111/j.1365-2125.2006.02687.x. PMC 1885178. PMID 16822281.
- Björklund O, Shang M, Tonazzini I, Daré E, Fredholm BB (2008). "Adenosine A1 and A3 receptors protect astrocytes from hypoxic damage". Eur J Pharmacol. 596 (1–3): 6–13. doi:10.1016/j.ejphar.2008.08.002. PMID 18727925. Epub 2008 Aug 13.
- Carmona MA, Murai KK, Wang L, Roberts AJ, Pasquale EB (2009). "Glial ephrin-A3 regulates hippocampal dendritic spine morphology and glutamate transport". Proc Natl Acad Sci U S A. 106 (30): 12524–12529. Bibcode:2009PNAS..10612524C. doi:10.1073/pnas.0903328106. PMC 2718351. PMID 19592509.
- Ben Haim L, Carrillo-de Sauvage MA, Ceyzériat K, Escartin C (2015). "Elusive roles for reactive astrocytes in neurodegenerative diseases". Front Cell Neurosci. 9: 278. doi:10.3389/fncel.2015.00278. PMC 4522610. PMID 26283915. eCollection 2015
- Gessi S, Merighi S, Stefanelli A, Fazzi D, Varani K, Borea PA (2013). "A(1) and A(3) adenosine receptors inhibit LPS-induced hypoxia-inducible factor-1 accumulation in murine astrocytes". Pharmacol Res. 76: 157–70. doi:10.1016/j.phrs.2013.08.002. PMID 23969284. Epub 2013 Aug 19.
- "TCIRG1". Genetics Home Reference.
- Kaifu T, Nakahara J, Inui M, Mishima K, Momiyama T, Kaji M, Sugahara A, Koito H, Ujike-Asai A, Nakamura A, Kanazawa K, Tan-Takeuchi K, Iwasaki K, Yokoyama WM, Kudo A, Fujiwara M, Asou H, Takai T (2003). "Osteopetrosis and thalamic hypomyelinosis with synaptic degeneration in DAP12-deficient mice". J Clin Invest. 111 (3): 323–32. doi:10.1172/JCI16923. PMC 151867. PMID 12569157.
External links
- Yao X, Forte JG (2003). "Cell biology of acid secretion by the parietal cell". Annu. Rev. Physiol. 65: 103–31. doi:10.1146/annurev.physiol.65.072302.114200. PMID 12500969.
- Kühlbrandt W (April 2004). "Biology, structure and mechanism of P-type ATPases". Nat. Rev. Mol. Cell Biol. 5 (4): 282–95. doi:10.1038/nrm1354. PMID 15071553. S2CID 24927167.
- Dunbar LA, Caplan MJ (August 2001). "Ion pumps in polarized cells: sorting and regulation of the Na+, K+- and H+, K+-ATPases". J. Biol. Chem. 276 (32): 29617–20. doi:10.1074/jbc.R100023200. PMID 11404365.
- Sachs G, Shin JM, Briving C, Wallmark B, Hersey S (1995). "The pharmacology of the gastric acid pump: the H+,K+ ATPase". Annu. Rev. Pharmacol. Toxicol. 35: 277–305. doi:10.1146/annurev.pa.35.040195.001425. PMID 7598495.
- Potassium+Hydrogen+ATPase at the U.S. National Library of Medicine Medical Subject Headings (MeSH)