Massospondylus
Massospondylus (/ˌmæsoʊˈspɒndɪləs/ mas-oh-SPON-di-ləs; from Greek, μάσσων (massōn, "longer") and σπόνδυλος (spondylos, "vertebra")) is a genus of sauropodomorph dinosaur from the Early Jurassic (Hettangian to Pliensbachian ages, ca. 200–183 million years ago). It was described by Sir Richard Owen in 1854 from remains discovered in South Africa, and is thus one of the first dinosaurs to have been named. Fossils have since been found at other locations in South Africa, Lesotho, and Zimbabwe. Material from Arizona's Kayenta Formation, India, and Argentina has been assigned to the genus at various times, but the Arizonan and Argentinian material are now assigned to other genera.
| Massospondylus Temporal range: Early Jurassic, | |
|---|---|
 | |
| Reconstructed skeleton on display in the Royal Ontario Museum | |
| Scientific classification | |
| Domain: | Eukaryota |
| Kingdom: | Animalia |
| Phylum: | Chordata |
| Clade: | Dinosauria |
| Clade: | Saurischia |
| Clade: | †Sauropodomorpha |
| Clade: | †Bagualosauria |
| Clade: | †Plateosauria |
| Clade: | †Massopoda |
| Family: | †Massospondylidae |
| Genus: | †Massospondylus Owen, 1854 |
| Species | |
| Synonyms | |
|
List
| |
The type species is M. carinatus; seven other species have been named during the past 150 years, but only M. kaalae is still considered valid. Early sauropodomorph systematics have undergone numerous revisions during the last several years, and many scientists disagree where exactly Massospondylus lies on the dinosaur evolutionary tree. The family name Massospondylidae was once coined for the genus, but because knowledge of an early sauropod relationship is in a state of flux, it is unclear which other dinosaurs—if any—belong in a natural grouping of massospondylids; several 2007 papers support the family's validity.
Although Massospondylus was long depicted as quadrupedal, a 2007 study found it to be bipedal. It was probably a plant eater (herbivore), although it is speculated that the early sauropodomorphs may have been omnivorous. The genus was 4–6 metres (13–20 ft) long, and had a long neck and tail and a small head and slender body. On each of its forefeet, it bore a sharp thumb claw that was used in defense or feeding. Recent studies indicate that Massospondylus grew steadily throughout its lifespan, possessed air sacs similar to those of birds, and may have cared for its young.
History of discovery
.jpg.webp)
The first fossils of Massospondylus were described by paleontologist Sir Richard Owen in 1854.[1] Originally, Owen did not recognize the finds as those of a dinosaur; instead he attributed them to "large, extinct, carnivorous reptiles"[2] that were related to modern lizards, chameleons, and iguanas.[1] The material, a collection of 56 bones, was found in 1853 by the government surveyor Joseph Millard Orpen in the Upper Elliot Formation at Harrismith, South Africa and was donated to the Hunterian Museum at the Royal College of Surgeons in London.[1][3][4] Among the remains were vertebrae from the neck, back, and tail; a shoulder blade; a humerus; a partial pelvis; a femur; a tibia; and bones of the hands and feet.[5] All these bones were found disarticulated, making it difficult to determine if all material belongs to a single species or not. However, Owen was able to distinguish three different types of caudal vertebrae, which he attributed to three different genera: Pachyspondylus, Leptospondylus and Massospondylus. Massospondylus was separated from the other two genera on the basis of its much longer caudal vertebrae, which also led to the scientific name that has been derived from the Greek terms masson/μάσσων 'longer' and spondylos/σπόνδυλος 'vertebra', explained by Owen as "because the vertebrae are proportionally longer than those of the extinct Crocodile called Macrospondylus".[1][3] However, later it was shown that the putative caudal vertebrae of Massospondylus were actually cervical vertebrae and that all the material probably belongs only to a single species. On May 10, 1941, the Hunterian Museum was demolished by a German bomb, destroying all the fossils; only casts remain. Because the plaster casts of the lost type specimen fossils were not adequate to accurately diagnose a genus and species under modern taxonomic practices and for research purposes, Yates and Barrett (2010) designated BP/1/4934, a skull and a largely complete postcranial skeleton in the Bernard Price Institute for Palaeontological Research, as the neotype specimen.[3]
Massospondylus remains have been found in the Upper Elliot Formation, the Clarens Formation, and the Bushveld Sandstone of South Africa and Lesotho, as well as the Forest Sandstone of Zimbabwe. These remains consist of at least 80 partial skeletons and four skulls, representing both juveniles and adults.[6] The report of Massospondylus from Arizona's Kayenta Formation is based on a skull described in 1985. The skull of the Kayenta specimen from Arizona is 25% larger than the largest skull from any African specimen.[7] The Kayenta specimen possesses four teeth in the premaxilla and sixteen in the maxilla. Uniquely among dinosaurs, it also had tiny, one-millimetre-(0.04 in-) long palatal teeth.[8] A 2004 restudy of African Massospondylus skulls, however, indicated that the Kayenta specimen did not pertain to Massospondylus.[9] This Kayenta skull and associated postcranial elements, identified collectively as MCZ 8893, was referred to the new genus Sarahsaurus in 2010.[10]
Massospondylus had also been reported from Argentina,[6] but the material has been reassessed as a closely related but distinct genus. The fossils included several partial skeletons and at least one skull, found in the Lower Jurassic Canon del Colorado Formation of San Juan, Argentina. lt was assigned to Adeopapposaurus in 2009.[11] A specimen from South Africa previously assigned to Massospondylus, BP/1/4779, became the holotype of the new genus and species Ngwevu intloko in 2019.[12]
Species
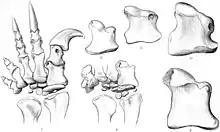
Many species have been named, although most are no longer considered valid. M. carinatus, named by Richard Owen, is the type species.[13] Other named species include: M. browni Seeley, 1895,[14] M. harriesi Broom 1911,[4] M. hislopi Lydekker, 1890,[15] M. huenei Cooper, 1981,[2] M. kaalae Barrett 2009,[16] M. rawesi Lydekker, 1890,[15] and M. schwarzi Haughton, 1924.[17]
M. browni, M. harriesi, and M. schwarzi were all found in the Upper Elliot Formation of Cape Province, South Africa. All three are based on fragmentary material, and were regarded as indeterminate in the most recent review.[6] M. browni is based on two cervical, two back, and three caudal vertebrae and miscellaneous hind limb elements.[14] M. harriesi is known from a well-preserved forelimb and parts of a hindlimb.[4] M. schwarzi is known from an incomplete hind limb and sacrum.[17] M. hislopi and M. rawesi were named from fossils found in India.[15] M. hislopi is based on vertebrae from the Upper Triassic Maleri Formation of Andhra Pradesh, whereas M. rawesi is based on a tooth from the Upper Cretaceous Takli Formation of Maharashtra.[18] M. hislopi was tentatively retained as an indeterminate sauropodomorph in the latest review,[6] but M. rawesi may be a theropod[19] or nondinosaur.[5] M. huenei is a combination derived by Cooper for Lufengosaurus huenei, as he considered Lufengosaurus and Massospondylus to be synonyms.[2] This synonymy is no longer accepted.[6]
M. kaalae was described in 2009 on the basis of a partial skull from the Upper Elliot Formation in Eastern Cape of South Africa. The species is known from the same time and region as some specimens of M. carinatus. It differs from the type species in the morphology of the braincase, as well as in several other characters of the skull such as the proportions of the premaxilla.[16]
Dubious names
Several genera have been previously synonymized with Massospondylus. These include the above-mentioned Leptospondylus and Pachyspondylus, as well as Aristosaurus, Dromicosaurus, Gryponyx taylori and Hortalotarsus, which are dubious names of little scientific value.[2] Together with Massospondylus carinatus, Owen named Leptospondylus capensis and Pachyspondylus orpenii for vertebrae from the same unit and location. Aristosaurus erectus was named by E.C.N. van Hoepen in 1920 based on a nearly complete skeleton, and Hoepen also named Dromicosaurus gracilis, which consisted of a partial skeleton. Gryponyx taylori was named by Sidney H. Haughton in 1924. It consists of hip bones. All of the above fossils come from the Hettangian or Sinemurian faunal stages of South Africa, where Massospondylus has been found.[20] Under the rules of zoological nomenclature, these names are junior synonyms. Massospondylus was described in a scientific paper; therefore the name Massospondylus takes priority.
Ignavusaurus, known from a young specimen, was considered by Yates et al. (2011) to be a probable synonym of Massospondylus.[21] Cladistic analyses by Chapelle & Choiniere (2018) and Chapelle et al. (2019) agreed with Yates et al. (2011) in demonstrating the massospondylid nature of Ignavusaurus, but nevertheless recovered it as a distinct taxon of massospondylid.[22][23]
Description

Massospondylus was a mid-size sauropodomorph, around 4 metres (13 ft) in length,[24][25][26] and weighed approximately 1000 kilograms (2200 lb),[26] although a few sources have estimated its length at up to 6 metres (20 ft).[27][28] It was a typical early sauropodomorph, with a slender body, a long neck and a proportionally very small head.[6]

The vertebral column was composed of nine cervical (neck) vertebrae, 13 dorsal (back) vertebrae, three sacral (hip) vertebrae, and at least 40 caudal (tail) vertebrae. The pubis faced forward, as with most saurischians. It had a slighter build than that of Plateosaurus, an otherwise similar dinosaur.[6] The neck was proportionally longer than in most other plateosaurids, with the foremost cervicals being four times the length of their width.[29] The forelimbs were only half the length of the hindlimbs[30] but quite powerful, as indicated by the broad upper end of the humerus that provided attachment areas for a large arm musculature.[29] Like Plateosaurus, it had five digits on each hand and foot. The hand was short and wide, with a large sickle shaped thumb claw used for feeding or defense against predators. The thumb was the longest finger in the hand, while the fourth and fifth digits were tiny, giving the forepaws a lopsided look.[29][2]
Cranial anatomy
The small head of Massospondylus was approximately half the length of the femur. Numerous openings, or fenestrae, in the skull reduced its weight and provided space for muscle attachment and sensory organs. These fenestrae were present in pairs, one on each side of the skull. At the front of the skull were two large, elliptical nares, which were roughly half the size of the orbits. The orbits were proportionally larger in Massospondylus than in related genera, such as Plateosaurus. The antorbital fenestrae, smaller than those seen in Plateosaurus, were situated between the eyes and the nose. At the rear of the skull were two more pairs of temporal fenestrae: the lateral temporal fenestrae immediately behind the eye sockets, which were shaped like an inverted "T" in Massospondylus,[9] and the supratemporal fenestrae on top of the skull. Small fenestrae also penetrated each mandible.[6] The shape of the skull is traditionally restored as wider and shorter than that of Plateosaurus, but this appearance may be due just to differential crushing experienced by the various specimens.[6] Some features of the skull are variable between individuals; for example, the thickness of the upper border of the orbit and the height of the posterior maxilla. These differences may be due to sexual dimorphism[31] or individual variation.[32]

Tooth count is variable between individuals and increases with skull size.[31] The premaxilla shows the constant number of four teeth per side in all known skulls; however, in the maxilla, the tooth count ranges from 14 to 22. There are 26 teeth in each side of the lower jaw in the largest known skull.[9][31] The height of the teeth crowns decreases from front to back in the upper jaw, but was more or less constant in the lower jaw.[9][31] The lack of pronounced tooth wear and the variable height of the crowns suggests that the teeth were replaced by succeeding new ones in relatively short time intervals.[31] Notably, there was variation in the tooth morphology based on the position of the teeth in the jaw.[31] The heterodonty present in Massospondylus is greater than that present in Plateosaurus, although not as pronounced as the specialization of teeth in Heterodontosaurus.[7] Teeth closer to the front of the snout had round cross-sections and tapered to points, unlike the back teeth, which were spatulate and had oval cross-sections.[6][7]
As with other early sauropodomorphs, it has been proposed that Massospondylus had cheeks. This theory was proposed because there are a few large holes for blood vessels on the surfaces of the jaw bones, unlike the numerous small holes present on the jaws of cheekless reptiles. The cheeks would have prevented food from spilling out when Massospondylus ate.[6] Crompton and Attridge (1986) described skulls of Massospondylus as possessing pronounced overbites and suggested the presence of a horny beak on the tip of the lower jaw to make up the difference in length and account for tooth wear on the teeth at the tip of the snout.[33] However, the difference in length may be a misinterpretation based on crushing in a top–bottom plane, and the possession of a beak is considered unlikely in recent studies.[9][31][34]
Classification
Basal sauropodomorph systematics continue to undergo revision, and many genera once considered classic "prosauropods" have recently been removed from the group in phylogenetic nomenclature, on the grounds that their inclusion would not constitute a clade (a natural grouping containing all descendants of a single common ancestor). Yates and Kitching (2003) published a clade consisting of Riojasaurus, Plateosaurus, Coloradisaurus, Massospondylus, and Lufengosaurus.[35] Galton and Upchurch (2004) included Ammosaurus, Anchisaurus, Azendohsaurus, Camelotia, Coloradisaurus, Euskelosaurus, Jingshanosaurus, Lessemsaurus, Lufengosaurus, Massospondylus, Melanorosaurus, Mussaurus, Plateosaurus, Riojasaurus, Ruehleia, Saturnalia, Sellosaurus, Thecodontosaurus, Yimenosaurus and Yunnanosaurus in a monophyletic Prosauropoda.[6] Wilson (2005) considered Massospondylus, Jingshanosaurus, Plateosaurus, and Lufengosaurus a natural group, with Blikanasaurus and Antetonitrus possible sauropods.[36] Bonnan and Yates (2007) considered Camelotia, Blikanasaurus and Melanorosaurus possible sauropods.[37] Yates (2007) placed Antetonitrus, Melanorosaurus, and Blikanasaurus as basal sauropods and declined to use the term Prosauropoda, as he considered it synonymous with Plateosauridae. However, he did not rule out the possibility that a small group of prosauropods consisting of Plateosaurus, Riojasaurus, Massospondylus and their closest kin were monophyletic.[38]
Massospondylus is the type genus of the proposed family Massospondylidae, to which it gives its name. The Massospondylidae may also include Yunnanosaurus,[39] although Lu et al. (2007) placed Yunnanosaurus in its own family.[40] Yates (2007) considered Massospondylus, Coloradisaurus, and Lufengosaurus massospondylids, with Yunnanosaurus in Anchisauria.[38] Smith and Pol (2007) also found a Massospondylidae in their phylogenetic analysis, including Massospondylus, Coloradisaurus, and Lufengosaurus, as well as their new genus, Glacialisaurus.[41] Adeopapposaurus, based on the fossils once thought to belong to a South American form of Massospondylus, was also classified as a massospondylid,[11] as was Leyesaurus, another South American genus that was named in 2011.[42] Pradhania was originally regarded as a more basal sauropodomorph but new cladistic analysis performed by Novas et al., 2011 suggests that Pradhania is a massospondylid. Pradhania presents two shared traits of the Massospondylidae recovered in their phylogenetic analysis, and the fossils of Pradhania were discovered from the same region and basin in India as M. hislopi.[43]

The following cladogram shows the position of Massospondylus within Massospondylidae, according to Fernando E. Novas and colleagues, 2011:[43]
| Massopoda |
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||
The following cladogram shows the position of Massospondylus within Massopoda, according to Oliver W. M. Rauhut and colleagues, 2020:[44]
| Massopoda |
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Paleobiology

As with all dinosaurs, much of the biology of Massospondylus, including its behavior, coloration, and physiology, remains unknown. However, recent studies have allowed for informed speculation on subjects such as growth patterns,[45] diet,[46] posture,[30] reproduction,[47] and respiration.[48]
A 2007 study suggested that Massospondylus may have used its short arms for defense against predators ("defensive swats"), in intraspecies combat, or in feeding, although its arms were too short to reach its mouth. Scientists speculate that Massospondylus could have used its large pollex (thumb) claw in combat, to strip plant material from trees,[30] digging, or for grooming.[7]
Growth

A 2005 study indicated that Massospondylus' sister taxon, Plateosaurus, exhibited growth patterns affected by environmental factors. The study indicated that, when food was plentiful or when the climate was favorable, Plateosaurus exhibited accelerated growth. This pattern of growth is called "developmental plasticity". It is unseen in other dinosaurs, including Massospondylus, despite the close relationship between the two. The study indicated that Massospondylus grew along a specific growth trajectory, with little variation in the growth rate and ultimate size of an individual.[45] Another study of age determination indicated that Massospondylus grew at a maximum rate of 34.6 kg (76.3 lb) per year and was still growing at around 15 years of age.[49]
Diet
Early sauropodomorphs such as Massospondylus may have been herbivorous or omnivorous. As recently as the 1980s, paleontologists debated the possibility of carnivory in the "prosauropods".[2][8] However, the hypothesis of carnivorous "prosauropods" has been discredited, and all recent studies favor a herbivorous or omnivorous lifestyle for these animals. Galton and Upchurch (2004) found that cranial characteristics (such as jaw articulation) of most basal sauropodomorphs are closer to those of herbivorous reptiles than those of carnivorous ones, and the shape of the tooth crown is similar to those of modern herbivorous or omnivorous iguanas. The maximum width of the crown was greater than that of the root, resulting in a cutting edge similar to those of extant herbivorous or omnivorous reptiles.[6] Barrett (2000) proposed that basal sauropodomorphs supplemented their herbivorous diets with small prey or carrion.[50] Gastroliths (gizzard stones) have been found in association with three Massospondylus fossils from the Forest-Sandstone in Zimbabwe,[51] and with a Massospondylus-like animal from the Late Triassic of Virginia.[46] Until recently, scientists believed that these stones functioned as a gastric mill to aid ingestion of plant material, compensating for its inability to chew, as it is the case in many modern birds. However, Wings and Sander (2007) showed that the polished nature and the abundance of those stones precluded a use as an effective gastric mill in most non-theropod dinosaurs, including Massospondylus.[29][52]
Gait and range of motion

Although long assumed to have been quadrupedal, a 2007 anatomical study of the forelimbs has questioned this, arguing that their limited range of motion precluded effective habitual quadrupedal gait. Neither could the forelimbs swing fore and behind in a fashion similar to the hindlimbs, nor could the hand be rotated with the palmar surfaces facing downwards. This inability to pronate the hand is also supported by in-situ finds of articulated (still-connected) arms that always show unrotated hands with palmar faces facing each other. The study also ruled out the possibility of "knuckle-walking" and other forms of locomotion that would make an effective locomotion possible without the need to pronate the hand. Although its mass suggests a quadrupedal nature, Massospondylus would have been restricted to its hind legs for locomotion.[30]
Since the discovery of rudimentary and nonfunctional clavicles in ceratopsians, it was assumed that these shoulder bones were reduced in all dinosaurs that did not have true furculae.[54] Robert Bakker (1987) suggested that this would have allowed the shoulder blades to swing with the forelimbs in quadrupedal dinosaurs, increasing their functional forelimb length.[55] This would have reduced the discrepancy of length between fore- and hindlimbs in a quadrupedal Massospondylus. However, a recent discovery shows that Massospondylus possessed well-developed clavicles that were joined in a furcula-like arrangement, acting like a clasp between the right and left shoulder blades and prohibiting any rotation of these bones. This discovery indicates that the clavicle reduction is limited to the evolutionary line leading to the ceratopsians. It also indicates that the furcula of birds is derived from clavicles.[54]
Michael Cooper (1981) noted that the zygapophyses of the neck vertebrae were inclined, prohibiting significant horizontal movement of the neck, so that "consequently any significant movement in this direction must have been accomplished by a change in the position of the entire body".[2] This was contradicted in a recent study, noting that only the basalmost cervicals show inclined zygapophyses, allowing sufficient horizontal movement of the neck as a whole.[34]
Reproduction
%252C_Golden_Gate_National_Park%252C_South_Africa%252C_Early_Jurassic_-_Royal_Ontario_Museum_-_DSC00145.JPG.webp)
In 1976, a clutch of seven 190-million-year-old Massospondylus eggs was found in Golden Gate Highlands National Park in South Africa by James Kitching, who identified them as most likely belonging to Massospondylus.[56] It was nearly 30 years before extraction was started on the fossils of the 15-centimetre- (6 in-) long embryos. They remain the oldest dinosaur embryos ever found.[47] By early 2012, at least 10 egg clutches from at least four fossiliferous horizons had been found, with up to 34 eggs per clutch. This indicates that this nesting site may have been used repeatedly (site fidelity), by groups of animals (colonial nesting); in both cases, these represent the oldest evidence of this behaviour.[57] Sedimentary structures indicate that the nesting area was in the vicinity of a lake.[57] The eggshells were very thin (about 0.1 mm), allowing gas exchange even in a low oxygen and carbon dioxide rich environment, which indicates that the eggs were at least partly buried in the substrate.[57] There are no hints that Massospondylus constructed nests; however, the arrangement of the eggs in tight rows indicates that the eggs were pushed in this position by the adults.[57]
The embryos probably represented near-hatchlings.[56] While the skeletal features were similar to those of the adults, the body proportions were very dissimilar. The head was big with a short snout and very large orbits, whose diameter amounts 39% of the entire skull length. The neck was short, contrasting to the very long neck in the adults. Girdle bones and caudals were relatively tiny.[47] The forelimbs were of equal length to the hindlimbs, indicating that newly hatched Massospondylus were quadrupedal, unlike the bipedal adults.[47] However, the reliability of these results has been questioned.[58] The discovery of hatching footprints with manus impressions confirmed their quadrupedality. These impressions show that the hand was not pronated, with palm faces facing each other and the thumb facing forwards.[57] The unpronated manus and the big head indicate that an effective locomotion was not possible for newly hatched Massospondylus.[30] Notably, the near-hatchings had no teeth, suggesting they had no way of feeding themselves.[47] Based on the lack of teeth and the ineffective locomotion, scientists speculate that postnatal care might have been necessary.[30][47] This is further supported by evidence that the hatchings remained at the nest sites until they had doubled in size.[57]

Newly hatched juveniles are known from a second sauropodomorph, Mussaurus; these remains resemble those of the embryonic Massospondylus, suggesting that quadrupedality was present in newly hatched Mussaurus and presumably other basal sauropodomorphs as well.[30][56] The quadrupedality of the hatchings suggests that the quadrupedal posture of later sauropods may have evolved from retention of juvenile characteristics in adult animals, an evolutionary phenomenon known as paedomorphosis.[47] This discovery therefore "sheds some light in the evolutionary pathways through which the peculiar adaptations of giant dinosaurs were attained", stated French paleontologist Eric Buffetaut.[59] However, a recent study, which evaluated locomotion in extinct taxa based on limb robusticity, found that previous conclusions that Massospondylus transitioned from quadrupedality to bipedality through ontogeny were based on unreliable allometric comparisons between limb lengths, but a model based on the circumferences of the humerus and femur supported bipedality throughout ontogeny.[58]
Respiratory system
Many saurischian dinosaurs possessed vertebrae and ribs that contained hollowed-out cavities (pneumatic foramina), which reduced the weight of the bones and may have served as a basic 'flow-through ventilation' system similar to that of modern birds. In such a system, the neck vertebrae and ribs are hollowed out by the cervical air sac; the upper back vertebrae, by the lung; and the lower back and sacral (hip) vertebrae, by the abdominal air sac. These organs constitute a complex and very efficient method of respiration.[60] "Prosauropods" are the only major group of saurischians without an extensive system of pneumatic foramina. Although possible pneumatic indentations have been found in Plateosaurus and Thecodontosaurus, the indentations were very small. One study in 2007 concluded that basal sauropodomorphs like Massospondylus likely had abdominal and cervical air sacs, based on the evidence for them in sister taxa (theropods and sauropods). The study concluded that it was impossible to determine whether basal sauropodomorphs had a bird-like flow-through lung, but that the air sacs were almost certainly present.[48]
Paleoecology
The faunas and floras of the Early Jurassic were similar worldwide, with conifers adapted for hot weather becoming the common plants;[61] basal sauropodomorphs and theropods were the main constituents of a worldwide dinosaur fauna.[62] The environment of early Jurassic southern Africa has been described as a desert.[63] African Massospondylus was a contemporary of temnospondyli; turtles; a sphenodont; rauisuchids; early crocodylomorphs; tritylodontid and trithelodontid therapsids; morganucodontid mammals;[6] and dinosaurs including the small theropod Megapnosaurus and several genera of early ornithischians, such as Lesothosaurus and the heterodontosaurids Abrictosaurus, Heterodontosaurus, Lycorhinus and Pegomastax.[20][64][65] Until recently, Massospondylus was regarded as the only known sauropodomorph from the Upper Elliot Formation.[16] However, newer finds revealed a diverse contemporary sauropodomorph fauna with six additional species, including Ignavusaurus,[66] Arcusaurus[67] and two unnamed taxa as well as two unnamed sauropods.[16]
It is not clear which carnivores may have preyed on Massospondylus. Most of the theropods that have been discovered in rocks of Early Jurassic age in southern Africa, such as Coelophysis, were smaller than mid-sized sauropodomorphs like Massospondylus. These smaller predators have been postulated as using fast slashing attacks to wear down sauropodomorphs, which could have defended themselves with their large hand and foot claws.[63] The 6-metre-(20 ft-) long[68] carnivorous theropod Dracovenator lived during the same period (Hettangian to Sinemurian stages) as Massospondylus and has also been found in the Elliot Formation of South Africa.[69]
References
- Owen, Richard (1854). "Descriptive catalogue of the Fossil organic remains of Reptilia and Pisces contained in the Museum of the Royal College of Surgeons of England." London pp. 80, 97 OCLC 14825172
- Cooper, M. R. (1980). "The prosauropod dinosaur Massospondylus carinatus Owen from Zimbabwe: its biology, mode of life and phylogenetic significance". Occasional Papers of the National Museums and Monuments of Rhodesia, Series B, Natural Sciences. 6 (10): 689–840.
- Yates, Adam M.; Paul M. Barrett (2010). "Massospondylus carinatus Owen 1854 (Dinosauria: Sauropodomorpha) from the Lower Jurassic of South Africa: Proposed conservation of usage by designation of a neotype". Palaeontologia Africana. 45: 7–10.
- Broom, Robert (1911). "On the dinosaurs of the Stormberg, South Africa". Annals of the South African Museum. 7 (4): 291–308.
- Glut, Donald F. (1997). "Massospondylus". Dinosaurs: The Encyclopedia. Jefferson, North Carolina: McFarland & Co. pp. 581–586. ISBN 978-0-89950-917-4.
- Galton, P.M. and Upchurch, P. (2004). "Prosauropoda". Weishampel & als: The Dinosauria (2nd edition), pp. 232–258.
- "Massospondylus." In: Dodson, Peter & Britt, Brooks & Carpenter, Kenneth & Forster, Catherine A. & Gillette, David D. & Norell, Mark A. & Olshevsky, George & Parrish, J. Michael & Weishampel, David B. The Age of Dinosaurs. Publications International, LTD. p. 39. ISBN 0-7853-0443-6.
- Attridge, J.; A. W. Crompton; Farish A. Jenkins, Jr. (1985). "The southern Liassic prosauropod Massospondylus discovered in North America". Journal of Vertebrate Paleontology. 5 (2): 128–132. doi:10.1080/02724634.1985.10011850.
- Sues, H.-D.; R. R. Reisz; S. Hinic; M. A. Raath (2004). "On the skull of Massospondylus carinatus Owen, 1854 (Dinosauria: Sauropodomorpha) from the Elliot and Clarens formations (Lower Jurassic) of South Africa". Annals of Carnegie Museum. 73 (4): 239–257. doi:10.5962/p.316084. S2CID 251542198.
- Rowe, Timothy B.; Hans-Dieter Sues; Robert R. Reisz (2010). "Dispersal and diversity in the earliest North American sauropodomorph dinosaurs, with a description of a new taxon". Proc. R. Soc. B. 278 (1708): 1044–53. doi:10.1098/rspb.2010.1867. PMC 3049036. PMID 20926438.
- Martínez, Ricardo N. (2009). "Adeopapposaurus mognai, gen. et sp. nov (Dinosauria: Sauropodomorpha), with comments on adaptations of basal sauropodomorpha". Journal of Vertebrate Paleontology. 29 (1): 142–164. doi:10.1671/039.029.0102. S2CID 85074392.
- Chapelle, Kimberley E.J.; Barrett, Paul M.; Botha, Jennifer; Choiniere, Jonah N. (2019). "Ngwevu intloko: a new early sauropodomorph dinosaur from the Lower Jurassic Elliot Formation of South Africa and comments on cranial ontogeny in Massospondylus carinatus". PeerJ. 7: e7240. doi:10.7717/peerj.7240. PMC 6687053. PMID 31403001.
- "Massospondylus". The Paleobiology Database. 2007. Retrieved 2007-11-03.
- Seeley, H.G. (1895). "On the type of the genus Massospondylus and on some Vertebrae and limb-bone of M. (?) browni". Annals and Magazine of Natural History. 15 (85): 102–125. doi:10.1080/00222939508677852.
- Lydekker, Richard (1890). "Note on certain vertebrate remains from the Nagpur district". Records of the Geological Survey of India. 23 (1): 21–24.
- Barrett, P. M. (2009). "A new basal sauropodomorph dinosaur from the Upper Elliot Formation (Lower Jurassic) of South Africa". Journal of Vertebrate Paleontology. 29 (4): 1032–1045. doi:10.1671/039.029.0401. S2CID 128489192.
- Haughton, Sydney H. (1924). "The fauna and stratigraphy of the Stormberg Series". Annals of the South African Museum. 12: 323–497.
- Weishampel, David B. (1990). "Dinosaur distribution". In David B. Weishampel; Peter Dodson; Halszka Osmólska (eds.). The Dinosauria (1st ed.). Berkeley: University of California Press. pp. 63–139. ISBN 978-0-520-06727-1.
- Molnar, R. E. (1990). Problematic Theropoda: "Carnosaurs". In Weishampel & als: The Dinosauria (1st ed.), pp. 306–317.
- Weishampel, David B.; Paul M. Barrett; Rodolfo Coria; Jean Le Loeuff; Zhao Xijin Xu Xing; Ashok Sahni; Elizabeth M. P. Gomani; Christopher R. Noto (2004). "Dinosaur Distribution". In David B. Weishampel; Peter Dodson; Halszka Osmólska (eds.). The Dinosauria (2nd ed.). Berkeley: University of California Press. pp. 517–606. ISBN 978-0-520-24209-8.
- Yates, Adam M.; Matthew F. Bonnan; Johann Neveling (2011). "A new basal sauropodomorph dinosaur from the Early Jurassic of South Africa". Journal of Vertebrate Paleontology. 31 (3): 610–625. doi:10.1080/02724634.2011.560626. S2CID 85589914.
- Chapelle, KEJ; Choiniere, JN (2018). "A revised cranial description of Massospondylus carinatus Owen (Dinosauria: Sauropodomorpha) based on computed tomographic scans and a review of cranial characters for basal Sauropodomorpha". PeerJ. 6: e4224. doi:10.7717/peerj.4224. PMC 5768178. PMID 29340238.
- Chapelle, KEJ; Barrett, PM; Botha, J; Choiniere, JN (2019). "Ngwevu intloko: a new early sauropodomorph dinosaur from the Lower Jurassic Elliot Formation of South Africa and comments on cranial ontogeny in Massospondylus carinatus". PeerJ. 7: e7240. doi:10.7717/peerj.7240. PMC 6687053. PMID 31403001.
- Lambert, David; the Diagram Group (1983). A Field Guide to Dinosaurs. New York: Avon Books. p. 103. ISBN 978-0-380-83519-5.
- Lambert, David; the Diagram Group (1990). The Dinosaur Data Book. New York: Avon Books. p. 75. ISBN 978-0-380-75896-8.
- Seebacher, Frank (2001). "A new method to calculate allometric length-mass relationships of dinosaurs" (PDF). Journal of Vertebrate Paleontology. 21 (1): 51–60. CiteSeerX 10.1.1.462.255. doi:10.1671/0272-4634(2001)021[0051:ANMTCA]2.0.CO;2. S2CID 53446536.
- Lessem, Don; Donald F. Glut (1993). "Massospondylus". The Dinosaur Society Dinosaur Encyclopedia. Random House. p. 295. ISBN 978-0-679-41770-5.
- Glut, Donald F. (2000). "Massospondylus". Dinosaurs: The Encyclopedia: Supplement One. Jefferson, North Carolina: McFarland & Co. p. 258. ISBN 978-0-7864-0591-6.
- Yates, Adam M. (2012). "Basal Sauropodomorpha: The "Prosauropods"". In M. K. Brett-Surman; James O. Farlow; Thomas R. Holtz (eds.). The Complete Dinosaur (2 ed.). Indiana University Press. pp. 430, 435. ISBN 978-0-253-35701-4.
- Bonnan, Matthew F.; Phil Senter (2007). "Were the basal sauropodomorph dinosaurs Plateosaurus and Massospondylus habitual quadrupeds?". In Paul M. Barrett; D. J. Batten (eds.). Evolution and Palaeobiology of Early Sauropodomorph Dinosaurs. Special Papers in Palaeontology. London: The Palaeontological Association. pp. 139–155. ISBN 978-1-4051-6933-2.
- Gow, Christoper E.; J. W. Kitching; Michael K. Raath (1990). "Skulls of the prosauropod dinosaur Massospondylus carinatus Owen in the collections of the Bernard Price Institute for Palaeontological Research". Palaeontologia Africana. 27: 45–58.
- Galton, Peter M. (1997). "Comments on sexual dimorphism in the prosauropod dinosaur Plateosaurus engelhardti (Upper Triassic, Trossingen)". Neues Jahrbuch für Geologie und Paläontologie, Monatshefte. 1997 (11): 674–682. doi:10.1127/njgpm/1997/1997/674.
- Crompton, A.W.; John Attridge (1986). "Masticatory apparatus of the larger herbivores during Late Triassic and Early Jurassic times". In Kevin Padian (ed.). The Beginning of the Age of Dinosaurs: Faunal Change Across the Triassic–Jurassic Boundary. New York: Cambridge University Press. pp. 223–236. ISBN 978-0-521-36779-0.
- Barrett, Paul; Paul Upchurch (2007). "The evolution of feeding mechanisms in early sauropodomorph dinosaurs". In Paul M. Barrett; D. J. Batten (eds.). Evolution and Palaeobiology of Early Sauropodomorph Dinosaurs. Special Papers in Palaeontology. London: The Palaeontological Association. pp. 91–112. ISBN 978-1-4051-6933-2.
- Yates, Adam M.; James W. Kitching (2003). "The earliest known sauropod dinosaur and the first steps towards sauropod locomotion". Proceedings of the Royal Society B. 270 (1525): 1753–1758. doi:10.1098/rspb.2003.2417. PMC 1691423. PMID 12965005.
- Wilson, Jeffrey A.; Kristina Curry Rogers (2005). The Sauropods: Evolution and Paleobiology. Berkeley: University of California Press. pp. 15–49. ISBN 978-0-520-24623-2.
- Bonnan, Matthew F. and Adam M. Yates, "A new description of the forelimbs of the basal sauropodomorph Melanorosaurus: implications for the evolution of pronation, manus shape and quadrupedalism in sauropod dinosaurs". In Barrett & Batten (eds.), Evolution and Palaeobiology (2007), pp. 157–168.
- Yates, Adam M., "The first complete skull of the Triassic dinosaur Melanorosaurus Haughton (Sauropodomorpha: Anchisauria)". In Barrett & Batten (eds.), Evolution and Palaeobiology (2007), pp. 9–55.
- Sereno, Paul C. (1999). "The evolution of dinosaurs". Science. 284 (5423): 2137–2147. doi:10.1126/science.284.5423.2137. PMID 10381873.
- Lu, J.; T. Li; S. Zhong; Y. Azuma; M. Fujita; Z Dong; Q. Ji (2007). "New yunnanosaurid dinosaur (Dinosauria, Prosauropoda) From the Middle Jurassic Zhanghe Formation of Yuanmou, Yunnan province of China". Memoir of the Fukui Prefectural Dinosaur Museum. 6: 1–15.
- Smith, Nathan D.; Diego Pol (2007). "Anatomy of a basal sauropodomorph dinosaur from the Early Jurassic Hanson Formation of Antarctica" (PDF). Acta Palaeontologica Polonica. 52 (4): 657–674.
- Apaldetti, Cecilia; Ricardo N. Martinez; Oscar A. Alcober; Diego Pol (2011). "A New Basal Sauropodomorph (Dinosauria: Saurischia) from Quebrada del Barro Formation (Marayes-El Carrizal Basin), Northwestern Argentina". PLOS ONE. 6 (1): e26964. doi:10.1371/journal.pone.0026964.g002.
- Novas, Fernando E.; Martin D. Ezcurra; Sankar Chatterjee; T. S. Kutty (2011). "New dinosaur species from the Upper Triassic Upper Maleri and Lower Dharmaram formations of central India". Earth and Environmental Science Transactions of the Royal Society of Edinburgh. 101 (3–4): 333–349. doi:10.1017/S1755691011020093. S2CID 128620874.
- Rauhut, O. W. M.; Holwerda, F. M.; Furrer, H. (2020). "A derived sauropodiform dinosaur and other sauropodomorph material from the Late Triassic of Canton Schaffhausen, Switzerland". Swiss Journal of Geosciences. 113 (1): 8. doi:10.1186/s00015-020-00360-8. S2CID 220294939.
- Sander, P. Martin; Nicole Klein (2005). "Developmental plasticity in the life history of a prosauropod dinosaur". Science. 310 (5755): 1800–1802. Bibcode:2005Sci...310.1800S. doi:10.1126/science.1120125. PMID 16357257. S2CID 19132660.
- Weems, Robert E.; Michelle J. Culp; Oliver Wings (2007). "Evidence for Prosauropod Dinosaur Gastroliths in the Bull Run Formation (Upper Triassic, Norian) of Virginia". Ichnos. 13 (3–4): 271–295. doi:10.1080/10420940601050030. S2CID 128683430.
- Reisz, Robert R.; Diane Scott; Hans-Dieter Sues; David C. Evans; Michael A. Raath (2005). "Embryos of an Early Jurassic prosauropod dinosaur and their evolutionary significance" (PDF). Science. 309 (5735): 761–764. Bibcode:2005Sci...309..761R. doi:10.1126/science.1114942. PMID 16051793. S2CID 37548361.
- Wedel, Mathew (2007). "What pneumaticity tells us about 'prosauropods', and vice versa" (PDF). Special Papers in Palaeontology. 77: 207–222. Archived from the original (PDF) on 2008-07-05. Retrieved 2007-10-31.
- Erickson, Gregory M.; Kristina Curry Rogers; Scott A. Yerby (2001). "Dinosaurian growth patterns and rapid avian growth rates". Nature. 412 (6845): 429–433. Bibcode:2001Natur.412..429E. doi:10.1038/35086558. PMID 11473315. S2CID 4319534.
- Barrett, P.M. (2000). "Prosauropod dinosaurs and iguanas: Speculations on the diets of extinct reptiles". In Hans-Dieter Sues (ed.). Evolution of Herbivory in Terrestrial Vertebrates: Perspectives from the Fossil Record. Cambridge: Cambridge University Press. pp. 42–78. ISBN 978-0-521-59449-3.
- Raath, Michael A. (1974). "Fossil vertebrate studies in Rhodesia: further evidence of gastroliths in prosauropod dinosaurs". Arnoldia Rhodesia.
- Wings, Oliver; P. Martin Sander (2007). "No gastric mill in sauropod dinosaurs: new evidence from analysis of gastrolith mass and function in ostriches". Proceedings of the Royal Society B: Biological Sciences. 274 (1610): 635–640. doi:10.1098/rspb.2006.3763. PMC 2197205. PMID 17254987.
- Lindsay, William; Nigel Zarkin; Neil Smith (1996). "Displaying Dinosaurs at The Natural History Museum, London". Curator: The Museum Journal. 39 (4): 262–279. doi:10.1111/j.2151-6952.1996.tb01102.x. ISSN 2151-6952.
- Yates, Adam M.; Cecilio C. Vasconcelos (2005). "Furcula-like clavicles in the prosauropod dinosaur Massospondylus". Journal of Vertebrate Paleontology. 25 (2): 466–468. doi:10.1671/0272-4634(2005)025[0466:FCITPD]2.0.CO;2. ISSN 0272-4634. S2CID 85679726.
- Bakker, R. T. (1987). "The return of the dancing dinosaurs". Dinosaurs Past and Present. 1: 38–69.
- Reisz, Robert R.; David C. Evans; Hans-Dieter Sues; Diane Scott (2010-11-01). "Embryonic Skeletal Anatomy of the Sauropodomorph Dinosaur Massospondylus from the Lower Jurassic of South Africa". Journal of Vertebrate Paleontology. 30 (6): 1653, 1664. doi:10.1080/02724634.2010.521604. ISSN 0272-4634. S2CID 84599991.
- Reisz, Robert R.; David C. Evans; Eric M. Roberts; Hans-Dieter Sues; Adam M. Yates (2012). "Oldest known dinosaurian nesting site and reproductive biology of the Early Jurassic sauropodomorph Massospondylus". Proceedings of the National Academy of Sciences of the United States of America. 109 (7): 2428–2433. Bibcode:2012PNAS..109.2428R. doi:10.1073/pnas.1109385109. PMC 3289328. PMID 22308330.
- Chapelle, Kimberley E. J.; Benson, Roger B. J.; Stiegler, Josef; Otero, Alejandro; Zhao, Qi; Choiniere, Jonah N. (March 2020). "A quantitative method for inferring locomotory shifts in amniotes during ontogeny, its application to dinosaurs and its bearing on the evolution of posture". Palaeontology. 63 (2): 229–242. doi:10.1111/pala.12451. ISSN 0031-0239. S2CID 210278749.
- Stokstad, E. (2005). "Dinosaur embryos hint at evolution of giants". Science. 309 (5735): 679. doi:10.1126/science.309.5735.679. PMID 16051755. S2CID 83313135.
- O'Connor, Patrick M.; Leon P.A.M. Claessens (2006). "Basic avian pulmonary design and flow-through ventilation in non-avian theropod dinosaurs". Nature. 436 (7048): 253–256. Bibcode:2005Natur.436..253O. doi:10.1038/nature03716. PMID 16015329. S2CID 4390587.
- Russell, Dale A. (1989). An Odyssey in Time: Dinosaurs of North America. Minocqua, Wisconsin: NorthWord Press. p. 45. ISBN 978-1-55971-038-1.
- Holtz, Thomas R. Jr.; Chapman, Ralph E.; Lamanna, Matthew C. (2004). "Mesozoic biogeography of Dinosauria". In Weishampel, David B; Dodson, Peter (eds.). The Dinosauria (2nd ed.). University of California Press. pp. 627–642. ISBN 978-0-520-24209-8.
- Paul, Gregory S. (1988). "Coelophysis rhodesiensis". Predatory Dinosaurs of the World. New York: Simon and Schuster. pp. 262–265. ISBN 978-0-671-61946-6.
- Knoll, F. (2005). "The tetrapod fauna of the Upper Elliot and Clarens formations in the main Karoo Basin (South Africa and Lesotho)". Bulletin de la Société Géologique de France. 176 (1): 81–91. doi:10.2113/176.1.81.
- Sereno, Paul (2012-10-03). "Taxonomy, morphology, masticatory function and phylogeny of heterodontosaurid dinosaurs". ZooKeys (226): 1–225. doi:10.3897/zookeys.226.2840. ISSN 1313-2970. PMC 3491919. PMID 23166462.
- Knoll, F. (2010). "A primitive sauropodomorph from the upper Elliot Formation of Lesotho". Geological Magazine. 147 (6): 814–829. Bibcode:2010GeoM..147..814K. doi:10.1017/S001675681000018X. S2CID 129174260. Archived from the original on 2013-01-12.
- Yates, A. M.; M. F. Bonnan; J. Neveling (2011). "A new basal sauropodomorph dinosaur from the Early Jurassic of South Africa". Journal of Vertebrate Paleontology. 31 (3): 610–625. doi:10.1080/02724634.2011.560626. S2CID 85589914.
- Smith, N. D.; P. J. Makovicky; D. Pol; W. R. Hammer; P. J. Currie (2007). "The Dinosaurs of the Early Jurassic Hanson Formation of the Central Transantarctic Mountains: Phylogenetic Review and Synthesis" (PDF). In Alan Cooper; Carol Raymond; et al. (eds.). Antarctica: A Keystone in a Changing World—Online Proceedings for the Tenth International Symposium on Antarctic Earth Sciences. Reston, Virginia: U.S. Geological Survey. doi:10.3133/of2007-1047.srp003. ISBN 978-1-4113-1788-8. OCLC 85782509.
- Yates, A.M. (2006). "A new theropod dinosaur from the Early Jurassic of South Africa and its implications for the early evolution of theropods". Palaeontologia Africana. 41: 105–122.
Further reading
- Chinsamy, A. (1992). "Ontogenetic growth of the dinosaurs Massospondylus carinatus and Syntarsus rhodesiensis". In: Abstracts of papers. Society of Vertebrate Paleontology, fifty-second annual meeting. Royal Ontario Museum Toronto, Ontario". Journal of Vertebrate Paleontology. 12 (3): 23A. doi:10.1080/02724634.1992.10011483.
- Gow, C. E. (1990). "Morphology and growth of the Massospondylus braincase (Dinosauria, Prosauropoda)". Palaeontologia Africana. 27: 59–75.
- Hinic, S. (2002). "The cranial anatomy of Massospondylus carinatus Owen, 1854 and its implications for prosauropod phylogeny". Journal of Vertebrate Paleontology. Abstracts of papers. Society of Vertebrate Paleontology, 22, Supplement to number 3, 65A.
- Martínez, R. (1999). "The first South American record of Massospondylus (Dinosauria: Sauropodomorpha)". Journal of Vertebrate Paleontology, Abstracts of papers. Society of Vertebrate Paleontology, 20–23 October, 19, Suppl. 3, 61A.
- Martínez, R.N. (1999). "Massospondylus (Dinosauria: Sauropodomorpha) in northwestern Argentina". Abstracts VII International Symposium on Mesozoic Terrestrial Ecosystems, Buenos Aires, 40.
External links
- Locomotion and Ontogeny of Massospondylus carinatus from the University of Toronto at Missisauga.
- Massospondylidae from Palaeos.com (technical).
- Massospondylus from PrimeOrigins.co.za (for children).
- "Oldest known dinosaur embryos identified" University of the Witwatersrand.
- "Rare fossil embryos reveal growth". Archived from the original on May 3, 2007. Retrieved October 26, 2007. National Geographic.









| |||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||