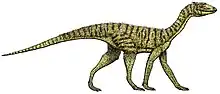
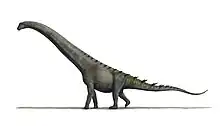
Sauropodomorpha
Sauropodomorpha (/ˌsɔːrəˌpɒdəˈmɔːrfə/[1] SOR-ə-POD-ə-MOR-fə; from Greek, meaning "lizard-footed forms") is an extinct clade of long-necked, herbivorous, saurischian dinosaurs that includes the sauropods and their ancestral relatives. Sauropods generally grew to very large sizes, had long necks and tails, were quadrupedal, and became the largest animals to ever walk the Earth. The prosauropods, which preceded the sauropods, were smaller and were often able to walk on two legs. The sauropodomorphs were the dominant terrestrial herbivores throughout much of the Mesozoic Era, from their origins in the Late Triassic (approximately 230 Ma) until their decline and extinction at the end of the Cretaceous.
| Sauropodomorphs Temporal range: | |
|---|---|
 | |
| Montage of nine different representatives of sauropodomorph dinosaurs. Clockwise from upper left: Eoraptor lunensis, Plateosaurus engelhardti, Brontosaurus excelsus, Europasaurus holgeri, Mamenchisaurus hochuanensis, Nigersaurus taqueti, Argentinosaurus huinculensis, Diplodocus carnegii, Brachiosaurus altithorax. | |
| Scientific classification | |
| Domain: | Eukaryota |
| Kingdom: | Animalia |
| Phylum: | Chordata |
| Clade: | Dinosauria |
| Clade: | Saurischia |
| Clade: | Eusaurischia |
| Clade: | †Sauropodomorpha Huene, 1932 |
| Subgroups | |
| |
Description
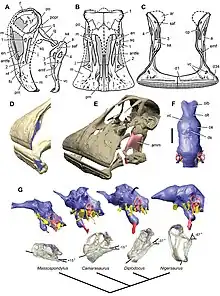
Sauropodomorphs were adapted to browsing higher than any other contemporary herbivore, giving them access to high tree foliage. This feeding strategy is supported by many of their defining characteristics, such as: a light, tiny skull on the end of a long neck (with ten or more elongated cervical vertebrae) and a counterbalancing long tail (with one to three extra sacral vertebrae).
Their teeth were weak, and shaped like leaves or spoons (lanceolate or spatulate). Instead of grinding teeth, they had stomach stones (gastroliths), similar to the gizzard stones of montul birds and crocodiles, to help digest tough plant fibers. The front of the upper mouth bends down in what may be a beak.
One of the earliest known sauropodomorphs, Saturnalia, was small and slender (1.5 metres, or 5 feet long); but, by the end of the Triassic, they were the largest dinosaurs of their time, and throughout the Jurassic and Cretaceous they kept on growing. Ultimately the largest sauropods, like Supersaurus, Diplodocus hallorum, Patagotitan, and Argentinosaurus, reached 30–40 metres (98–131 ft) in length, and 60,000–100,000 kilograms (65–110 US short tons) or more in mass.
Initially bipedal, as their size increased they evolved a four-legged graviportal gait adapted only to walking slowly on land, like elephants. The early sauropodomorphs were most likely omnivores as their shared common ancestor with the other saurischian lineage (the theropods) was a carnivore. Therefore, their evolution to herbivory went hand in hand with their increasing size and neck length.
They also had large nostrils (nares), and retained a thumb (pollex) with a big claw, which may have been used for defense — though their primary defensive adaptation was their extreme size.
Distinguishing anatomical features
Sauropodomorphs can be distinguished as a group on the basis of some of the following synapomorphies:[2]
- The presence of large nares.
- The distal part of the tibia is covered by an ascending process of the astragalus.
- Their hind limbs are short when compared to their torso length.
- The presence of three or more sacral vertebrae.
- The teeth are thin, flat and are spatula-like, with bladed and serrated crowns.
- The presence of a minimum of 10 cervical vertebrae that are typically elongated
- The presence of 25 presacral vertebrae
- The manus had a large digit I.
Evolutionary history
Among the first dinosaurs to evolve in the Late Triassic Period,[3] about 230 million years ago (Mya), they became the dominant herbivores by halfway through the late Triassic (during the Norian stage). Their perceived decline in the early Cretaceous is most likely a bias in fossil sampling, as most fossils are known from Europe and North America, but sauropods were still the dominant herbivores in the Gondwanan landmasses. The spread of flowering plants (angiosperms) and "advanced" ornithischians, another major group of herbivorous dinosaurs (noted for their highly developed chewing mechanisms), are most likely not a major factor in sauropod decline in the northern continents. Like all non-avian dinosaurs, the sauropodomorphs became extinct 66 Mya, during the Cretaceous–Paleogene extinction event.
The earliest and most basal sauropodomorphs known are Chromogisaurus novasi and Panphagia protos, both from the Ischigualasto Formation, dated to 231.4 million years ago (late Carnian age of the Late Triassic according to the ICS[4][5]). Some studies have found Eoraptor lunensis (also from the Ischigualasto Formation), traditionally considered a theropod, to be an early member of the sauropodomorph lineage, which would make it the most basal sauropodomorph known.[6]
Classification


Sauropodomorpha is one of the two major clades within the order Saurischia. The sauropodomorphs' sister group, the Theropoda, includes bipedal carnivores like Velociraptor and Tyrannosaurus; as well as birds. However, sauropodomorphs also share a number of characteristics with the Ornithischia, so a small minority of palaeontologists, like Bakker, have historically placed both sets of herbivores within a group called "Phytodinosauria" or "Ornithischiformes".
In Linnaean taxonomy, Sauropodomorpha (which means "lizard feet forms") is either a suborder or is left unranked. It was originally established by Friedrich von Huene in 1932, who broke it into two groups: the basal forms within Prosauropoda, and their descendants, the giant Sauropoda.
Phylogenetic analyses by Adam Yates (2004, 2006) and others firmly placed Sauropoda within a paraphyletic "Prosauropoda". Recent cladistic analyses suggest that the clade Prosauropoda, which was named by Huene in 1920 and was defined by Sereno, in 1998, as all animals more closely related to Plateosaurus engelhardti than to Saltasaurus loricatus,[7] is a junior synonym of Plateosauridae as both contain the same taxa.[8][9]
Most modern classification schemes break the prosauropods into a half-dozen groups that evolved separately from one common lineage. While they have a number of shared characteristics, the evolutionary requirements for giraffe-like browsing high in the trees may have caused convergent evolution, where similar traits evolve separately because they faced the same evolutionary pressure, instead of (homologous) traits derived from a shared ancestor.[10]
Phylogeny

Cladogram after Novas et al., 2011:[9]
| Sauropodomorpha |
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
The phylogenetic analysis of Otero et al., 2015 found Sauropodomorpha to be in a polytomy with Agnosphitys and Theropoda within Eusaurischia, with Herrerasauridae and Eoraptor external to it within Saurischia.[11] A large phylogenetic analysis of early dinosaurs published by Matthew Baron, David Norman and Paul Barrett (2017) in the journal Nature redefined Sauropodomorpha and Saurischia and recovered Herrerasauridae as the sister group to Sauropodomorpha within Saurischia. This resulted from the proposed removal of Theropoda from Saurischia and the formation of Ornithoscelida, a clade containing Theropoda and Ornithischia.[12]
Below is a cladogram of basal sauropodomorpha after Müller, 2019.[13]
| Sauropodomorpha |
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Paleobiology
Comparisons between the scleral rings of several sauropodomorph genera (Diplodocus, Lufengosaurus, Nemegtosaurus, Plateosaurus, and Riojasaurus) and modern birds and reptiles suggest that they may have been cathemeral, active throughout the day at short intervals.[14]
Sauropodomorphs reached the age of sexual maturity well before they were fully-grown adults.[15][16] A study by Griebeler et al. (2013) concluded that the maximum growth rates of sauropodomorphs were comparable to those of precocial birds and the black rhinoceros but lower than the growth rates of average mammals.[16]
A long-standing hypothesis has been that early sauropodomorphs were carnivorous, as expected for most early dinosaurs. This hypothesis is supported by the current basalmost sauropodomorph, Buriolestes, and its plesiomorphic, theropod-like teeth.[17] The teeth of more derived sauropodomorphs such as Eoraptor, Panphagia, and Pampadromaeus are better-suited for herbivorous (or possibly omnivorous) diets.[18] Regardless of the phylogenetic position of herrerasaurids or Eoraptor (which are in flux),[18][19][20] ancestral state reconstructions recover carnivory as being ancestral to sauropodomorphs.[17]
References
- Cf. "sauropodomorph - definition of sauropodomorph in English from the Oxford dictionary". OxfordDictionaries.com. Archived from the original on November 1, 2015. Retrieved 2016-01-20.
- Martin, A.J. (2006). Introduction to the Study of Dinosaurs. Second Edition. Oxford, Blackwell Publishing. pg. 299-300. ISBN 1-4051-3413-5.
- Müller, Rodrigo T.; Garcia, Maurício S. (2019-03-08). "Rise of an empire: analysing the high diversity of the earliest sauropodomorph dinosaurs through distinct hypotheses". Historical Biology. 32 (10): 1334–1339. doi:10.1080/08912963.2019.1587754. ISSN 0891-2963. S2CID 92177386.
- Luciano A. Leal; Sergio A. K. Azevodo; Alexander W. A. Kellner & Átila A. S. da Rosa (October 18, 2004). "A new early dinosaur (Sauropodomorpha) from the Caturrita Formation (Late Triassic), Paraná Basin, Brazil" (PDF). Zootaxa. 690: 1–24. doi:10.11646/zootaxa.690.1.1. (Warning: abstract is 12 kb PDF)
- Martínez, Ricardo N.; Alcober, Oscar A. (2009). Sereno, Paul (ed.). "A basal sauropodomorph (Dinosauria: Saurischia) from the Ischigualasto Formation (Triassic, Carnian) and the early evolution of Sauropodomorpha". PLOS ONE. 4 (2): 1–12. Bibcode:2009PLoSO...4.4397M. doi:10.1371/journal.pone.0004397. PMC 2635939. PMID 19209223.
- Ricardo N. Martinez; Paul C. Sereno; Oscar A. Alcober; Carina E. Colombi; Paul R. Renne; Isabel P. Montañez & Brian S. Currie (2011). "A Basal Dinosaur from the Dawn of the Dinosaur Era in Southwestern Pangaea". Science. 331 (6014): 206–210. Bibcode:2011Sci...331..206M. doi:10.1126/science.1198467. hdl:11336/69202. PMID 21233386. S2CID 33506648.
- Sereno, P.C. (1998). "A rationale for phylogenetic definitions, with applications to the higher-level taxonomy of Dinosauria". Neues Jahrbuch für Geologie und Paläontologie, Abhandlungen. 210: 41–83. doi:10.1127/njgpa/210/1998/41.
- Yates, Adam M. (2007). Barrett, Paul M.; Batten, David J. (eds.). "The first complete skull of the Triassic dinosaur Melanorosaurus Haughton (Sauropodomorpha: Anchisauria)". Evolution and Palaeobiology: 9–55.
- Fernando E. Novas; Martin D. Ezcurra; Sankar Chatterjee & T. S. Kutty (2011). "New dinosaur species from the Upper Triassic Upper Maleri and Lower Dharmaram formations of central India". Earth and Environmental Science Transactions of the Royal Society of Edinburgh. 101 (3–4): 333–349. doi:10.1017/S1755691011020093. S2CID 128620874.
- Pol D.; Garrido A.; Cerda I.A. (2011). Farke, Andrew Allen (ed.). "A New Sauropodomorph Dinosaur from the Early Jurassic of Patagonia and the Origin and Evolution of the Sauropod-type Sacrum". PLOS ONE. 6 (1): e14572. Bibcode:2011PLoSO...614572P. doi:10.1371/journal.pone.0014572. PMC 3027623. PMID 21298087.
- Otero, Alejandro; Krupandan, Emil; Pol, Diego; Chinsamy, Anusuya; Choiniere, Jonah (2015). "A new basal sauropodiform from South Africa and the phylogenetic relationships of basal sauropodomorphs". Zoological Journal of the Linnean Society. 174 (3): 589. doi:10.1111/zoj.12247.
- Baron, M.G.; Norman, D.B.; Barrett, P.M. (2017). "A new hypothesis of dinosaur relationships and early dinosaur evolution". Nature. 543 (7646): 501–506. Bibcode:2017Natur.543..501B. doi:10.1038/nature21700. PMID 28332513. S2CID 205254710.
- Müller, Rodrigo Temp (2019). "Craniomandibular osteology of Macrocollum itaquii (Dinosauria: Sauropodomorpha) from the Late Triassic of southern Brazil". Journal of Systematic Palaeontology. 18 (10): 805–841. doi:10.1080/14772019.2019.1683902. S2CID 209575985.
- Schmitz, L.; Motani, R. (2011). "Nocturnality in Dinosaurs Inferred from Scleral Ring and Orbit Morphology". Science. →→→§332 (6030): 705–8. Bibcode:2011Sci...332..705S. doi:10.1126/science.1200043. PMID 21493820. S2CID 33253407.
- Sander, PM (2000). "Long bone histology of the Tendaguru sauropods: implications for growth and biology" (PDF). Paleobiology. 26 (3): 466–488. doi:10.1666/0094-8373(2000)026<0466:lhotts>2.0.co;2. S2CID 86183725.
- Griebeler, EM; Klein, N; Sander, PM (2013). "Aging, Maturation and Growth of Sauropodomorph Dinosaurs as Deduced from Growth Curves Using Long Bone Histological Data: An Assessment of Methodological Constraints and Solutions". PLOS ONE. 8 (6): e67012. Bibcode:2013PLoSO...867012G. doi:10.1371/journal.pone.0067012. PMC 3686781. PMID 23840575.
- Cabreira, S.F.; Kellner, A.W.A.; Dias-da-Silva, S.; da Silva, L.R.; Bronzati, M.; de Almeida Marsala, J.C.; Müller, R.T.; de Souza Bittencourt, J.; Batista, B.J.; Raugust, T.; Carrilho, R.; Brodt, A.; Langer, M.C. (2016). "A Unique Late Triassic Dinosauromorph Assemblage Reveals Dinosaur Ancestral Anatomy and Diet". Current Biology. 26 (22): 3090–3095. doi:10.1016/j.cub.2016.09.040. PMID 27839975.
- Sereno, Paul C.; Martínez, Ricardo N.; Alcober, Oscar A. (2013). "Osteology of Eoraptor lunensis (Dinosauria, Sauropodomorph a). Basal sauropodomorphs and the vertebrate fossil record of the Ischigualasto Formation (Late Triassic: Carnian-Norian) of Argentina". Journal of Vertebrate Paleontology Memoir. 12: 83–179. doi:10.1080/02724634.2013.820113. S2CID 86006363.
- Sues, Hans-Dieter; Nesbitt, Sterling J.; Berman, David S.; Henrici, Amy C. (2011). "A late-surviving basal theropod dinosaur from the latest Triassic of North America". Proceedings of the Royal Society B. 278 (1723): 3459–3464. doi:10.1098/rspb.2011.0410. PMC 3177637. PMID 21490016.
- Hendrickx, C.; Hartman, S.A.; Mateus, O. (2015). "An Overview of Non- Avian Theropod Discoveries and Classification". PalArch's Journal of Vertebrate Palaeontology. 12 (1): 1–73.
Sources
- Wilson J. A. (2002). "Sauropod dinosaur phylogeny: critique and cladistic analysis" (PDF). Zoological Journal of the Linnean Society. 136 (2): 217–276. doi:10.1046/j.1096-3642.2002.00029.x.
External links
- Sauropodomorpha: Overview, from Palæos.
- Sauropodmorpha Archived 2019-07-02 at the Wayback Machine, from When Dinosaurs Ruled Texas, by Jon A. Baskin.
- Geol 104 Dinosaurs: A natural history: Sauropodomorpha: Size matters, by Thomas R. Holtz Jr., from the University of Maryland.
 Data related to Sauropodomorpha at Wikispecies
Data related to Sauropodomorpha at Wikispecies
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||