Cichlid
Cichlids /ˈsɪklɪdz/[lower-alpha 1] are fish from the family Cichlidae in the order Cichliformes. Traditionally Cichlids were classed in a suborder, the Labroidei, along with the wrasses (Labridae), in the order Perciformes,[1] but molecular studies have contradicted this grouping.[2] On the basis of fossil evidence, it first appeared in Tanzania during the Eocene epoch, about 46–45 million years ago.[3][4] The closest living relative of cichlids is probably the convict blenny, and both families are classified in the 5th edition of Fishes of the World as the two families in the Cichliformes, part of the subseries Ovalentaria.[5] This family is large, diverse, and widely dispersed. At least 1,650 species have been scientifically described,[6] making it one of the largest vertebrate families. New species are discovered annually, and many species remain undescribed. The actual number of species is therefore unknown, with estimates varying between 2,000 and 3,000.[7]
| Cichlid | |
|---|---|
 | |
| A mbuna | |
| Scientific classification | |
| Domain: | Eukaryota |
| Kingdom: | Animalia |
| Phylum: | Chordata |
| Class: | Actinopterygii |
| Clade: | Percomorpha |
| (unranked): | Ovalentaria |
| Order: | Cichliformes |
| Family: | Cichlidae Bonaparte, 1835 |
| Subfamilies and Tribes | |
Alternate taxonomy:
| |
Many cichlids, particularly tilapia, are important food fishes, while others, such as the Cichla species, are valued game fish. The family also includes many popular freshwater aquarium fish kept by hobbyists, including the angelfish, oscars, and discus.[8][9] Cichlids have the largest number of endangered species among vertebrate families, most in the haplochromine group.[10] Cichlids are particularly well known for having evolved rapidly into many closely related but morphologically diverse species within large lakes, particularly Lakes Tanganyika, Victoria, Malawi, and Edward.[11][12] Their diversity in the African Great Lakes is important for the study of speciation in evolution.[13] Many cichlids introduced into waters outside of their natural range have become nuisances.[14]
All cichlids practice some form of parental care for their eggs and fry, usually in the form of guarding the eggs and fry or mouthbrooding.
Anatomy and appearance
Cichlids span a wide range of body sizes, from species as small as 2.5 cm (1 in) in length (e.g., female Neolamprologus multifasciatus) to much larger species approaching 1 m (3 ft) in length (Boulengerochromis and Cichla). As a group, cichlids exhibit a similar diversity of body shapes, ranging from strongly laterally compressed species (such as Altolamprologus, Pterophyllum, and Symphysodon) to species that are cylindrical and highly elongated (such as Julidochromis, Teleogramma, Teleocichla, Crenicichla, and Gobiocichla).[8] Generally, however, cichlids tend to be of medium size, ovate in shape, and slightly laterally compressed, and generally similar to the North American sunfishes in morphology, behavior, and ecology.[15]
Cichlids share a single key trait - the fusion of the lower pharyngeal bones into a single tooth-bearing structure. A complex set of muscles allows the upper and lower pharyngeal bones to be used as a second set of jaws for processing food, allowing a division of labor between the "true jaws" (mandibles) and the "pharyngeal jaws". Cichlids are efficient and often highly specialized feeders that capture and process a very wide variety of food items. This is assumed to be one reason why they are so diverse.[8]
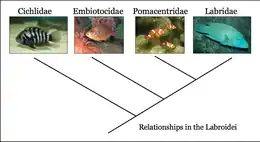
The features that distinguish them from the other families in the Labroidei include:[16]
- A single nostril on each side of the forehead, instead of two
- No bony shelf below the orbit of the eye
- Division of the lateral line organ into two sections, one on the upper half of the flank and a second along the midline of the flank from about halfway along the body to the base of the tail (except for genera Teleogramma and Gobiocichla)
- A distinctively shaped otolith
- The small intestine's left-side exit from the stomach instead of its right side as in other Labroidei
Taxonomy
Kullander (1998) recognizes eight subfamilies of cichlids: the Astronotinae, Cichlasomatinae, Cichlinae, Etroplinae, Geophaginae, Heterochromidinae, Pseudocrenilabrinae, and Retroculinae.[17] A ninth subfamily, the Ptychochrominae, was later recognized by Sparks and Smith.[18] Cichlid taxonomy is still debated, and classification of genera cannot yet be definitively given. A comprehensive system of assigning species to monophyletic genera is still lacking, and there is not complete agreement on what genera should be recognized in this family.[19]
As an example of the classification problems, Kullander[20] placed the African genus Heterochromis phylogenetically within Neotropical cichlids, although later papers concluded otherwise. Other problems center upon the identity of the putative common ancestor for the Lake Victoria superflock (many closely related species sharing a single habitat), and the ancestral lineages of Lake Tanganyikan cichlids.

Phylogeny derived from morphological characters shows differences at the genus level with phylogeny based on genetic loci.[21] A consensus remains that the Cichlidae as a family are monophyletic.[22]
In cichlid taxonomy, dentition was formerly used as a classifying characteristic, but this was complicated because in many cichlids, tooth shapes change with age, due to wear, and cannot be relied upon. Genome sequencing and other technologies transformed cichlid taxonomy.
Alternatively, all cichlid species native to the new world, can be classified under the subfamily Cichlinae, while Etroplinae can classify all cichlid species native to the old world.
Distribution and habitat

Cichlids are one of the largest vertebrate families in the world. They are most diverse in Africa and South America. Africa alone is host to at least an estimated 1,600 species.[19] Central America and Mexico have about 120 species, as far north as the Rio Grande in South Texas. Madagascar has its own distinctive species (Katria, Oxylapia, Paratilapia, Paretroplus, Ptychochromis, and Ptychochromoides), only distantly related to those on the African mainland.[16][24] Native cichlids are largely absent in Asia, except for 9 species in Israel, Lebanon, and Syria (Astatotilapia flaviijosephi, Oreochromis aureus, O. niloticus, Sarotherodon galilaeus, Coptodon zillii, and Tristramella spp.), two in Iran (Iranocichla), and three in India and Sri Lanka (Etroplus and Pseudetroplus).[19] If disregarding Trinidad and Tobago (where the few native cichlids are members of genera that are widespread in the South American mainland), the three species from the genus Nandopsis are the only cichlids from the Antilles in the Caribbean, specifically Cuba and Hispaniola. Europe, Australia, Antarctica, and North America north of the Rio Grande drainage have no native cichlids, although in Florida, Hawaii, Japan, northern Australia, and elsewhere, feral populations of cichlids have become established as exotics.[23][25][26][27][28][29][30]
Although most cichlids are found at relatively shallow depths, several exceptions do exist. The deepest known occurrences are Trematocara at more than 300 m (1,000 ft) below the surface in Lake Tanganyika.[31] Others found in relatively deep waters include species such as Alticorpus macrocleithrum and Pallidochromis tokolosh down to 150 m (500 ft) below the surface in Lake Malawi,[32][33] and the whitish (nonpigmented) and blind Lamprologus lethops, which is believed to live as deep as 160 m (520 ft) below the surface in the Congo River.[34]
Cichlids are less commonly found in brackish and saltwater habitats, though many species tolerate brackish water for extended periods; Mayaheros urophthalmus, for example, is equally at home in freshwater marshes and mangrove swamps, and lives and breeds in saltwater environments such as the mangrove belts around barrier islands.[8] Several species of Tilapia, Sarotherodon, and Oreochromis are euryhaline and can disperse along brackish coastlines between rivers.[19] Only a few cichlids, however, inhabit primarily brackish or salt water, most notably Etroplus maculatus, Etroplus suratensis, and Sarotherodon melanotheron.[35] The perhaps most extreme habitats for cichlids are the warm hypersaline lakes where the members of the genera Alcolapia and Danakilia are found. Lake Abaeded in Eritrea encompasses the entire distribution of D. dinicolai, and its temperature ranges from 29 to 45 °C (84 to 113 °F).[36]
With the exception of the species from Cuba, Hispaniola, and Madagascar, cichlids have not reached any oceanic island and have a predominantly Gondwanan distribution, showing the precise sister relationships predicted by vicariance: Africa-South America and India-Madagascar.[37] The dispersal hypothesis, in contrast, requires cichlids to have negotiated thousands of kilometers of open ocean between India and Madagascar without colonizing any other island, or for that matter, crossing the Mozambique Channel to Africa. Although the vast majority of Malagasy cichlids are entirely restricted to fresh water, Ptychochromis grandidieri and Paretroplus polyactis are commonly found in coastal brackish water and are apparently salt tolerant,[38][39] as is also the case for Etroplus maculatus and E. suratensis from India and Sri Lanka.[40][41]
Ecology
Feeding
Within the cichlid family, carnivores, herbivores, omnivores, planktivores, and detritivores are known, meaning the Cichlidae encompass essentially the full range of food consumption possible in the animal kingdom. Various species have morphological adaptations for specific food sources,[42] but most cichlids consume a wider variety of foods based on availability. Carnivorous cichlids can be further divided into piscivorous and molluscivorous, since the morphology and hunting behavior differ greatly between the two categories. Piscivorous cichlids eat other fish, fry, larvae, and eggs. Some species eat the offspring of mouthbrooders by head-ramming, wherein the hunter shoves its head into the mouth of a female to expel her young and eat them.[43] Molluscivorous cichlids have several hunting strategies amongst the varieties within the group. Lake Malawi cichlids consume substrate and filter it out through their gill rakers to eat the mollusks that were in the substrate. Gill rakers are finger-like structures that line the gills of some fish to catch any food that might escape through their gills.[44]

Many cichlids are primarily herbivores, feeding on algae (e.g. Petrochromis) and plants (e.g. Etroplus suratensis). Small animals, particularly invertebrates, are only a minor part of their diets.
Other cichlids are detritivores and eat organic material, called Aufwuchs (offal); among these species are the tilapiines of the genera Oreochromis, Sarotherodon, and Tilapia.
Other cichlids are predatory and eat little or no plant matter. These include generalists that catch a variety of small animals, including other fishes and insect larvae (e.g. Pterophyllum), as well as variety of specialists. Trematocranus is a specialized snail-eater, while Pungu maclareni feeds on sponges. A number of cichlids feed on other fish, either entirely or in part. Crenicichla species are stealth predators that lunge from concealment at passing small fish, while Rhamphochromis species are open-water pursuit predators that chase down their prey.[46] Paedophagous cichlids such as the Caprichromis species eat other species' eggs or young, in some cases ramming the heads of mouthbrooding species to force them to disgorge their young.[47][48][49][50] Among the more unusual feeding strategies are those of Corematodus, Docimodus evelynae, Plecodus, Perissodus, and Genyochromis spp., which feed on scales and fins of other fishes, a behavior known as lepidophagy,[51][52][53] along with the death-mimicking behaviour of Nimbochromis and Parachromis species, which lay motionless, luring small fish to their side prior to ambush.[54][55]
This variety of feeding styles has helped cichlids to inhabit similarly varied habitats. Its pharyngeal teeth (in the throat) afford cichlids so many "niche" feeding strategies, because the jaws pick and hold food, while the pharyngeal teeth crush the prey.
Behavior
Aggression
Aggressive behavior in cichlids is ritualized and consists of multiple displays used to seek confrontation while being involved in evaluation of competitors,[56] coinciding with temporal proximity to mating. Displays of ritualized aggression in cichlids include a remarkably rapid change in coloration, during which a successfully dominant[56] territorial male assumes a more vivid and brighter coloration, while a subordinate or "nonterritorial" male assumes a dull-pale coloration.[57] In addition to color displays, cichlids employ their lateral lines to sense movements of water around their opponents to evaluate the competing male for physical traits/fitness.[58] Male cichlids are very territorial due to the pressure of reproduction, and establish their territory and social status by physically driving out[59] challenging males (novel intruders)[60] through lateral displays (parallel orientation, uncovering gills),[61] biting, or mouth fights (head-on collisions of open mouths, measuring jaw sizes, and biting each other's jaws). The cichlid social dichotomy is composed of a single dominant with multiple subordinates, where the physical aggression of males becomes a contest for resources[59] (mates, territory, food). Female cichlids prefer to mate with a successfully alpha male with vivid coloration, whose territory has food readily available.
Mating
Cichlids mate either monogamously or polygamously.[8] The mating system of a given cichlid species is not consistently associated with its brooding system. For example, although most monogamous cichlids are not mouthbrooders, Chromidotilapia, Gymnogeophagus, Spathodus, and Tanganicodus all include – or consist entirely of – monogamous mouthbrooders. In contrast, numerous open- or cave-spawning cichlids are polygamous; examples include many Apistogramma, Lamprologus, Nannacara, and Pelvicachromis species.[8][62]
Most adult male cichlids, specifically in the cichlid tribe Haplochromini, exhibit a unique pattern of oval-shaped color dots on their anal fins. These phenomena, known as egg spots, aid in the mouthbrooding mechanisms of cichlids. The egg spots consist of carotenoid-based pigment cells, which indicate a high cost to the organism, when considering that fish are not able to synthesize their own carotenoids.[63]
The mimicry of egg spots is used by males for the fertilization process. Mouthbrooding females lay eggs and immediately snatch them up with their mouths. Over millions of years, male cichlids have evolved egg spots to initiate the fertilization process more efficiently.[64] When the females are snatching up the eggs into their mouth, the males gyrate their anal fins, which illuminates the egg spots on his tail. Afterwards, the female, believing these are her eggs, places her mouth to the anal fin (specifically the genital papilla) of the male, which is when he discharges sperm into her mouth and fertilizes the eggs.[63]
The genuine color of egg spots is a yellow, red, or orange inner circle with a colorless ring surrounding the shape. Through phylogenetic analysis, using the mitochondrial ND2 gene, the true egg spots are thought to have evolved in the common ancestor of the Astatoreochromis lineage and the modern Haplochrominis species. This ancestor was most likely riverine in origin, based on the most parsimonious representation of habitat type in the cichlid family.[65] The presence of egg spots in a turbid riverine environment would seem particularly beneficial and necessary for intraspecies communication.[65]
Two pigmentation genes are found to be associated with egg-spot patterning and color arrangement. These are fhl2-a and fhl2-b, which are paralogs.[64] These genes aid in pattern formation and cell-fate determination in early embryonic development. The highest expression of these genes was temporally correlated with egg-spot formation. A short, interspersed, repetitive element was also seen to be associated with egg spots. Specifically, it was evident upstream of the transcriptional start site of fhl2 in only Haplochrominis species with egg spots[64]
Brood care
Pit spawning in cichlids
Pit spawning, also referred to as substrate breeding, is a behavior in cichlid fish in which a fish builds a pit in the sand or ground, where a pair court and consequently spawn.[66] Many different factors go into this behavior of pit spawning, including female choice of the male and pit size, as well as the male defense of the pits once they are dug in the sand.[67]
Cichlids are often divided into two main groups: mouthbrooders and substrate brooders. Different parenting investment levels and behaviors are associated with each type of reproduction.[68] As pit spawning is a reproductive behavior, many different physiological changes occur in the cichlid while this process is occurring that interfere with social interaction.[69] Different kinds of species that pit spawn, and many different morphological changes occur because of this behavioral experience.[66]
Pit spawning is an evolved behavior across the cichlid group. Phylogenetic evidence from cichlids in Lake Tanganyika could be helpful in uncovering the evolution of their reproductive behaviors.[70] Several important behaviors are associated with pit spawning, including parental care, food provisioning,[71] and brood guarding.[72]
Mouth brooding vs. pit spawning
One of the differences studied in African cichlids is reproductive behavior. Some species pit spawn and some are known as mouth brooders. Mouthbrooding is a reproductive technique where the fish scoop up eggs and fry for protection.[68] While this behavior differs from species to species in the details, the general basis of the behavior is the same. Mouthbrooding also affects how they choose their mates and breeding grounds. In a 1995 study, Nelson found that in pit-spawning females choose males for mating based on the size of the pit that they dig, as well as some of the physical characteristics seen in the males.[67] Pit spawning also differs from mouth brooding in the size and postnatal care exhibited. Eggs that have been hatched from pit-spawning cichlids are usually smaller than those of mouthbrooders. Pit-spawners' eggs are usually around 2 mm, while mouthbrooders are typically around 7 mm. While different behaviors take place postnatally between mouthbrooders and pit spawners, some similarities exist. Females in both mouthbrooders and pit-spawning cichlids take care of their young after they are hatched. In some cases, both parents exhibit care, but the female always cares for the eggs and newly hatched fry.[73]
Pit spawning process
Many species of cichlids use pit spawning, but one of the less commonly studied species that exhibits this behavior is the Neotropical Cichlasoma dimerus. This fish is a substrate breeder that displays biparental care after the fry have hatched from their eggs. One study[66] examined reproductive and social behaviors of this species to see how they accomplished their pit spawning, including different physiological factors such as hormone levels, color changes, and plasma cortisol levels. The entire spawning process could take about 90 minutes and 400~800 eggs could be laid. The female deposits about 10 eggs at a time, attaching them to the spawning surface, which may be a pit constructed on the substrate or another surface. The number of eggs laid was correlated to the space available on the substrate. Once the eggs were attached, the male swam over the eggs and fertilized them. The parents would then dig pits in the sand, 10–20 cm wide and 5–10 cm deep, where larvae were transferred after hatching. Larvae began swimming 8 days after fertilization and parenting behaviors and some of the physiological factors measured changed.
Color changes
In the same study, color changes were present before and after the pit spawning occurred. For example, after the larvae were transferred and the pits were beginning to be protected, their fins turned a dark grey color.[66] In another study, of the rainbow cichlid, Herotilapia multispinosa,[69] colr changes occurred throughout the spawning process. Before spawning, the rainbow cichlid was an olive color with grey bands. Once spawning behaviors started, the body and fins of the fish became a more golden color. When the eggs were finished being laid, the pelvic fin all the way back to the caudal fin turned to a darker color and blackened in both the males and the females.[69]
Pit sizes
Females prefer a bigger pit size when choosing where to lay eggs.[67] Differences are seen in the sizes of pits that created, as well as a change in the morphology of the pits.[74] Evolutionary differences between species of fish may cause them to either create pits or castles when spawning. The differences were changes in the way that each species fed, their macrohabitats, and the abilities of their sensory systems.[74]
Evolution
Cichlids are renowned for their recent, rapid evolutionary radiation, both across the entire clade and within different communities across separate habitats.[68][70][74][75][76][77] Within their phylogeny, many parallel instances are seen of lineages evolving to the same trait and multiple cases of reversion to an ancestral trait.
The family Cichlidae arose between 80 and 100 million years ago within the order Perciformes (perch-like fishes).[75] Cichlidae can be split into a few groups based on their geographic location: Madagascar, Indian, African, and Neotropical (or South American). The most famous and diverse group, the African cichlids, can be further split either into Eastern and Western varieties, or into groups depending on which lake the species is from: Lake Malawi, Lake Victoria, or Lake Tanganyika.[75][76] Of these subgroups, the Madagascar and Indian cichlids are the most basal and least diverse.
Of the African cichlids, the West African or Lake Tanganyika cichlids are the most basal.[70][75] Cichlids' common ancestor is believed to have been a spit-spawning species.[76] Both Madagascar and Indian cichlids retain this feature. However, of the African cichlids, all extant substrate brooding species originate solely from Lake Tanganyika.[68][76] The ancestor of the Lake Malawi and Lake Victoria cichlids were mouthbrooders. Similarly, only around 30% of South American cichlids are thought to retain the ancestral substrate-brooding trait. Mouthbrooding is thought to have evolved individually up to 14 times, and a return to substrate brooding as many as three separate times between both African and Neotropical species.[76]
Associated behaviors
Cichlids have a great variety of behaviors associated with substrate brooding, including courtship and parental care alongside the brooding and nest-building behaviors needed for pit spawning. Cichlids' behavior typically revolves around establishing and defending territories when not courting, brooding, or raising young. Encounters between males and males or females and females are agonistic, while an encounter between a male and female leads to courtship.[78] Courtship in male cichlids follows the establishment of some form of territory, sometimes coupled with building a bower to attract mates.[67][74][78] After this, males may attempt to attract female cichlids to their territories by a variety of lekking display strategies or otherwise seek out females of their species.[67] However, cichlids, at the time of spawning, undergo a behavioral change such that they become less receptive to outside interactions.[78] This is often coupled with some physiological change in appearance.[66][69][78]
Brood care
Cichlids can have maternal, paternal, or biparental care. Maternal care is most common among mouthbrooders, but cichlids' common ancestor is thought to exhibit paternal-only care.[76] Other individuals outside of the parents may also play a role in raising young; in the biparental daffodil cichlid (Neolamprologus pulcher), closely related satellite males, those males that surround other males' territories and attempt to mate with female cichlids in the area, help rear the primary males' offspring and their own.[79]
A common form of brood care involves food provisioning. For example, females of lyretail cichlids (Neolamprologus modabu) dig at sandy substrate more to push nutritional detritus and zooplankton into the surrounding water. Adult of N. modabu perform this strategy to collect food for themselves, but dig more when offspring are present, likely to feed their fry.[72][80] This substrate-disruption strategy is rather common and can also be seen in convict cichlids (Cichlasoma nigrofasciatum).[71][80] Other cichlids have an ectothermal mucus that they grow and feed to their young, while still others chew and distribute caught food to offspring. These strategies, however, are less common in pit-spawning cichlids.[80]

Cichlids have highly organized breeding activities.[19] All species show some form of parental care for both eggs and larvae, often nurturing free-swimming young until they are weeks or months old. Communal parental care, where multiple monogamous pairs care for a mixed school of young have also been observed in multiple cichlid species, including Amphilophus citrinellus, Etroplus suratensis, and Tilapia rendalli.[81][82][83] Comparably, the fry of Neolamprologus brichardi, a species that commonly lives in large groups, are protected not only by the adults, but also by older juveniles from previous spawns.[84] Several cichlids, including discus (Symphysodon spp.), some Amphilophus species, Etroplus, and Uaru species, feed their young with a skin secretion from mucous glands.[8][85]
The species Neolamprologus pulcher uses a cooperative breeding system, in which one breeding pair has many helpers that are subordinate to the dominant breeders.
Parental care falls into one of four categories:[85] substrate or open brooders, secretive cave brooders (also known as guarding speleophils[86]), and at least two types of mouthbrooders, ovophile mouthbrooders and larvophile mouthbrooders.[87]
Open brooding
Open- or substrate-brooding cichlids lay their eggs in the open, on rocks, leaves, or logs. Examples of open-brooding cichlids include Pterophyllum and Symphysodon species and Anomalochromis thomasi. Male and female parents usually engage in differing brooding roles. Most commonly, the male patrols the pair's territory and repels intruders, while the female fans water over the eggs, removing the infertile ones, and leading the fry while foraging. Both sexes are able to perform the full range of parenting behaviours.[87]
Cave brooding

Secretive cave-spawning cichlids lay their eggs in caves, crevices, holes, or discarded mollusc shells, frequently attaching the eggs to the roof of the chamber. Examples include Pelvicachromis spp., Archocentrus spp., and Apistogramma spp.[85] Free-swimming fry and parents communicate in captivity and in the wild. Frequently, this communication is based on body movements, such as shaking and pelvic fin flicking. In addition, open- and cave-brooding parents assist in finding food resources for their fry. Multiple neotropical cichlid species perform leaf-turning and fin-digging behaviors.[87]
Ovophile mouthbrooding
Ovophile mouthbrooders incubate their eggs in their mouths as soon as they are laid, and frequently mouthbrood free-swimming fry for several weeks. Examples include many East African Rift lakes (Lake Malawi, Lake Tanganyika, and Lake Victoria) endemics, e.g.: Maylandia, Pseudotropheus, Tropheus, and Astatotilapia burtoni, along with some South American cichlids such as Geophagus steindachneri.
Larvophile mouthbrooding
Larvophile mouthbrooders lay eggs in the open or in a cave and take the hatched larvae into the mouth. Examples include some variants of Geophagus altifrons, and some Aequidens, Gymnogeophagus, and Satanoperca, as well as Oreochromis mossambicus and Oreochromis niloticus.[8][85] Mouthbrooders, whether of eggs or larvae, are predominantly females. Exceptions that also involve the males include eretmodine cichlids (genera Spathodus, Eretmodus, and Tanganicodus), some Sarotherodon species (such as Sarotherodon melanotheron[88]), Chromidotilapia guentheri, and some Aequidens species.[8][87][89] This method appears to have evolved independently in several groups of African cichlids.[19]
Speciation
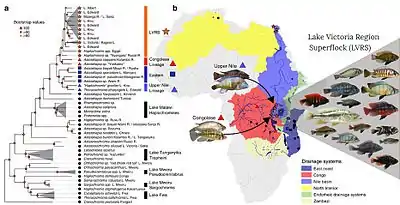
Cichlids provide scientists with a unique perspective of speciation, having become extremely diverse in the recent geological past, those of Lake Victoria actually within the last 10,000 to 15,000 years, a small fraction of the millions taken for Galápagos finch speciation in Darwin's textbook case.[91] Some of the contributing factors to their diversification are believed to be the various forms of prey processing displayed by cichlid pharyngeal jaw apparatus. These different jaw apparatus allow for a broad range of feeding strategies, including algae scraping, snail crushing, planktivory, piscivory, and insectivory.[92] Some cichlids can also show phenotypic plasticity in their pharyngeal jaws, which can also help lead to speciation. In response to different diets or food scarcity, members of the same species can display different jaw morphologies that are better suited to different feeding strategies. As species members begin to concentrate around different food sources and continue their lifecycle, they most likely spawn with like individuals. This can reinforce the jaw morphology and given enough time, create new species.[93] Such a process can happen through allopatric speciation, whereby species diverge according to different selection pressures in different geographical areas, or through sympatric speciation, by which new species evolve from a common ancestor while remaining in the same area. In Lake Apoyo in Nicaragua, Amphilophus zaliosus and its sister species Amphilophus citrinellus display many of the criteria needed for sympatric speciation.[94] In the African rift lake system, cichlid species in numerous distinct lakes evolved from a shared hybrid swarm.[90]
Population status
In 2010, the International Union for Conservation of Nature classified 184 species as vulnerable, 52 as endangered, and 106 as critically endangered.[95] At present, the IUCN only lists Yssichromis sp. nov. argens as extinct in the wild, and six species are listed as entirely extinct, but many more possibly belong in these categories (for example, Haplochromis aelocephalus, H. apogonoides, H. dentex, H. dichrourus, and numerous other members of the genus Haplochromis have not been seen since the 1980s, but are maintained as critically endangered on the small chance that tiny –but currently unknown– populations survive).[95]
Lake Victoria

Because of the introduced Nile perch (Lates niloticus), Nile tilapia (Oreochromis niloticus), and water hyacinth, deforestation that led to water siltation, and overfishing, many Lake Victoria cichlid species have become extinct or been drastically reduced. By around 1980, lake fisheries yielded only 1% cichlids, a drastic decline from 80% in earlier years.[97]
By far the largest Lake Victoria group is the haplochromine cichlids, with more than 500 species, but at least 200 of these (about 40%) have become extinct,[98][99][100] and many others are seriously threatened.[101] Initially it was feared that the percentage of extinct species was even higher,[102] but some species have been rediscovered after the Nile perch started to decline in the 1990s.[99][103] Some species have survived in nearby small satellite lakes,[103] or in refugia among rocks or papyrus sedges (protecting them from the Nile perch),[104] or have adapted to the human-induced changes in the lake itself.[99][100] The species were often specialists and these were not affected to the same extent. For example, the piscivorous haplochromines were particularly hard hit with a high number of extinctions,[105] while the zooplanktivorous haplochromines reached densities in 2001 that were similar to before the drastic decline, although consisting of fewer species and with some changes in their ecology.[99]
Food and game fish
Although cichlids are mostly small- to medium-sized, many are notable as food and game fishes. With few thick rib bones and tasty flesh, artisan fishing is not uncommon in Central America and South America, as well as areas surrounding the African rift lakes.[97]
Tilapia
The most important food cichlids, however, are the tilapiines of North Africa. Fast growing, tolerant of stocking density, and adaptable, tilapiine species have been introduced and farmed extensively in many parts of Asia and are increasingly common aquaculture targets elsewhere.
Farmed tilapia production is about 1,500,000 tonnes (1,700,000 short tons) annually, with an estimated value of US$1.8 billion,[106] about equal to that of salmon and trout.
Unlike those carnivorous fish, tilapia can feed on algae or any plant-based food. This reduces the cost of tilapia farming, reduces fishing pressure on prey species, avoids concentrating toxins that accumulate at higher levels of the food chain, and makes tilapia the preferred "aquatic chickens" of the trade.[97]
Game fish
Many large cichlids are popular game fish. The peacock bass (Cichla species) of South America is one of the most popular sportfish. It was introduced in many waters around the world. In Florida, this fish generates millions of hours of fishing and sportfishing revenue of more than US$8 million a year.[107] Other cichlids preferred by anglers include the oscar, Mayan cichlid (Cichlasoma urophthalmus), and jaguar guapote (Parachromis managuensis).[107]
Aquarium fish

Since 1945, cichlids have become increasingly popular as aquarium fish.[8][85][87][108][109][110][111]
The most common species in hobbyist aquaria is Pterophyllum scalare from the Amazon River basin in tropical South America, known in the trade as the "angelfish". Other popular or readily available species include the oscar (Astronotus ocellatus), convict cichlid (Archocentrus nigrofasciatus) and discus fish (Symphysodon).[8]
Hybrids and selective breeding

Some cichlids readily hybridize with related species, both in the wild and under artificial conditions.[112] Other groups of fishes, such as European cyprinids, also hybridize.[113] Unusually, cichlid hybrids have been put to extensive commercial use, in particular for aquaculture and aquaria.[9][114] The hybrid red strain of tilapia, for example, is often preferred in aquaculture for its rapid growth. Tilapia hybridization can produce all-male populations to control stock density or prevent reproduction in ponds.[9]
Aquarium hybrids
The most common aquarium hybrid is perhaps the blood parrot cichlid, which is a cross of several species, especially from species in the genus Amphilophus. (There are many hypotheses, but the most likely is: Amphilophus labiatus × Vieja synspillus With a triangular-shaped mouth, an abnormal spine, and an occasionally missing caudal fin (known as the "love heart" parrot cichlid), the fish is controversial among aquarists. Some have called blood parrot cichlids "the Frankenstein monster of the fish world".[115] Another notable hybrid, the flowerhorn cichlid, was very popular in some parts of Asia from 2001 until late 2003, and is believed to bring good luck to its owner.[116] The popularity of the flowerhorn cichlid declined in 2004.[117] Owners released many specimens into the rivers and canals of Malaysia and Singapore, where they threaten endemic communities.[118]

Numerous cichlid species have been selectively bred to develop ornamental aquarium strains. The most intensive programs have involved angelfish and discus, and many mutations that affect both coloration and fins are known.[8][119][120] Other cichlids have been bred for albino, leucistic, and xanthistic pigment mutations, including oscars, convict cichlid and Pelvicachromis pulcher.[8][85] Both dominant and recessive pigment mutations have been observed.[121] In convict cichlids, for example, a leucistic coloration is recessively inherited,[122] while in Oreochromis niloticus niloticus, red coloration is caused by a dominant inherited mutation.[123]
This selective breeding may have unintended consequences. For example, hybrid strains of Mikrogeophagus ramirezi have health and fertility problems.[124] Similarly, intentional inbreeding can cause physical abnormalities, such as the notched phenotype in angelfish.[125]
Genera
The genus list is as per FishBase. Studies are continuing, however, on the members of this family, particularly the haplochromine cichlids of the African rift lakes.[16]
|
|
|
Gallery
 The oscar (Astronotus ocellatus) is one of the most popular cichlids in the fishkeeping hobby.
The oscar (Astronotus ocellatus) is one of the most popular cichlids in the fishkeeping hobby.
 The Nile tilapia (Oreochromis niloticus) is farmed extensively as food fish in many parts of the world.
The Nile tilapia (Oreochromis niloticus) is farmed extensively as food fish in many parts of the world. The angelfish (Pterophyllum scalare) has long been commercially bred for the aquarium trade.
The angelfish (Pterophyllum scalare) has long been commercially bred for the aquarium trade. Sexual dimorphism is common in cichlids. Shown here are a male (front, with egg spots) and a female (rear) Maylandia lombardoi.
Sexual dimorphism is common in cichlids. Shown here are a male (front, with egg spots) and a female (rear) Maylandia lombardoi. A pair of blue rams (Mikrogeophagus ramirezi), male in front, female behind. Many cichlids form strong pair bonds while breeding.
A pair of blue rams (Mikrogeophagus ramirezi), male in front, female behind. Many cichlids form strong pair bonds while breeding. A discus (Symphysodon spp.) is guarding its eggs. Advanced broodcare is one of the defining characteristics of cichlids.
A discus (Symphysodon spp.) is guarding its eggs. Advanced broodcare is one of the defining characteristics of cichlids. Lake Malawi, Eastern Africa, is home to numerous cichild species including this Livingston's cichlid (Nimbochromis livingstonii).
Lake Malawi, Eastern Africa, is home to numerous cichild species including this Livingston's cichlid (Nimbochromis livingstonii). Also from Lake Malawi
Also from Lake Malawi Also from Lake Malawi
Also from Lake Malawi
.jpg.webp) The Texas cichlid (Herichthys cyanoguttatus) is the only cichlid native to the United States.
The Texas cichlid (Herichthys cyanoguttatus) is the only cichlid native to the United States..jpg.webp) Pelvicachromis pulcher is a West African riverine cichlid, and part of the aquarists dwarf cichlid group.
Pelvicachromis pulcher is a West African riverine cichlid, and part of the aquarists dwarf cichlid group. The flowerhorn cichlid is a man-made hybrid that has recently gained popularity among aquarists, particularly in Asia.
The flowerhorn cichlid is a man-made hybrid that has recently gained popularity among aquarists, particularly in Asia.
 The red terror cichlid is a highly aggressive species from the rivers of Northeast South America.
The red terror cichlid is a highly aggressive species from the rivers of Northeast South America. A juvenile female Maylandia lombardoi with faint stripes
A juvenile female Maylandia lombardoi with faint stripes A juvenile Aequidens diadema
A juvenile Aequidens diadema
Footnotes
- Cichlid is frequently mispronounced in the pet trade as if spelled "chicklid" /ˈtʃɪklɪd/, presumably from confusion with names like Chiclets, and with Italian words like cioppino and ciao that start with ci- and the sound /tʃ/.
References
- Stiassny, M.L.J.; Jensen, J.S. (1987). "Labroid intrarelationships revisited: morphological complexity, key innovations, and the study of comparative diversity". Bulletin of the Museum of Comparative Zoology. 151: 269–319.
- Wainwright, Peter C.; et al. (2012). "The evolution of pharyngognathy: A phylogenetic and functional appraisal of the pharyngeal jaw key innovation in labroid fishes and beyond". Systematic Biology. 61 (6): 1001–1027. doi:10.1093/sysbio/sys060. PMID 22744773.
- Murray, A.M. (2001). "Eocene cichlid fishes from Tanzania, east Africa". Journal of Vertebrate Paleontology. 20 (4): 651–664. doi:10.1671/0272-4634(2000)020[0651:ECFFTE]2.0.CO;2. JSTOR 4524146. S2CID 86093448.
- Murray, A.M. (2001). "The oldest fossil cichlids (Teleostei: Perciformes): Indication of a 45 million year-old species flock". Proceedings: Biological Sciences. 268 (1468): 679–684. doi:10.1098/rspb.2000.1570. JSTOR 3067613. PMC 1088656. PMID 11321055.
- Nelson, J.S.; Grande, T.C.; Wilson, M.V.H. (2016). Fishes of the World (5th ed.). Wiley. p. 752. ISBN 978-1-118-34233-6. Archived from the original on 8 April 2019. Retrieved 5 November 2018.
- Froese, Rainer; Pauly, Daniel, eds. (February 2012). "List of nominal species of Cichlidae". FishBase.org.
{{cite web}}: CS1 maint: date and year (link) - Stiassny, M.; Teugels, G.G.; Hopkins, C.D. (2007). The Fresh and Brackish Water Fishes of Lower Guinea, West-Central Africa. Vol. 2. Musée Royal de l'Afrique Centrale. p. 269. ISBN 978-90-74752-21-3.
- Loiselle, P.V. (1994). The Cichlid Aquarium. Tetra Press. ISBN 978-1-56465-146-4.
- Kosswig, Curt (June 1963). "Ways of speciation in fishes". Copeia. 1963 (2): 238–244. doi:10.2307/1441338. JSTOR 1441338.
- Reid, G.M. (December 1990). "Captive breeding for the conservation of cichlid fishes". Journal of Fish Biology. 37: 157–166. doi:10.1111/j.1095-8649.1990.tb05031.x.
- Salzburger, W.; Mack, T.; Verheyen, E.; Meyer, A. (2005). "Out of Tanganyika: Genesis, explosive speciation, key-innovations and phylogeography of the haplochromine cichlid fishes". BMC Evolutionary Biology. 5 (17): 17. doi:10.1186/1471-2148-5-17. PMC 554777. PMID 15723698.
- Snoeks, J. (2004). The Cichlid Diversity of Lake Malawi / Nyasa / Niassa: Identification, distribution, and taxonomy. Cichlid Press. ISBN 978-0-9668255-8-9.
- Kornfield, Irv; Smith, Peter (November 2000). "African Cichlid Fishes: Model Systems for Evolutionary Biology". Annual Review of Ecology and Systematics. 31: 163–196. doi:10.1146/annurev.ecolsys.31.1.163.
- "Fact sheet for Oreochromis mossambicus (Peters, 1852)". Gulf States Marine Fisheries Commission. Archived from the original on 18 August 2007. Retrieved 20 October 2006.
- Helfman G.; Collette B.; Facey D. (1997). The Diversity of Fishes. Blackwell Publishing, Inc. pp. 256–257. ISBN 978-0-86542-256-8.
- Froese, Rainer, and Daniel Pauly, eds. (2006). "Cichlidae" in FishBase. April 2006 version.
- Kullander, S.O. (1998). "A phylogeny and classification of the South American Cichlidae (Teleostei: Perciformes)". In L.R. Malabarba; R.E. Reis; R.P. Vari; Z.M. Lucena; C.A.S. Lucena (eds.). Phylogeny and classification of neotropical fishes. Porto Alegre: EDIPUCRS. pp. 461–498. ISBN 978-85-7430-035-1.
- Sparks, J.S.; Smith, W.L. (2004). "Phylogeny and biogeography of cichlid fishes (Teleostei: Perciformes: Cichlidae)". Cladistics. 20 (6): 501–517. CiteSeerX 10.1.1.595.2118. doi:10.1111/j.1096-0031.2004.00038.x. PMID 34892958. S2CID 36086310.
- Nelson, Joseph, S. (2006). Fishes of the World. John Wiley & Sons, Inc. ISBN 978-0-471-25031-9.
{{cite book}}: CS1 maint: multiple names: authors list (link) - "Phylogeny of major groups of cichlids". Archived from the original on 18 January 2012. Retrieved 10 June 2007.
- Streelman, J.T.; Zardoya, R.; Meyer, A.; Karl, S. A. (1 July 1998). "Multilocus phylogeny of cichlid fishes (Pisces: Perciformes): evolutionary comparison of microsatellite and single-copy nuclear loci". Molecular Biology and Evolution. 15 (7): 798–808. doi:10.1093/oxfordjournals.molbev.a025985. PMID 10766579.
- Salzburger, Walter; Meyer, Axel (June 2004). "The species flocks of East African cichlid fishes: Recent advances in molecular phylogenetics and population genetics". Naturwissenschaften. 91 (6): 277–90. Bibcode:2004NW.....91..277S. doi:10.1007/s00114-004-0528-6. PMID 15241604. S2CID 5816449.
- Koehn, John D.; MacKenzie, Rachel F. (August 2004). "Priority management actions for alien freshwater fish species in Australia". New Zealand Journal of Marine and Freshwater Research. 38 (3): 457–472. doi:10.1080/00288330.2004.9517253. S2CID 83569110.
- Boruchowitz, D. E. (2006). Guide to Cichlids. T.F.H. Publications. ISBN 978-0-7938-0584-6.
- ABC Far North Queensland. "Tilapia :: Far North Queensland". Australian Broadcasting Corporation. Archived from the original on 17 October 2007. Retrieved 19 April 2007.
- Froese, R.; D. Pauly (eds.). "Archocentrus nigrofasciatus, Convict cichlid". FishBase. Archived from the original on 1 December 2008. Retrieved 29 March 2007.
- Yamamoto, M.N.; Tagawa, A.W. (2000). Hawai'i's native and exotic freshwater animals. Honolulu, Hawaii: Mutual Publishing. p. 200.
- Page, L.M.; Burr, B.M. (1991). A field guide to freshwater fishes of North America north of Mexico. Boston: Houghton Mifflin Company. pp. 432. ISBN 978-0-395-35307-3.
- University of Southern Mississippi/College of Marine Sciences/Gulf Coast Research Laboratory (3 August 2005). "Fact Sheet for Tilapia zilli (Gervais, 1848)". Gulf States Marine Fisheries Commission. Archived from the original on 18 August 2007. Retrieved 10 February 2007.
- Fuller, Pam L.; Nico, Leo G. (11 October 2002). "Nonindigenous Fishes of Florida – With a Focus on South Florida". U.S. Department of the Interior, U.S. Geological Survey, Center for Coastal Geology. Archived from the original on 15 January 2003.
- Loiselle, Paul (1994). The Cichlid Aquarium, p. 304. Tetra Press, Germany. ISBN 978-1564651464.
- Froese, Rainer; Pauly, Daniel (eds.) (2006). "Alticorpus macrocleithrum" in FishBase. April 2006 version.
- Froese, Rainer; Pauly, Daniel (eds.) (2006). "Pallidochromis tokolosh" in FishBase. April 2006 version.
- Norlander, Britt (20 April 2009). Rough waters: one of the world's most turbulent rivers is home to a wide array of fish species. Now, large dams are threatening their future. Science World
- Frank Schäfer (2005). Brackish-Water Fishes. Aqualog. ISBN 978-3-936027-82-2.
- Stiassny, Melanie L. J.; Marchi, Giuseppe De; Lamboj, Anton (29 November 2010). "A new species of Danakilia (Teleostei, Cichlidae) from Lake Abaeded in the Danakil Depression of Eritrea (East Africa)" (PDF). Zootaxa. 2690 (1): 43–52. doi:10.11646/zootaxa.2690.1.4. S2CID 87705274. Archived from the original (PDF) on 12 February 2020.
- Chakrabarty, Prosanta (June 2004). "Cichlid biogeography: comment and review". Fish and Fisheries. 5 (2): 97–119. doi:10.1111/j.1467-2979.2004.00148.x. hdl:2027.42/72313.
- Stiassny, Melanie L.J; Sparks, John S (2006). "Phylogeny and Taxonomic Revision of the Endemic Malagasy Genus Ptychochromis (Teleostei: Cichlidae), with the Description of Five New Species and a Diagnosis for Katria, New Genus". American Museum Novitates (3535): 1. doi:10.1206/0003-0082(2006)3535[1:PATROT]2.0.CO;2. S2CID 322493.
- Sparks, John S. (2008). "Phylogeny of the Cichlid Subfamily Etroplinae and Taxonomic Revision of the Malagasy Cichlid Genus Paretroplus (Teleostei: Cichlidae)". Bulletin of the American Museum of Natural History. 314 (1): 1. doi:10.1206/314.1. S2CID 84071748.
- Froese, Rainer; Pauly, Daniel (eds.) (2006). "Etroplus maculatus" in FishBase. April 2006 version.
- Froese, Rainer; Pauly, Daniel (eds.) (2006). "Etroplus suratensis" in FishBase. April 2006 version.
- Kullander, S.O. "Family Cichlidae-Cichlids". FishBase. Retrieved 4 December 2019.
- Jonna, R. Jamil. "Cichlidae". Animal Diversity Web. University of Michigan Museum of Zoology. Retrieved 3 December 2019.
- De Iuliis, Gerardo; Pulerà, Dino (2011). "The Shark". The Dissection of Vertebrates. pp. 27–77. doi:10.1016/B978-0-12-375060-0.00003-6. ISBN 978-0-12-375060-0.
- Ribbink, A.J.; Lewis, D.S.C. (1981). "Melanochromis Crabro Sp. Nov.: a Cichlid Fish From Lake Malawi Which Feeds On Ectoparasites and Catfish Eggs". Netherlands Journal of Zoology. 32 (1): 72–87. doi:10.1163/002829682X00058.
- Oliver, M.K. (18 November 1999). "Rhamphochromis esox". malawicichlids.com: The Cichlid Fishes of Lake Malawi. Retrieved 19 April 2007.
- Ribbink, A.J.; Ribbink, A.C. (1997). "Paedophagia among cichlid fishes of Lake Victoria and Lake Malawi/Nyasa". South African Journal of Science. 93: 509–512.
- McKaye, K.R.; Kocher, T. (1983). "Head ramming behaviour by three paedophagous cichlids in Lake Malawi, Africa". Animal Behaviour. 31: 206–210. doi:10.1016/S0003-3472(83)80190-0. S2CID 53156766.
- Wilhelm, W. (1980). "The disputed feeding behavior of a paedophagous haplochromine cichlid (Pisces) observed and discussed". Behaviour. 74 (3): 310–322. doi:10.1163/156853980X00528.
- Konings, A. (2007). "Paedophagy in Malawi cichlids". Cichlid News. 16: 28–32.
- Trewavas, E. (1947). "An example of "mimicry" in fishes". Nature. 160 (4056): 120. Bibcode:1947Natur.160..120T. doi:10.1038/160120a0. PMID 20256157. S2CID 4140785.
- Eccles, D.H.; D.S.C. Lewis (1976). "A taxonomic study of the genus Docimodus Boulenger (Pisces, Cichlidae) a group of fishes with unusual feeding habits from Lake Malawi". Zoological Journal of the Linnean Society. 58 (2): 165–172. doi:10.1111/j.1096-3642.1976.tb00826.x.
- Nshombo, M. (1991). "Occasional egg-eating by the scale-eater Plecodus straeleni (Cichlidae) of Lake Tanganyika". Environmental Biology of Fishes. 31 (2): 207–212. doi:10.1007/BF00001022. S2CID 38219021.
- Tobler, M. (2005). "Feigning death in the Central American cichlid Parachromis friedrichsthalii". Journal of Fish Biology. 66 (3): 877–881. doi:10.1111/j.0022-1112.2005.00648.x.
- McKaye, K.R. (1981). "Field observation on death feigning: a unique hunting behavior by the predatory cichlid, Haplochromis livingstoni, of Lake Malawi". Environmental Biology of Fishes. 6 (3–4): 361–365. doi:10.1007/BF00005766. S2CID 24244576.
- Shwartz, Mark (25 January 2007). "Study: Cichlids can determine their social rank by observation". Stanford University. Retrieved 11 December 2018.
- Chen, Chun-Chun; Fernald, Russell D. (25 May 2011). "Visual Information Alone Changes Behavior and Physiology during Social Interactions in a Cichlid Fish (Astatotilapia burtoni)". PLOS ONE. 6 (5): e20313. Bibcode:2011PLoSO...620313C. doi:10.1371/journal.pone.0020313. PMC 3102105. PMID 21633515.
- Knight, Kathryn (1 October 2015). "Fighting cichlids size up opposition with lateral line". Journal of Experimental Biology. 218 (20): 3161. doi:10.1242/jeb.132563.
- Fernald, Russell D. (1 January 2017). "Cognitive skills and the evolution of social systems". Journal of Experimental Biology. 220 (1): 103–113. doi:10.1242/jeb.142430. PMC 5278620. PMID 28057833.
- Fernald, Russell D.; Kent, Kai R.; Hilliard, Austin T.; Becker, Lisa; Alcazar, Rosa M. (15 August 2016). "Two types of dominant male cichlid fish: behavioral and hormonal characteristics". Biology Open. 5 (8): 1061–1071. doi:10.1242/bio.017640. PMC 5004607. PMID 27432479.
- Arnott, Gareth; Ashton, Charlotte; Elwood, Robert W. (23 October 2011). "Lateralization of lateral displays in convict cichlids". Biology Letters. 7 (5): 683–685. doi:10.1098/rsbl.2011.0328. PMC 3169077. PMID 21508024.
- Martin, E.; M. Taborsky (1997). "Alternative male mating acttics in a cichlid, Pelvicachromis pulcher: a comparison of reproductive effort and success". Behavioral Ecology and Sociobiology. 41 (5): 311–319. doi:10.1007/s002650050391. S2CID 27852520.
- Egger, Bernd; Klaefiger, Yuri; Theis, Anya; Salzburger, Walter (18 October 2011). "A sensory bias has triggered the evolution of egg-spots in cichlid fishes". PLOS ONE. 6 (10): e25601. Bibcode:2011PLoSO...625601E. doi:10.1371/journal.pone.0025601. PMC 3196499. PMID 22028784.
- Santos, M. Emília; Braasch, Ingo; Boileau, Nicolas; Meyer, Britta S.; Sauteur, Loïc; Böhne, Astrid; et al. (9 October 2014). "The evolution of cichlid fish egg-spots is linked with a cis-regulatory change". Nature Communications. 5: 5149. Bibcode:2014NatCo...5.5149S. doi:10.1038/ncomms6149. PMC 4208096. PMID 25296686.
- Salzburger, Walter; Mack, Tanja; Verheyen, Erik; Meyer, Axel (1 January 2005). "Out of Tanganyika: Genesis, explosive speciation, key-innovations and phylogeography of the haplochromine cichlid fishes". BMC Evolutionary Biology. 5: 17. doi:10.1186/1471-2148-5-17. PMC 554777. PMID 15723698.
- Alonso, Felipe; Cánepa, Maximiliano; Moreira, Renata Guimarães; Pandolfi, Matías (22 July 2011). "Social and reproductive physiology and behavior of the Neotropical cichlid fish Cichlasoma dimerus under laboratory conditions". Neotropical Ichthyology. 9 (3): 559–570. doi:10.1590/S1679-62252011005000025.
- Nelson, C. Mindy (1 January 1995). "Male size, spawning pit size and female mate choice in a lekking cichlid fish". Animal Behaviour. 50 (6): 1587–1599. doi:10.1016/0003-3472(95)80013-1. S2CID 54249065.
- Duponchelle, Fabrice; Paradis, Emmanuel; Ribbink, Anthony J.; Turner, George F. (7 October 2008). "Parallel life history evolution in mouthbrooding cichlids from the African Great Lakes". Proceedings of the National Academy of Sciences. 105 (40): 15475–15480. Bibcode:2008PNAS..10515475D. doi:10.1073/pnas.0802343105. PMC 2563094. PMID 18824688.
- Brown, Dean H.; Marshall, Joseph A. (1978). "Reproductive Behaviour of the Rainbow Cichlid, Herotilapia Multispinosa (Pisces, Cichlidae)". Behaviour. 67 (3–4): 299–321. doi:10.1163/156853978X00378.
- Muschick, Moritz; Indermaur, Adrian; Salzburger, Walter (December 2012). "Convergent Evolution within an Adaptive Radiation of Cichlid Fishes". Current Biology. 22 (24): 2362–2368. doi:10.1016/j.cub.2012.10.048. PMID 23159601. S2CID 18363916.
- Wisenden, Brian D.; Lanfranconi-Izawa, Tanya L.; Keenleyside, Miles H.A. (March 1995). "Fin digging and leaf lifting by the convict cichlid, Cichlasoma nigrofasciatum: examples of parental food provisioning". Animal Behaviour. 49 (3): 623–631. doi:10.1016/0003-3472(95)80195-2. S2CID 41529998.
- Ota, Kazutaka; Kohda, Masanori (9 June 2014). "Maternal Food Provisioning in a Substrate-Brooding African Cichlid". PLOS ONE. 9 (6): e99094. Bibcode:2014PLoSO...999094O. doi:10.1371/journal.pone.0099094. PMC 4049616. PMID 24911060.
- Sefc, Kristina M. (21 July 2011). "Mating and parental care in Lake Tanganyika's cichlids". International Journal of Evolutionary Biology. 2011: 470875. doi:10.4061/2011/470875. PMC 3142683. PMID 21822482.
- York, Ryan A.; Patil, Chinar; Hulsey, C. Darrin; Streelman, J. Todd; Fernald, Russell D. (27 February 2015). "Evolution of bower building in Lake Malawi cichlid fish: phylogeny, morphology, and behavior". Frontiers in Ecology and Evolution. 3. doi:10.3389/fevo.2015.00018. S2CID 7458974.
- Zardoya, Rafael; Vollmer, Dana M.; Craddock, Clark; Streelman, Jeffrey T.; Karl, Steve; Meyer, Axel (22 November 1996). "Evolutionary conservation of microsatellite flanking regions and their use in resolving the phylogeny of cichlid fishes (Pisces: Perciformes)". Proceedings of the Royal Society of London. Series B: Biological Sciences. 263 (1376): 1589–1598. Bibcode:1996RSPSB.263.1589Z. doi:10.1098/rspb.1996.0233. PMID 8952095. S2CID 45834758.
- Goodwin, Nicholas B.; Balshine-Earn, Sigal; Reynolds, John D. (7 December 1998). "Evolutionary transitions in parental care in cichlid fish". Proceedings of the Royal Society of London. Series B: Biological Sciences. 265 (1412): 2265–2272. doi:10.1098/rspb.1998.0569. PMC 1689529.
- Hulsey, C Darrin; Hollingsworth, Phillip R; Fordyce, James A (December 2010). "Temporal diversification of Central American cichlids". BMC Evolutionary Biology. 10 (1): 279. doi:10.1186/1471-2148-10-279. PMC 2944184. PMID 20840768.
- Burchard, John E. (26 April 2010). "Family Structure in the Dwarf Cichlid Apistogramma trifasciatum Eigenmann and Kennedy)". Zeitschrift für Tierpsychologie. 22 (2): 150–162. doi:10.1111/j.1439-0310.1965.tb01428.x. PMID 5890861.
- Dierkes, P. (1 September 1999). "Reproductive parasitism of broodcare helpers in a cooperatively breeding fish". Behavioral Ecology. 10 (5): 510–515. doi:10.1093/beheco/10.5.510.
- Zworykin, Dmitry (2001). "Parental brood provisioning in cichlid fishes by means of stirring up of the loose material of bottom substrate: a brief review". Cichlid Research: State of the Art. pp. 269–286. doi:10.13140/2.1.1937.5048.
- McKaye, K.R.; N.M. McKaye (1977). "Communal Care and Kidnapping of Young by Parental Cichlids". Evolution. 31 (3): 674–681. doi:10.2307/2407533. JSTOR 2407533. PMID 28563477.
- Ward, J.A.; Wyman, R.L. (1977). "Ethology and ecology of cichlid fishes of the genus Etroplus in Sri Lanka: Preliminary findings". Environmental Biology of Fishes. 2 (2): 137–145. doi:10.1007/BF00005369. S2CID 8307811.
- Ribbink, A.J.; Marsh, A.C.; Marsh, B.A. (1981). "Nest-building and communal care of young by Tilapia rendalli dumeril (pisces, cichlidae) in Lake Malawi". Environmental Biology of Fishes. 6 (2): 219–222. doi:10.1007/BF00002787. S2CID 9546191.
- Steeves, Greg. Neolamprologus brichardi. Archived 5 May 2008 at the Wayback Machine africancichlids.net. Retrieved 8 April 2008
- Baensch, H.A. (1996). Riehl, Rüdiger (ed.). Aquarium Atlas (5th ed.). Germany: Tetra Press. ISBN 978-3-88244-050-8.
- Balon, E.K. (1975). "Reproductive guilds of fishes: A proposal and definition". Journal of the Fisheries Research Board of Canada. 32 (6): 821–864. doi:10.1139/f75-110.
- Keenleyside, M.H.A. (1991). "Parental care". Cichlid Fishes: Behaviour, ecology and evolution. London, UK: Chapman and Hall. pp. 191–208. ISBN 978-0-412-32200-6.
- Kishida, Mitsuyo; Specker, Jennifer L. (February 2000). "Paternal mouthbrooding in the black-chinned tilapia, Sarotherodon melanotheron (Pisces: Cichlidae): Changes in gonadal steroids and potential for vitellogenin transfer to larvae" (PDF). Hormones and Behavior. 37 (1): 40–48. doi:10.1006/hbeh.1999.1556. PMID 10712857. S2CID 8128737.
- Coleman, R. (January 1999). "Mysterious mouthbrooders". Cichlid News: 32–33.
- Meier, Joana I.; Marques, David A.; Mwaiko, Salome; Wagner, Catherine E.; Excoffier, Laurent; Seehausen, Ole (10 February 2017). "Ancient hybridization fuels rapid cichlid fish adaptive radiations". Nature Communications. 8: 14363. Bibcode:2017NatCo...814363M. doi:10.1038/ncomms14363. PMC 5309898. PMID 28186104.
- Meyer, Axel (April 2015). "The extraordinary evolution of cichlid fishes". Scientific American. Retrieved 29 April 2023.
- Albertson, R.C.; Markert, J.A.; Danley, P.D.; Kocher, T.D. (1999). "Phylogeny of a rapidly evolving clade: The cichlid fishes of Lake Malawi, East Africa". PNAS. 96 (9): 5107–5110. Bibcode:1999PNAS...96.5107A. doi:10.1073/pnas.96.9.5107. PMC 21824. PMID 10220426.
- Muschick, M.; Barluenga, M.; Salzburger, W.; Meyer, A. (2011). "Adaptive phenotypic plasticity in the Midas cichlid fish pharyngeal jaw and its relevance in adaptive radiation". BMC Evolutionary Biology. 11: 116. doi:10.1186/1471-2148-11-116. PMC 3103464. PMID 21529367.
- Barluenga, M.; Meyer, A.; Muschick, M.; Salzburger, W.; Stolting, K.N. (2006). "Sympatric speciation in Nicaraguan crater lake cichlid fish" (PDF). Nature. 439 (7077): 719–23. Bibcode:2006Natur.439..719B. doi:10.1038/nature04325. PMID 16467837. S2CID 3165729.
- IUCN Red List of Threatened Species (Report). 2010.4. IUCN. 2010. Retrieved 26 April 2011. IUCN does not allow linking to search results. Use "Other Search Options" on the IUCN frontpage, then choose "Cichlidae" (Animalia→Chordata→Actinopterygii→Perciformes) under taxonomy, and then the specific threat category (Vulnerable, Endangered, etc) under Assessment.
- Witte, F.; de Zeeuw, M.P.; Brooks, E. (2010). "Haplochromis thereuterion". IUCN Red List of Threatened Species. 2010: e.T185857A8492470. doi:10.2305/IUCN.UK.2010-3.RLTS.T185857A8492470.en.
- Barlow, G. W. (2000). The Cichlid Fishes. Cambridge, MA: Perseus Publishing. ISBN 978-0-7382-0376-8.
- Deweerdt, Sarah (28 February 2004). "Dark secret of the lake". New Scientist.
- Lowe-McConnell, Rosemary (December 2009). "Fisheries and Cichlid Evolution in the African Great Lakes: Progress and Problems". Freshwater Reviews. 2 (2): 131–151. doi:10.1608/frj-2.2.2. S2CID 54011001.
- van Rijssel, Jacco C.; Witte, Frans (March 2013). "Adaptive responses in resurgent Lake Victoria cichlids over the past 30 years". Evolutionary Ecology. 27 (2): 253–267. doi:10.1007/s10682-012-9596-9. S2CID 2291741.
- Fiedler, P.L.; Kareiva, P.M., eds. (1998). Conservation Biology: For the coming decade (2nd ed.). Springer. pp. 209–210. ISBN 978-0412096617.
- Goldschmidt, Tijs; Witte, Frans; Wanink, Jan (September 1993). "Cascading Effects of the Introduced Nile Perch on the Detritivorous/Phytoplanktivorous Species in the Sublittoral Areas of Lake Victoria" (PDF). Conservation Biology. 7 (3): 686–700. doi:10.1046/j.1523-1739.1993.07030686.x. S2CID 39341574. Archived from the original (PDF) on 19 February 2019.
- IUCN Red Lists: Geographic Patterns. Eastern Africa. Retrieved 25 March 2017.
- Chapman, Lauren J.; Chapman, Colin A.; Chandler, Mark (December 1996). "Wetland ecotones as refugia for endangered fishes". Biological Conservation. 78 (3): 263–270. CiteSeerX 10.1.1.689.2625. doi:10.1016/s0006-3207(96)00030-4.
- McGee, M. D.; Borstein, S. R.; Neches, R. Y.; Buescher, H. H.; Seehausen, O.; Wainwright, P. C. (27 November 2015). "A pharyngeal jaw evolutionary innovation facilitated extinction in Lake Victoria cichlids". Science. 350 (6264): 1077–1079. Bibcode:2015Sci...350.1077M. doi:10.1126/science.aab0800. PMID 26612951.
- Silva, Sena S. De; Nations, Food and Agriculture Organization of the United (2004). Tilapias as Alien Aquatics in Asia and the Pacific: A Review. Food & Agriculture Org. ISBN 978-92-5-105227-3.
- Florida Fish and Wildlife Conservation Commission. "Fact Exotic Freshwater Fishes". Retrieved 18 March 2007.
- Sands, D. (1994). A Fishkeepers Guide to Central American Cichlids. Belgium: Tetra Press. pp. 59–60.
- Mills, D. (1993). Aquarium Fish. Harper Collins. ISBN 0-7322-5012-9.
- Konings, A. (1997). Back to Nature Guide to Malawi Cichlids. Germany: Druckhaus Beltz. pp. 13–23.
- Leibel, W.S. (1993). A Fishkeepers Guide to South American Cichlids. Belgium: Tetra Press. pp. 12–14.
- Smith, P.F.; Konings, A.; Kornfield, I. (July 2003). "Hybrid origin of a cichlid population in Lake Malawi: Implications for genetic variation and species diversity". Molecular Ecology. 12 (9): 2497–2504. doi:10.1046/j.1365-294X.2003.01905.x. PMID 12919487. S2CID 2829927.
- Wood, A.B.; Jordan, D. R. (March 1987). "Fertility of roach × bream hybrids, Rutilus rutilus (L.) × Abramis brama (L.), and their identification". Journal of Fish Biology. 30 (3): 249–261. doi:10.1111/j.1095-8649.1987.tb05750.x.
- Clarke, Matt. "Frequently asked questions on parrot cichlids". Practical Fishkeeping. Archived from the original on 26 September 2007. Retrieved 20 October 2006.
- "It's The Frankenstein Monster of the Fish World: The Blood Parrot!". AquaFriend.com. 27 October 2002. Archived from the original on 16 May 2006. Retrieved 5 December 2006.
- Arnold, W. (1 July 2003). "Singapore's 'lucky' pet Luohan can outnumber people in homes". International Herald Tribune.
- "Crayfish the latest fad among pet lovers". New Straits Times. Singapore, Malaysia. 3 September 2004.
- "Flower horn: Joy in homes, a pest in rivers". New Straits Times. Singapore, Malaysia. 14 July 2004.
- Norton, J. (1982). "Angelfish genetics". Freshwater and Marine Aquarium Magazine. Vol. 5. p. 4.
- Koh, Tieh Ling; Khoo, Gideon; Fan, Li Qun; Phang, Violet Pan Eng (30 March 1999). "Genetic diversity among wild forms and cultivated varieties of Discus (Symphysodon spp.) as revealed by random amplified polymorphic DNA (RAPD) fingerprinting". Aquaculture. 173 (1): 485–497. doi:10.1016/S0044-8486(98)00478-5.
- Kornfield, I. (1991). "Genetics". In Keenleyside, M.H.A. (ed.). Cichlid Fishes: Behaviour, ecology and evolution. London, UK: Chapman and Hall. pp. 109–115.
- Itzkovich, J.; Rothbard, S.; Hulata, G. (1981). "Inheritance of pink body colouration in Cichlasoma nigrofasciatum Günther (Pisces, Cichlidae)". Genetica. 55: 15–16. doi:10.1007/bf00133997. S2CID 38828706.
- McAndrew, C.J.; Roubal, F.R.; Roberts, R.J.; Bullock, A.M.; McEwan, I.M. (1988). "The genetics and history of red, blond, and associated color variants in Oreochromis niloticus". Genetica. 76 (2): 127–137. doi:10.1007/bf00058811. S2CID 40666053.
- Linke H, Staeck L (1994) American cichlids I: Dwarf Cichlids. A handbook for their identification, care and breeding. Tetra Press. Germany. ISBN 1-56465-168-1
- Norton, J (1994). "Notched – An Angelfish Deformity". Freshwater and Marine Aquarium Magazine. 17 (3).
- Dunz, A.R.; Schliewen, U.K. (2013). "Molecular phylogeny and revised classification of the haplotilapiine cichlid fishes formerly referred to as "Tilapia"". Molecular Phylogenetics and Evolution. 68 (1): 64–80. doi:10.1016/j.ympev.2013.03.015. PMID 23542002.
- De la Maza-Benignos, M., Ornelas-García, C. P., Lozano-Vilano, M.d.L., García-Ramírez, M.E. & Doadrio, I. (2015). "Phylogeographic analysis of genus Herichthys (Perciformes: Cichlidae), with descriptions of Nosferatu new genus and H. tepehua n. sp". Hydrobiologia. 748 (1): 201–231. doi:10.1007/s10750-014-1891-8. hdl:10261/126238. S2CID 16769534.
{{cite journal}}: CS1 maint: multiple names: authors list (link)
Further reading
- Barlow, G.W. (2000). The Cichlid Fishes. Cambridge, MA: Perseus Publishing.
- "Cichlidae". Integrated Taxonomic Information System.: National Museum of Natural History, Washington, D.C., 2004-05-11).
- Sany, R.H. (2012). Taxonomy of Cichlids and Angels (web publication).
External links
- Cichlid at Curlie
- Oliver, Michael, Dr., ed. (15 October 2021) [7 May 1997]. "The cichlid fishes of Lake Malawi, Africa". MalawiCichlids.com (main page). Retrieved 1 May 2023.
{{cite web}}: CS1 maint: multiple names: editors list (link) - van der Meer, H.J., Dr. (2008–2013). Vision in cichlids: Ecomorphology of vision in haplochromine cichlids of Lake Victoria (Report).
{{cite report}}: CS1 maint: multiple names: authors list (link) - . Encyclopædia Britannica. Vol. 6 (11th ed.). 1911. p. 360.