Ancient protein
Ancient proteins are complex mixtures and the term palaeoproteomics is used to characterise the study of proteomes in the past.[1] Ancients proteins have been recovered from a wide range of archaeological materials, including bones,[2] teeth,[3] eggshells,[4] leathers,[5] parchments,[6] ceramics,[7] painting binders[8] and well-preserved soft tissues like gut intestines.[9] These preserved proteins have provided valuable information about taxonomic identification, evolution history (phylogeny), diet, health, disease, technology and social dynamics in the past.
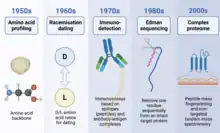
Like modern proteomics, the study of ancient proteins has also been enabled by technological advances. Various analytical techniques, for example, amino acid profiling, racemisation dating, immunodetection, Edman sequencing, peptide mass fingerprinting, and tandem mass spectrometry have been used to analyse ancient proteins.[10] The introduction of high-performance mass spectrometry (for example, Orbitrap) in 2000 has revolutionised the field, since the entire preserved sequences of complex proteomes can be characterised.[11]
Over the past decade, the study of ancient proteins has evolved into a well-established field in archaeological science. However, like the research of aDNA (ancient DNA preserved in archaeological remains), it has been limited by several challenges such as the coverage of reference databases, identification, contamination and authentication.[12] Researchers have been working on standardising sampling, extraction, data analysis and reporting for ancient proteins.[13] Novel computational tools such as de novo sequencing[14] and open research[15] may also improve the identification of ancient proteomes.
History: the pioneers of ancient protein studies
Philip Abelson, Edgar Hare and Thomas Hoering
Abelson, Hare and Hoering were leading the studies of ancient proteins between the 1950s and the early 1970s.[16] Abelson was directing the Geophysical Laboratory at the Carnegie Institute (Washington, DC) between 1953 and 1971, and he was the first to discover amino acids in fossils.[17] Hare joined the team and specialised in amino acid racemisation (the conversion of L- to D-amino acids after the death of organisms). D/L ratios were used to date various ancient tissues such as bones, shells and marine sediments.[18] Hoering was another prominent member, contributing to the advancement of isotopes and mass spectrometry.[19] This golden trio drew many talented biologists, geologists, chemists and physicists to the field, including Marilyn Fogel,[20] John Hedges[21] and Noreen Tuross.[22]
Ralph Wyckoff
Wyckoff was a pioneer in X-ray crystallography and electron microscopy.[23] Using microscopic images, he demonstrated the variability and damage of collagen fibres in ancient bones and shells.[24] His research contributed to the understanding of protein diagenesis (degradation) in the late 1960s, and highlighted that ancient amino acid profiles alone might not be sufficient for protein identification.[25]
Margaret Jope and Peter Wesbroek
Jope and Wesbroek were leading experts in shell proteins and crystallisation.[26] Wesbroek later established Geobiochemistry laboratory at the University of Leiden, focusing on biomineralisation and how this process facilitated protein survival.[27] He also pioneered the use of antibodies for the study of ancient proteins in the 1970s and 1980s, utilising different immunological techniques such as Ouchterlony double immunodiffusion (interactions of antibodies and antigens in a gel).[28]
Peggy Ostrom
Ostrom championed the use of mass spectrometry since the 1990s.[29] She was the first to improve the sequence coverage of ancient proteins by combining different techniques such as peptide mass fingerprinting and liquid chromatography-tandem mass spectrometry (LC-MS/MS).[30]
The biochemistry of ancient proteins
Formation & incorporation
Understanding how ancient proteins are formed and incorporated into archaeological materials are essential in sampling, evaluating contamination and planning analyses.[1] Generally, for ancient proteins in proteinaceous tissues, notably, collagens in bones, keratins in wool, amelogenins in tooth enamel, and intracrystalline proteins in shells, they might be incorporated during the time of tissue formation.[31][32][33] However, the formation of proteinaceous tissues is often complex, dynamic and affected by various factors such pH, metals, ion concentration, diet plus other biological, chemical and physical parameters.[34] One of the most characterised phenomena is bone mineralisation, a process by which hydroxyapatite crystals are deposited within collagen fibres, forming a matrix.[35] Despite extensive research, bone scaffolding is still a challenge, and the role of non-collagenous proteins (a wide range of proteoglycans and other proteins) remains poorly understood.[36]
Another category is complex and potentially mineralised tissues, such as ancient human dental calculi and ceramic vessels. Dental calculi are defined as calcified biofilms, created and mediated by interactions between calcium phosphate ions and a wide range of oral microbial, human, and food proteins during episodic biomineralisation.[37][38] Similarly, the minerals of a ceramic matrix might interact with food proteins during food processing and cooking. This is best explained by calcite deposits adhering to the inside of archaeological ceramic vessels.[7] These protein-rich mineralised deposits might be formed during repeated cooking using hard water and subsequent scaling.[39]
Preservation
Organic (containing carbon) biomolecules like proteins are prone to degradation.[40] For example, experimental studies demonstrate that robust, fibrous and hydrophobic keratins such as feathers and woollen fabrics decay quickly at room temperature.[41][42] Indeed ancient proteins are exceptional, and they are often recovered from extreme burial contexts, especially dry and cold environments.[43][44] This is because the lack of water and low temperature may slow down hydrolysis, microbial attack and enzymatic activities.[31]
There are also proteins whose chemical and physical properties may enable their preservation in the long term. The best example is Type 1 collagen; it is one of the most abundant proteins in skin (80-85%) and bone (80-90%) extracellular matrices.[45] It is also mineralised, organised in a triple helix and stabilised by hydrogen bonding.[46] Type 1 collagen has been routinely extracted from ancient bones, leathers, and parchments; these characteristics may contribute to its stability over time.[47][48] Another common protein in the archaeological record is milk beta-lactoglobulin, often recovered from ancient dental calculi.[49] Beta-lactoglobulin is a small whey protein with a molecular mass of around 18400 Da (dalton).[50] It is resistant to heating and enzymatic degradation; structurally, it has a beta-barrel associated with binding to small hydrophobic molecules such as fatty acids, forming stable polymers.[51][52]
Given that proteins vary in abundance, size, hydrophobicity (water insolubility), structure, conformation (shape), function and stability, understanding protein preservation is challenging.[12] While there are common determinants of protein survival, including thermal history (temperature/time), burial conditions (pH/soil chemistry/water table) and protein properties (neighbouring amino acids/secondary structure/tertiary folding/proteome content), there is no clear answer and protein diagenesis is still an active research field.[1]
Structure & damage patterns
Generally, proteins have four levels of structural complexity: quaternary (multiple polypeptides, or subunits), tertiary (the 3D folding of a polypeptide), secondary (alpha helices/beta sheets/random coils) and primary structure (linear amino acid sequences linked by peptide bonds).[53] Ancient proteins are expected to lose their structural integrity over time, due to denaturation (protein unfolding) or other diagenetic processes.[54]
Ancient proteins also tend to be fragmented, damaged and altered. Proteins can be cleaved into small fragments over time, since hydrolysis (the addition of water) breaks peptide bonds (covalent bonds between two neighbouring alpha-amino acids).[55] In terms of post-translational modifications (changes occur after RNA translation), ancient proteins are often characterised by extensive damage such as oxidation (methionine), hydroxylation (proline), deamidation (glutamine/asparagine), citrullination (arginine), phosphorylation (serine/threonine/tyrosine), N-terminus glutamate to pyroglutamate and the addition of advanced glycation products to lysine or arginine.[56][12] Among these modifications, glutamine deamidation is one of the most time-dependent processes.[57] Glutamine deamidation is mostly a non-enzymatic process, by which glutamine is converted to glutamic acid (+0.98406 Da) via side-chain hydrolysis or the formation of a glutarimide ring.[58] It is a slow conversion with a long half-time, depending on adjacent amino acids, secondary structures, 3D folding, pH, temperature and other factors.[59] Bioinformatic tools are available to calculate bulk and site-specific deamidation rates of ancient proteins.[60] The structural manifestation of these chemical changes within ancient proteins was first documented using scanning electron microscopy (SEM). Type-1 collagen protein fibrils of a permafrost-preserved woolly mammoth (Yukon, Canada) were directly imaged and shown to retain their characteristic banding pattern. These were compared against type-1 collagen fibrils from a temperate Columbian mammoth specimen (Montana, U.S.A.). The Columbian mammoth collagen fibrils, unlike those of the permafrost-frozen woolly mammoth, had lost their banding, indicating substantial chemical degradation of the constituent peptide sequences. This also constitutes the first time that collagen banding, or the molecular structure for any ancient protein, has been directly imaged with scanning electron microscopy.[47]
Palaeoproteomics
Overview
Palaeoproteomics is a fast-developing field that combines archaeology, biology, chemistry and heritage studies. Comparable to its high-profile sister field, aDNA analysis, the extraction, identification and authentication of ancient proteins are challenging, since both ancient DNA and proteins tend to be ultrashort, highly fragmented, extensively damaged and chemically modified.[1][61]
However, ancient proteins are still one of the most informative biomolecules. Proteins tend to degrade more slowly than DNA, especially biomineralised proteins.[32][62] While ancient lipids can be used to differentiate between marine, plant and animal fats,[63] ancient protein data is high-resolution with taxon- and tissue-specificities.
To date, ancient peptide sequences have been successfully extracted and securely characterised from various archaeological remains, including a 3.8 Ma (million year) ostrich eggshell,[32] 1.77 Ma Homo erectus teeth,[64] a 0.16 Ma Denisovan jawbone[65] and several Neolithic (6000-5600 cal BC) pots.[7] Hence, palaeoproteomics has provided valuable insight into past evolutionary relationships, extinct species and societies.
Extraction
Generally, there are two approaches: a digestion-free, top-down method and bottom-up proteomics. Top-down proteomics is seldom used to analyse ancient proteins due to analytical and computational difficulties.[66] For bottom-up, or shotgun proteomics, ancient proteins are digested into peptides using enzymes, for example trypsin. Mineralised archaeological remains such as bones, teeth, shells, dental calculi and ceramics require an extra demineralisation step to release proteins from mineral matrices.[1] This is often achieved by using a weak acid (ethylenediaminetetraacetic acid, EDTA) or cold (4 °C) hydrochloric acid (HCl) to minimise chemical modifications that may introduced during extraction.[67]
To make ancient proteins soluble, heat, sonication, chaotropic agents (urea/guanidine hydrochloride, GnHCl), detergents or other buffers can be used.[1] Alkylation and reduction are often included for cysteine to disrupt disulfide bonds and avoid crosslinking.[68]
After demineralisation, protein solubilisation, alkylation and reduction, buffer exchange is needed to ensure that extracts are compatible with downstream analysis. Currently, there are three widely-used protocols for ancient proteins and gels (GASP),[69] filters (FASP)[70] and magnetic beads (SP3)[71] can be used for this purpose. Once buffer exchange is completed, extracts are incubated with digestion enzymes, then concentrated, purified and desalted.
For non-mineralised archaeological materials such as parchments, leathers and paintings, demineralisation is not necessary, and protocols can be changed depending on sample preservation and sampling size.[6]
Instrumentation and data analysis
Nowadays, palaeoproteomics is dominated by two mass spectrometry-based techniques: MALDI-ToF (matrix-assisted laser desorption/ionisation-time-of-flight) and LC-MS/MS. MALDI-ToF is used to determine the mass-to-charge (m/z) ratios of ions and their peak patterns.[72] Digested peptides are spotted on a MALDI plate, co-crystallise with a matrix (mainly α-cyano-4-hydroxycinnamic acid, CHCA); a laser excites and ionises the matrix, then its time to travel a vacuum tube is measured and converted to a spectrum of m/z ratios and intensities.[73]
Since only peak patterns, not entire amino acid sequences of digested peptides are characterised, peptide markers are needed for pattern matching and ancient protein identification.[72] In archaeological contexts, MALDI-ToF has been routinely used for bones and collagens in a field known as ZooMS (zooarchaeolgy by mass spectrometry).[2]
LC-MS/MS is another widely used approach. It is a powerful analytical technique to separate, sequence and quantify complex protein mixtures.[74] The first step in LC-MS/MS is liquid chromatography. Protein mixtures are separated in a liquid mobile phase using a stationary column.[75] How liquid analytes interact with a stationary phase depends on their size, charge, hydrophobicity and affinity.[76] These differences lead to distinct elution and retention time (when a component of a mixture exit a column). After chromatographic separation, protein components are ionised and introduced into mass spectrometers.[77] During a first mass scan (MS1), the m/z ratios of precursor ions are measured. Selected precursors are further fragmented and the m/z ratios of fragment ions are determined in a second mass scan (MS2). There are different fragmentation methods, for example, high-energy C-trap dissociation (HCD) and collision induced dissociation (CID), but b- and y-ions are frequently targeted.[78]
Search engines and software tools are often used to process ancient MS/MS data, including MaxQuant, Mascot and PEAKS.[79][80][81] Protein sequence data can be downloaded from public genebanks (UniProt/NCBI) and exported as FASTA files for sequencing algorithms.[13] Recently, open search engines such as MetaMorpheus, pFind and Fragpipe have received attention, because they make it possible to identify all modifications associated with peptide spectral matches (PSMs).[82][83][84]
De novo sequencing is also possible for the analysis of ancient MS/MS spectra. It is a sequencing technique that assembles amino acid sequences directly from spectra without reference databases.[85] Advances in deep learning also lead to the development of multiple pipelines such as DeNovoGUI, DeepNovo2 and Casanovo.[86][87][88] However, it may be challenging to evaluate the outputs of de novo sequences and optimisation may be required for ancient proteins to minimise false positives and overfitting.[1]
Palaeoproteomes
- Bones. Ancient bones are one of the most well-characterised and iconic palaeoproteomes. Ancient bone proteomes have been sequenced from hominins, humans, mammoths, moas and now extinct rhinoceros.[89][90][3][91][92][93] Fibrillar collagens are the most abundant proteins in modern bones; similarly, Type 1 and III collagens are also common in the archaeological record.[31] While modern bones contain about 10% of non-collagenous proteins (NCPs), various NCPs have been recorded, including osteocalcin, biglycan and lumican.[92] Generally, NCPs are excellent targets for studying evolution history, since they have higher turnover rates than bones.[31] Given the abundance of ancient bone proteomes, a bottom-up proteomic workflow known as SPIN (Species by Proteome INvestigation) is available for the high-throughput analysis of 150 million mammalian bones.[94]
- Teeth. Tooth enamel is one of the hardest and most mineralised tissues in the human body, since it is mainly composed of hydroxyapatite crystals.[95] While an enamel proteome is small, ancient amelogenins and other ameloblast-relevant proteins are often well-preserved in a mineralised, closed system.[31] Ancient enamel proteins are useful when aDNA or other proteins do not survive, and they have been analysed to understand extinct species and evolution.[96][97]
- Shells. Archaeological shells also contain rich palaoproteomes.[98] Like tooth enamel, They are more or less close systems that isolate proteins from water or other forces of degradation.[1] Strathiocalcin-1 and -2 are securely identified in 3.8 Ma ostrich eggshell samples at the site of Laetoli in Tazania.[32] These C-type lectins are associated with biomineralisation, and they are also found in extinct big bird shells collected from Australia.[4] Given the age of the ostrich eggshell, it was verified by a combination of six methods: analytical replication (same samples analysed in different labs), amino acid racemisation (D/L ratios), carry-over analysis (pre- and after-injection washes to evaluation the extent of carry-over in mass spectrometers), damage patterns (deamidation/oxidation/phosphorylation/amidation/decomposition) and aDNA studies.[32] These independent procedures ensure the authenticity of the oldest peptide sequences.
Other complex mixtures
- Ceramics & food crusts. Various ancient dietary proteins have been characterised from ceramics and associated food crusts (charred and calcite deposits on ceramic vessels).[7] Cow, sheep and goat milk beta-lactoglobulin proteins are predominant in this context, but there are also milk caseins (alpha-, beta- and kappa-casein), animal blood haemoglobins and a wide range of plant proteins (wheat glutenins, barley hordeins, legumins and other seed storage proteins).[99][100][101] The identification of these ancient foodstuffs may be used to understand how food was prepared, cooked and consumed in the past. It is also clear that archaeological ceramics and food crusts are complex mixtures that contain metaproteomes (multiple proteomes).[1]
Analytical challenges
While palaeoproteomics is a useful tool for a wide array of research questions, there are some analytical challenges that prevent the field from reaching its potential. The first issue is preservation. Mineral-binding seems to stabilise proteins, but this is a complex, dynamic process that has not been systematically investigated in different archaeological and burial contexts.[12][32]
Destructive sampling is another problem that can cause irreparable damage to archaeological materials. Although minimally-destructive or non-destructive sampling methods are being developed for parchments, bones, mummified tissues and leathers, it is unclear if they are suitable for other types of remains such as dental calculi, ceramics and food crusts.[102][103][104]
It is equally difficult to extract mineral-bound proteins due to their low abundance, extensive degradation, and often strong intermolecular interactions (hydrogen bonding, dispersion, ion-dipole and dipole-dipole interactions) with mineral matrice.[105] Ancient proteins also vary in preservation states, hydrophobicity, solubility and optimum pH values; methodological development is still required to maximise protein recovery.[106][107]
Ancient protein identification is still a challenge, because database search algorithms are not optimised for low-intensity and damaged ancient proteins, increasing the probabilities of false positive and false negatives.[12] There is also the issue of dark proteomes (unknown protein regions that cannot be sequenced); approximately 44-54% of proteins in eukaryotes such as animals and plants are dark.[108] Reference databases are also biassed towards model organisms such as yeasts and mouses,[109] and current sequence data may not cover all archaeological materials.
Lastly, while cytosine deamination (cytosine being converted to uracil over time that causes misreadings) has been widely used in the authentication of aDNA, there are no standardised procedures to authenticate ancient proteins.[61][110][111] This authentication issue is highlighted by the claim identification of 78 Ma Brachylophosaurus canadensis (hadrosaur) and 68 Ma Tyrannosaurus rex collagen peptides.[112][113] The lack of post-translational modifications and subsequent experimental studies demonstrate that these sequences may be derived from bacterial biofilms, the cross-contamination of control samples or modern laboratory procedures.[114]
Future directions
Despite significant analytical challenges, palaeoproteomics is constantly evolving and adopting new technology. Latest high-performance mass spectrometry, for example, TimsToF (trapped ion mobility-time-of-flight) in a DIA mode (data independent acquisition) may help with the separation, selection and resolution of ancient MS/MS data.[1] Novel extraction protocols such as DES (Deep Eutectic Solvent)-assisted procedures may increase the numbers and types of extracted palaeoproteomes.[115] Identification tools are also improving thanks to progress of bioinformatics, machine learning and artificial intelligence.[116]
Useful tools
Public depositories for raw data
Reference databases
Open search
See also
Further reading
Wikipedia pages
References
- Warinner C, Korzow Richter K, Collins MJ (August 2022). "Paleoproteomics". Chemical Reviews. 122 (16): 13401–13446. doi:10.1021/acs.chemrev.1c00703. PMC 9412968. PMID 35839101.
- Buckley M, Collins M, Thomas-Oates J, Wilson JC (December 2009). "Species identification by analysis of bone collagen using matrix-assisted laser desorption/ionisation time-of-flight mass spectrometry". Rapid Communications in Mass Spectrometry. 23 (23): 3843–3854. Bibcode:2009RCMS...23.3843B. doi:10.1002/rcm.4316. PMID 19899187.
- Cappellini E, Welker F, Pandolfi L, Ramos-Madrigal J, Samodova D, Rüther PL, et al. (October 2019). "Early Pleistocene enamel proteome from Dmanisi resolves Stephanorhinus phylogeny". Nature. 574 (7776): 103–107. Bibcode:2019Natur.574..103C. doi:10.1038/s41586-019-1555-y. PMC 6894936. PMID 31511700.
- Demarchi B, Stiller J, Grealy A, Mackie M, Deng Y, Gilbert T, et al. (October 2022). "Ancient proteins resolve controversy over the identity of Genyornis eggshell". Proceedings of the National Academy of Sciences of the United States of America. 119 (43): e2109326119. Bibcode:2022PNAS..11909326D. doi:10.1073/pnas.2109326119. PMC 9995833. PMID 35609205. S2CID 249045755.
- Elnaggar A, Osama A, Anwar AM, Ezzeldin S, Abou Elhassan S, Ebeid H, et al. (2022-11-09). "Paleoproteomic profiling for identification of animal skin species in ancient Egyptian archaeological leather using liquid chromatography coupled with tandem mass spectrometry (LC-MS/MS)". Heritage Science. 10 (1): 182. doi:10.1186/s40494-022-00816-0. ISSN 2050-7445. S2CID 253399828.
- Fiddyment S, Teasdale MD, Vnouček J, Lévêque É, Binois A, Collins MJ (2019-06-07). "So you want to do biocodicology? A field guide to the biological analysis of parchment". Heritage Science. 7 (1): 35. doi:10.1186/s40494-019-0278-6. ISSN 2050-7445. S2CID 195245888.
- Hendy J, Colonese AC, Franz I, Fernandes R, Fischer R, Orton D, et al. (October 2018). "Ancient proteins from ceramic vessels at Çatalhöyük West reveal the hidden cuisine of early farmers". Nature Communications. 9 (1): 4064. Bibcode:2018NatCo...9.4064H. doi:10.1038/s41467-018-06335-6. PMC 6170438. PMID 30283003.
- Dallongeville S, Garnier N, Rolando C, Tokarski C (January 2016). "Proteins in Art, Archaeology, and Paleontology: From Detection to Identification". Chemical Reviews. 116 (1): 2–79. doi:10.1021/acs.chemrev.5b00037. PMID 26709533.
- Maixner F, Turaev D, Cazenave-Gassiot A, Janko M, Krause-Kyora B, Hoopmann MR, et al. (July 2018). "The Iceman's Last Meal Consisted of Fat, Wild Meat, and Cereals". Current Biology. 28 (14): 2348–2355.e9. doi:10.1016/j.cub.2018.05.067. PMC 6065529. PMID 30017480.
- Cappellini E, Prohaska A, Racimo F, Welker F, Pedersen MW, Allentoft ME, et al. (June 2018). "Ancient Biomolecules and Evolutionary Inference". Annual Review of Biochemistry. 87 (1): 1029–1060. doi:10.1146/annurev-biochem-062917-012002. hdl:21.11116/0000-0001-DF45-7. PMID 29709200. S2CID 14004952.
- Hendy J (January 2021). "Ancient protein analysis in archaeology". Science Advances. 7 (3). Bibcode:2021SciA....7.9314H. doi:10.1126/sciadv.abb9314. PMC 7810370. PMID 33523896.
- Hendy J, van Doorn N, Collins M (2020). "Proteomics". In Britton K, Richards MP (eds.). Archaeological Science: An Introduction. Cambridge: Cambridge University Press. pp. 35–69. doi:10.1017/9781139013826.003. ISBN 978-0-521-19522-5. S2CID 241941528.
- Hendy J, Welker F, Demarchi B, Speller C, Warinner C, Collins MJ (May 2018). "A guide to ancient protein studies" (PDF). Nature Ecology & Evolution. 2 (5): 791–799. doi:10.1038/s41559-018-0510-x. PMID 29581591. S2CID 256704765.
- Yilmaz M, Fondrie WE, Bittremieux W, Nelson R, Ananth V, Oh S, Noble WS (2023-01-04). "Sequence-to-sequence translation from mass spectra to peptides with a transformer model". bioRxiv: 2023.01.03.522621. doi:10.1101/2023.01.03.522621. S2CID 255441838.
- Chi H, Liu C, Yang H, Zeng WF, Wu L, Zhou WJ, et al. (October 2018). "Comprehensive identification of peptides in tandem mass spectra using an efficient open search engine". Nature Biotechnology. 36 (11): 1059–1061. doi:10.1038/nbt.4236. PMID 30295672. S2CID 52930101.
- Abelson PH (1 July 1956). "Paleobiochemistry". Scientific American. Retrieved 2023-02-19.
- "National Academy of Sciences: Abstracts of Papers Presented at the Annual Meeting April 26-28, 1954, Washington, D.C". Science. 119 (3096): 576–588. April 1954. doi:10.1126/science.119.3096.576. PMID 17777440.
- Wehmiller J, Hare PE (September 1971). "Racemization of amino acids in marine sediments". Science. 173 (4000): 907–911. Bibcode:1971Sci...173..907W. doi:10.1126/science.173.4000.907. PMID 17751312. S2CID 17286901.
- Abelson PH, Hoering TC. Carbon Isotope Fractionation in Formation of Amino Acids by Photosynthetic Organisms. OCLC 678738249.
- Hare PE, Fogel ML, Stafford Jr TW, Mitchell AD, Hoering TC (May 1991). "The isotopic composition of carbon and nitrogen in individual amino acids isolated from modern and fossil proteins". Journal of Archaeological Science. 18 (3): 277–292. Bibcode:1991JArSc..18..277E. doi:10.1016/0305-4403(91)90066-X. ISSN 0305-4403.
- Hedges JI, Hare PE (February 1987). "Amino acid adsorption by clay minerals in distilled water". Geochimica et Cosmochimica Acta. 51 (2): 255–259. Bibcode:1987GeCoA..51..255H. doi:10.1016/0016-7037(87)90237-7. ISSN 0016-7037.
- Tuross N, Fogel ML, Hare PE (1988-04-01). "Variability in the preservation of the isotopic composition of collagen from fossil bone". Geochimica et Cosmochimica Acta. 52 (4): 929–935. Bibcode:1988GeCoA..52..929T. doi:10.1016/0016-7037(88)90364-X. ISSN 0016-7037.
- Wyckoff RW, Mccaughey WF, Doberenz AR (November 1964). "The amino acid composition of proteins from pleistocene bones☆". Biochimica et Biophysica Acta (BBA) - General Subjects. 93 (2): 374–377. doi:10.1016/0304-4165(64)90387-3. PMID 14251315.
- Akiyama M, Wyckoff RW. The Total Amino Acid Content of Fossil Pecten Shells*. OCLC 678746870.
- Wyckoff RW, Doberenz AR (February 1965). "The Electron Microscopy of Rancho La Brea Bone". Proceedings of the National Academy of Sciences of the United States of America. 53 (2): 230–233. Bibcode:1965PNAS...53..230W. doi:10.1073/pnas.53.2.230. PMC 219495. PMID 14294054.
- Jope M (1967-02-01). "The protein of brachiopod shell—I. Amino acid composition and implied protein taxonomy". Comparative Biochemistry and Physiology. 20 (2): 593–600. doi:10.1016/0010-406X(67)90271-X. ISSN 0010-406X.
- Westbroek P (1992). Life as a geological force : dynamics of the earth. W.W. Norton. ISBN 0-393-30817-0. OCLC 26700519.
- de Jong EW, Westbroek P, Westbroek JW, Bruning JW (November 1974). "Preservation of antigenic properties of macromolecules over 70 Myr". Nature. 252 (5478): 63–64. Bibcode:1974Natur.252...63D. doi:10.1038/252063a0. PMID 4139661. S2CID 4286786.
- Ostrom PH, Macko SA, Engel MH, Silfer JA, Russell D (January 1990). "Geochemical characterization of high molecular weight material isolated from late cretaceous fossils". Organic Geochemistry. Proceedings of the 14th International Meeting on Organic Geochemistry. 16 (4): 1139–1144. Bibcode:1990OrGeo..16.1139O. doi:10.1016/0146-6380(90)90149-T. ISSN 0146-6380.
- Ostrom PH, Gandhi H, Strahler JR, Walker AK, Andrews PC, Leykam J, et al. (2006-04-15). "Unraveling the sequence and structure of the protein osteocalcin from a 42ka fossil horse". Geochimica et Cosmochimica Acta. 70 (8): 2034–2044. Bibcode:2006GeCoA..70.2034O. doi:10.1016/j.gca.2006.01.004. ISSN 0016-7037.
- Kendall C, Eriksen AM, Kontopoulos I, Collins MJ, Turner-Walker G (February 2018). "Diagenesis of archaeological bone and tooth" (PDF). Palaeogeography, Palaeoclimatology, Palaeoecology. 491: 21–37. Bibcode:2018PPP...491...21K. doi:10.1016/j.palaeo.2017.11.041. ISSN 0031-0182.
- Demarchi B, Hall S, Roncal-Herrero T, Freeman CL, Woolley J, Crisp MK, et al. (September 2016). Krause J (ed.). "Protein sequences bound to mineral surfaces persist into deep time". eLife. 5: e17092. doi:10.7554/eLife.17092. PMC 5039028. PMID 27668515.
- Brandt LØ, Taurozzi AJ, Mackie M, Sinding MS, Vieira FG, Schmidt AL, et al. (2022-07-27). "Palaeoproteomics identifies beaver fur in Danish high-status Viking Age burials - direct evidence of fur trade". PLOS ONE. 17 (7): e0270040. Bibcode:2022PLoSO..1770040B. doi:10.1371/journal.pone.0270040. PMC 9328512. PMID 35895633.
- Sharma V, Srinivasan A, Nikolajeff F, Kumar S (January 2021). "Biomineralization process in hard tissues: The interaction complexity within protein and inorganic counterparts". Acta Biomaterialia. Biomineralization: From Cells to Biomaterials. 120: 20–37. doi:10.1016/j.actbio.2020.04.049. PMID 32413577. S2CID 218657975.
- Burr DB (February 2019). "Chapter 1 - Bone Morphology and Organization". In Burr DB, Allen MR (eds.). Basic and Applied Bone Biology (Second ed.). Academic Press. pp. 3–26. ISBN 978-0-12-813259-3.
- Carvalho MS, Cabral JM, da Silva CL, Vashishth D (March 2021). "Bone Matrix Non-Collagenous Proteins in Tissue Engineering: Creating New Bone by Mimicking the Extracellular Matrix". Polymers. 13 (7): 1095. doi:10.3390/polym13071095. PMC 8036283. PMID 33808184.
- Warinner C, Speller C, Collins MJ (January 2015). "A new era in palaeomicrobiology: prospects for ancient dental calculus as a long-term record of the human oral microbiome". Philosophical Transactions of the Royal Society of London. Series B, Biological Sciences. 370 (1660): 20130376. doi:10.1098/rstb.2013.0376. PMC 4275884. PMID 25487328.
- Radini A, Nikita E, Buckley S, Copeland L, Hardy K (January 2017). "Beyond food: The multiple pathways for inclusion of materials into ancient dental calculus". American Journal of Physical Anthropology. 162 (Suppl 63): 71–83. doi:10.1002/ajpa.23147. PMID 28105717.
- Lin L, Jiang W, Xu X, Xu P (June 2020). "A critical review of the application of electromagnetic fields for scaling control in water systems: mechanisms, characterization, and operation". npj Clean Water. 3 (1): 1–19. doi:10.1038/s41545-020-0071-9. ISSN 2059-7037. S2CID 219156792.
- Brown T, Brown K (2011-02-04). Biomolecular Archaeology: An Introduction (1st ed.). Wiley. doi:10.1002/9781444392449. ISBN 978-1-4051-7960-7.
- Saitta ET, Rogers C, Brooker RA, Abbott GD, Kumar S, O'Reilly SS, et al. (2017). Smith A (ed.). "Low fossilization potential of keratin protein revealed by experimental taphonomy". Palaeontology. 60 (4): 547–556. Bibcode:2017Palgy..60..547S. doi:10.1111/pala.12299. S2CID 90509524.
- Solazzo C, Dyer JM, Clerens S, Plowman J, Peacock EE, Collins MJ (May 2013). "Proteomic evaluation of the biodegradation of wool fabrics in experimental burials". International Biodeterioration & Biodegradation. 80: 48–59. doi:10.1016/j.ibiod.2012.11.013. ISSN 0964-8305.
- Yang Y, Shevchenko A, Knaust A, Abuduresule I, Li W, Hu X, et al. (May 2014). "Proteomics evidence for kefir dairy in Early Bronze Age China". Journal of Archaeological Science. 45: 178–186. Bibcode:2014JArSc..45..178Y. doi:10.1016/j.jas.2014.02.005. ISSN 0305-4403.
- Hollemeyer K, Altmeyer W, Heinzle E, Pitra C (September 2008). "Species identification of Oetzi's clothing with matrix-assisted laser desorption/ionization time-of-flight mass spectrometry based on peptide pattern similarities of hair digests". Rapid Communications in Mass Spectrometry. 22 (18): 2751–2767. Bibcode:2008RCMS...22.2751H. doi:10.1002/rcm.3679. PMID 18720427.
- Amirrah IN, Lokanathan Y, Zulkiflee I, Wee MF, Motta A, Fauzi MB (September 2022). "A Comprehensive Review on Collagen Type I Development of Biomaterials for Tissue Engineering: From Biosynthesis to Bioscaffold". Biomedicines. 10 (9): 2307. doi:10.3390/biomedicines10092307. PMC 9496548. PMID 36140407.
- BONE CELL BIOMECHANICS, MECHANOBIOLOGY AND BONE DISEASES. [S.l.]: ELSEVIER ACADEMIC PRESS. 2023. ISBN 978-0-323-96123-3. OCLC 1336986913.
- Anderson L (December 2022). "Biomolecular histology as a novel proxy for ancient DNA and protein sequence preservation". Ecology and Evolution. 12 (12): e9518. doi:10.1002/ece3.9518. PMC 9743065. PMID 36518622.
- Martínez Cortizas A, López-Costas O (October 2020). "Linking structural and compositional changes in archaeological human bone collagen: an FTIR-ATR approach". Scientific Reports. 10 (1): 17888. Bibcode:2020NatSR..1017888M. doi:10.1038/s41598-020-74993-y. PMC 7578014. PMID 33087827.
- Hendy J, Warinner C, Bouwman A, Collins MJ, Fiddyment S, Fischer R, et al. (July 2018). "Proteomic evidence of dietary sources in ancient dental calculus". Proceedings. Biological Sciences. 285 (1883): 20180977. doi:10.1098/rspb.2018.0977. PMC 6083251. PMID 30051838.
- Liu HC, Chen WL, Mao SJ (February 2007). "Antioxidant nature of bovine milk beta-lactoglobulin". Journal of Dairy Science. 90 (2): 547–555. doi:10.3168/jds.s0022-0302(07)71538-2. PMID 17235131.
- Brownlow S, Morais Cabral JH, Cooper R, Flower DR, Yewdall SJ, Polikarpov I, et al. (April 1997). "Bovine beta-lactoglobulin at 1.8 A resolution--still an enigmatic lipocalin". Structure. 5 (4): 481–495. doi:10.1016/S0969-2126(97)00205-0. PMID 9115437.
- Le Maux S, Bouhallab S, Giblin L, Brodkorb A, Croguennec T (2014). "Bovine β-lactoglobulin/fatty acid complexes: binding, structural, and biological properties". Dairy Science & Technology. 94 (5): 409–426. doi:10.1007/s13594-014-0160-y. PMC 4121524. PMID 25110551.
- Clayden J (2001). Organic chemistry. Oxford: Oxford University Press. ISBN 0-19-850347-4. OCLC 43338068.
- Pavelka J, Smejda L, Hynek R, Kuckova SH (September 2016). "Immunological detection of denatured proteins as a method for rapid identification of food residues on archaeological pottery". Journal of Archaeological Science. 73: 25–35. Bibcode:2016JArSc..73...25P. doi:10.1016/j.jas.2016.07.004. ISSN 0305-4403.
- Fonseca B, Freeman CL, Collins MJ (September 2022). "Conformational analysis and water dynamics: a molecular dynamics study on the survival of a β-lactoglobulin peptide in the archaeological record". Chemical Physics. 561: 111602. Bibcode:2022CP....56111602F. doi:10.1016/j.chemphys.2022.111602. ISSN 0301-0104. S2CID 249454628.
- Müller MM (January 2018). "Post-Translational Modifications of Protein Backbones: Unique Functions, Mechanisms, and Challenges". Biochemistry. 57 (2): 177–185. doi:10.1021/acs.biochem.7b00861. PMC 5770884. PMID 29064683.
- Boudier-Lemosquet A, Mahler A, Bobo C, Dufossée M, Priault M (April 2022). "Introducing protein deamidation: Landmark discoveries, societal outreach, and tentative priming workflow to address deamidation". Methods. Intrinsic protein instability : deamidation. 200: 3–14. doi:10.1016/j.ymeth.2021.11.012. PMID 34843979. S2CID 244699364.
- Riggs DL, Silzel JW, Lyon YA, Kang AS, Julian RR (October 2019). "Analysis of Glutamine Deamidation: Products, Pathways, and Kinetics". Analytical Chemistry. 91 (20): 13032–13038. doi:10.1021/acs.analchem.9b03127. PMC 8805438. PMID 31498611.
- Robinson NE, Robinson AB (January 2001). "Molecular clocks". Proceedings of the National Academy of Sciences of the United States of America. 98 (3): 944–949. Bibcode:2001PNAS...98..944R. doi:10.1073/pnas.98.3.944. PMC 14689. PMID 11158575.
- Ramsøe A, Crispin M, Mackie M, McGrath K, Fischer R, Demarchi B, et al. (April 2021). "Assessing the degradation of ancient milk proteins through site-specific deamidation patterns". Scientific Reports. 11 (1): 7795. Bibcode:2021NatSR..11.7795R. doi:10.1038/s41598-021-87125-x. PMC 8032661. PMID 33833277.
- Orlando L, Allaby R, Skoglund P, Der Sarkissian C, Stockhammer PW, Ávila-Arcos MC, et al. (2021-02-11). "Ancient DNA analysis". Nature Reviews Methods Primers. 1 (1): 1–26. doi:10.1038/s43586-020-00011-0. ISSN 2662-8449. S2CID 233911592.
- Allentoft ME, Collins M, Harker D, Haile J, Oskam CL, Hale ML, et al. (December 2012). "The half-life of DNA in bone: measuring decay kinetics in 158 dated fossils". Proceedings. Biological Sciences. 279 (1748): 4724–4733. doi:10.1098/rspb.2012.1745. PMC 3497090. PMID 23055061.
- Craig OE, Saul H, Spiteri C (2020). "Residue Analysis". In Britton K, Richards MP (eds.). Archaeological Science: An Introduction. Cambridge: Cambridge University Press. pp. 70–98. doi:10.1017/9781139013826.004. ISBN 978-0-521-19522-5. S2CID 241568727. Retrieved 2023-02-21.
- Welker F, Ramos-Madrigal J, Gutenbrunner P, Mackie M, Tiwary S, Rakownikow Jersie-Christensen R, et al. (April 2020). "The dental proteome of Homo antecessor". Nature. 580 (7802): 235–238. Bibcode:2020Natur.580..235W. doi:10.1038/s41586-020-2153-8. PMC 7582224. PMID 32269345.
- Chen F, Welker F, Shen CC, Bailey SE, Bergmann I, Davis S, et al. (May 2019). "A late Middle Pleistocene Denisovan mandible from the Tibetan Plateau" (PDF). Nature. 569 (7756): 409–412. Bibcode:2019Natur.569..409C. doi:10.1038/s41586-019-1139-x. PMID 31043746. S2CID 256768558.
- Cleland TP, Schroeter ER, Colleary C (January 2021). "Diagenetiforms: A new term to explain protein changes as a result of diagenesis in paleoproteomics". Journal of Proteomics. 230: 103992. doi:10.1016/j.jprot.2020.103992. PMID 32992016. S2CID 222168890.
- Palmer KS, Makarewicz CA, Tishkin AA, Tur SS, Chunag A, Diimajav E, et al. (March 2021). "Comparing the Use of Magnetic Beads with Ultrafiltration for Ancient Dental Calculus Proteomics" (PDF). Journal of Proteome Research. 20 (3): 1689–1704. doi:10.1021/acs.jproteome.0c00862. PMID 33596076. S2CID 231953980.
- Sechi S, Chait BT (December 1998). "Modification of cysteine residues by alkylation. A tool in peptide mapping and protein identification". Analytical Chemistry. 70 (24): 5150–5158. doi:10.1021/ac9806005. PMID 9868912.
- Fischer R, Kessler BM (April 2015). "Gel-aided sample preparation (GASP)--a simplified method for gel-assisted proteomic sample generation from protein extracts and intact cells". Proteomics. 15 (7): 1224–1229. doi:10.1002/pmic.201400436. PMC 4409837. PMID 25515006.
- Wiśniewski RJ (2018). "Filter-Aided Sample Preparation for Proteome Analysis". In Becher D (ed.). Microbial Proteomics. Methods in Molecular Biology. Vol. 1841. New York, NY: Springer New York. pp. 3–10. doi:10.1007/978-1-4939-8695-8_1. ISBN 978-1-4939-8693-4. PMID 30259475.
- Hughes CS, Moggridge S, Müller T, Sorensen PH, Morin GB, Krijgsveld J (January 2019). "Single-pot, solid-phase-enhanced sample preparation for proteomics experiments". Nature Protocols. 14 (1): 68–85. doi:10.1038/s41596-018-0082-x. PMID 30464214. S2CID 256838729.
- Richter KK, Codlin MC, Seabrook M, Warinner C (May 2022). "A primer for ZooMS applications in archaeology". Proceedings of the National Academy of Sciences of the United States of America. 119 (20): e2109323119. Bibcode:2022PNAS..11909323R. doi:10.1073/pnas.2109323119. PMC 9171758. PMID 35537051.
- Hosseini S, Martinez-Chapa SO (2017). "Principles and Mechanism of MALDI-ToF-MS Analysis". In Hosseini S, Martinez-Chapa SO (eds.). Fundamentals of MALDI-ToF-MS Analysis. pp. 1–19. doi:10.1007/978-981-10-2356-9_1. ISBN 978-981-10-2356-9. Retrieved 2023-02-21.
{{cite book}}:|work=ignored (help) - Patterson SD, Aebersold RH (March 2003). "Proteomics: the first decade and beyond". Nature Genetics. 33 (3): 311–323. doi:10.1038/ng1106. PMID 12610541. S2CID 9800076.
- Akash MS, Rehman K (2020). "High Performance Liquid Chromatography". In Akash MS, Rehman K (eds.). Essentials of Pharmaceutical Analysis. Singapore: Springer Nature. pp. 175–184. doi:10.1007/978-981-15-1547-7_14. ISBN 978-981-15-1547-7. S2CID 212917192. Retrieved 2023-02-21.
- Niessen WM (2006). Liquid chromatography--mass spectrometry (3rd ed.). Boca Raton. ISBN 0-429-11680-2. OCLC 1329091536.
{{cite book}}: CS1 maint: location missing publisher (link) - Seger C, Salzmann L (August 2020). "After another decade: LC-MS/MS became routine in clinical diagnostics". Clinical Biochemistry. Advancement and Applications of Mass Spectrometry in Laboratory Medicine. 82: 2–11. doi:10.1016/j.clinbiochem.2020.03.004. PMID 32188572. S2CID 213186669.
- Lindon JC, Tranter GE, Koppenaal DW (2016). Encyclopedia of spectroscopy and spectrometry (3rd ed.). Kidlington, Oxford, United Kingdom. ISBN 978-0-12-803225-1. OCLC 960910529.
{{cite book}}: CS1 maint: location missing publisher (link) - Tyanova S, Temu T, Cox J (December 2016). "The MaxQuant computational platform for mass spectrometry-based shotgun proteomics". Nature Protocols. 11 (12): 2301–2319. doi:10.1038/nprot.2016.136. PMID 27809316. S2CID 21258415.
- Hirosawa M, Hoshida M, Ishikawa M, Toya T (April 1993). "MASCOT: multiple alignment system for protein sequences based on three-way dynamic programming". Computer Applications in the Biosciences. 9 (2): 161–167. doi:10.1093/bioinformatics/9.2.161. PMID 8481818.
- Ma B, Zhang K, Hendrie C, Liang C, Li M, Doherty-Kirby A, Lajoie G (2003-10-30). "PEAKS: powerful software for peptide de novo sequencing by tandem mass spectrometry". Rapid Communications in Mass Spectrometry. 17 (20): 2337–2342. Bibcode:2003RCMS...17.2337M. doi:10.1002/rcm.1196. PMID 14558135.
- Solntsev SK, Shortreed MR, Frey BL, Smith LM (May 2018). "Enhanced Global Post-translational Modification Discovery with MetaMorpheus". Journal of Proteome Research. 17 (5): 1844–1851. doi:10.1021/acs.jproteome.7b00873. PMID 29578715.
- Sun J, Shi J, Wang Y, Wu S, Zhao L, Li Y, et al. (December 2019). "Open-pFind Enhances the Identification of Missing Proteins from Human Testis Tissue". Journal of Proteome Research. 18 (12): 4189–4196. doi:10.1021/acs.jproteome.9b00376. PMID 31657219. S2CID 204947980.
- Geiszler DJ, Kong AT, Avtonomov DM, Yu F, Leprevost FD, Nesvizhskii AI (2021-01-01). "PTM-Shepherd: Analysis and Summarization of Post-Translational and Chemical Modifications From Open Search Results". Molecular & Cellular Proteomics. 20: 100018. doi:10.1074/mcp.TIR120.002216. PMC 7950090. PMID 33568339.
- Tran NH, Zhang X, Xin L, Shan B, Li M (August 2017). "De novo peptide sequencing by deep learning". Proceedings of the National Academy of Sciences of the United States of America. 114 (31): 8247–8252. Bibcode:2017PNAS..114.8247T. doi:10.1073/pnas.1705691114. PMC 5547637. PMID 28720701.
- Muth T, Weilnböck L, Rapp E, Huber CG, Martens L, Vaudel M, Barsnes H (February 2014). "DeNovoGUI: an open source graphical user interface for de novo sequencing of tandem mass spectra". Journal of Proteome Research. 13 (2): 1143–1146. doi:10.1021/pr4008078. PMC 3923451. PMID 24295440.
- Qiao R, Tran NH, Xin L, Chen X, Li M, Shan B, Ghodsi A (2021). "Computationally instrument-resolution-independent de novo peptide sequencing for high-resolution devices". Nature Machine Intelligence. 3 (5): 420–425. doi:10.1038/s42256-021-00304-3. ISSN 2522-5839. S2CID 233670194.
- Yilmaz M, Fondrie W, Bittremieux W, Oh S, Noble WS (2022-06-28). "De novo mass spectrometry peptide sequencing with a transformer model". Proceedings of the 39th International Conference on Machine Learning. PMLR: 25514–25522.
- Bona A, Papai Z, Maasz G, Toth GA, Jambor E, Schmidt J, et al. (2014-01-27). "Mass spectrometric identification of ancient proteins as potential molecular biomarkers for a 2000-year-old osteogenic sarcoma". PLOS ONE. 9 (1): e87215. Bibcode:2014PLoSO...987215B. doi:10.1371/journal.pone.0087215. PMC 3903643. PMID 24475253.
- Cappellini E, Jensen LJ, Szklarczyk D, Ginolhac A, da Fonseca RA, Stafford TW, et al. (February 2012). "Proteomic analysis of a pleistocene mammoth femur reveals more than one hundred ancient bone proteins". Journal of Proteome Research. 11 (2): 917–926. doi:10.1021/pr200721u. PMID 22103443.
- Cleland TP, Schroeter ER, Schweitzer MH (June 2015). "Biologically and diagenetically derived peptide modifications in moa collagens". Proceedings. Biological Sciences. 282 (1808): 20150015. doi:10.1098/rspb.2015.0015. PMC 4455796. PMID 25972464.
- Welker F, Hajdinjak M, Talamo S, Jaouen K, Dannemann M, David F, et al. (October 2016). "Palaeoproteomic evidence identifies archaic hominins associated with the Châtelperronian at the Grotte du Renne". Proceedings of the National Academy of Sciences of the United States of America. 113 (40): 11162–11167. Bibcode:2016PNAS..11311162W. doi:10.1073/pnas.1605834113. PMC 5056053. PMID 27638212.
- Hublin JJ, Sirakov N, Aldeias V, Bailey S, Bard E, Delvigne V, et al. (May 2020). "Initial Upper Palaeolithic Homo sapiens from Bacho Kiro Cave, Bulgaria" (PDF). Nature. 581 (7808): 299–302. Bibcode:2020Natur.581..299H. doi:10.1038/s41586-020-2259-z. PMID 32433609. S2CID 218592678.
- Rüther PL, Husic IM, Bangsgaard P, Gregersen KM, Pantmann P, Carvalho M, et al. (May 2022). "SPIN enables high throughput species identification of archaeological bone by proteomics". Nature Communications. 13 (1): 2458. Bibcode:2022NatCo..13.2458R. doi:10.1038/s41467-022-30097-x. PMC 9072323. PMID 35513387.
- Bai Y, Yu Z, Ackerman L, Zhang Y, Bonde J, Li W, et al. (August 2020). "Protein nanoribbons template enamel mineralization". Proceedings of the National Academy of Sciences of the United States of America. 117 (32): 19201–19208. Bibcode:2020PNAS..11719201B. doi:10.1073/pnas.2007838117. PMC 7431033. PMID 32737162.
- Welker F, Ramos-Madrigal J, Kuhlwilm M, Liao W, Gutenbrunner P, de Manuel M, et al. (December 2019). "Enamel proteome shows that Gigantopithecus was an early diverging pongine". Nature. 576 (7786): 262–265. Bibcode:2019Natur.576..262W. doi:10.1038/s41586-019-1728-8. PMC 6908745. PMID 31723270.
- Nogueira FC, Neves LX, Pessoa-Lima C, Langer MC, Domont GB, Line SR, et al. (May 2021). "Ancient enamel peptides recovered from the South American Pleistocene species Notiomastodon platensis and Myocastor cf. coypus". Journal of Proteomics. 240: 104187. doi:10.1016/j.jprot.2021.104187. PMID 33757878. S2CID 232337986.
- Sakalauskaite J, Marin F, Pergolizzi B, Demarchi B (September 2020). "Shell palaeoproteomics: First application of peptide mass fingerprinting for the rapid identification of mollusc shells in archaeology". Journal of Proteomics. 227: 103920. doi:10.1016/j.jprot.2020.103920. PMID 32712371. S2CID 220798638.
- Chowdhury MP, Campbell S, Buckley M (August 2021). "Proteomic analysis of archaeological ceramics from Tell Khaiber, southern Iraq". Journal of Archaeological Science. 132: 105414. Bibcode:2021JArSc.132j5414C. doi:10.1016/j.jas.2021.105414. ISSN 0305-4403. S2CID 236243360.
- Siano F, Picariello G, Caruso T, Esposito S, Rescigno C, Addeo F, Vasca E (May 2022). "Proteomics and Integrated Techniques to Characterize Organic Residues in Funerary Findings from Italic Populations of the First Millennium BC". Journal of Proteome Research. 21 (5): 1330–1339. doi:10.1021/acs.jproteome.2c00093. PMID 35347988. S2CID 247776564.
- Tanasi D, Cucina A, Cunsolo V, Saletti R, Di Francesco A, Greco E, Foti S (February 2021). "Paleoproteomic profiling of organic residues on prehistoric pottery from Malta". Amino Acids. 53 (2): 295–312. doi:10.1007/s00726-021-02946-4. PMC 7910365. PMID 33582869.
- McGrath K, Rowsell K, Gates St-Pierre C, Tedder A, Foody G, Roberts C, et al. (July 2019). "Identifying Archaeological Bone via Non-Destructive ZooMS and the Materiality of Symbolic Expression: Examples from Iroquoian Bone Points". Scientific Reports. 9 (1): 11027. Bibcode:2019NatSR...911027M. doi:10.1038/s41598-019-47299-x. PMC 6667708. PMID 31363122.
- Demarchi B, Boano R, Ceron A, Dal Bello F, Favero-Longo SE, Fiddyment S, et al. (July 2020). "Never boring: Non-invasive palaeoproteomics of mummified human skin". Journal of Archaeological Science. 119: 105145. Bibcode:2020JArSc.119j5145D. doi:10.1016/j.jas.2020.105145. ISSN 0305-4403. S2CID 219750410.
- Martisius NL, Welker F, Dogandžić T, Grote MN, Rendu W, Sinet-Mathiot V, et al. (May 2020). "Non-destructive ZooMS identification reveals strategic bone tool raw material selection by Neandertals". Scientific Reports. 10 (1): 7746. Bibcode:2020NatSR..10.7746M. doi:10.1038/s41598-020-64358-w. PMC 7210944. PMID 32385291.
- Craig OE, Collins MJ (March 2000). "An improved method for the immunological detection of mineral bound protein using hydrofluoric acid and direct capture". Journal of Immunological Methods. 236 (1–2): 89–97. doi:10.1016/S0022-1759(99)00242-2. PMID 10699582.
- Procopio N, Buckley M (February 2017). "Minimizing Laboratory-Induced Decay in Bone Proteomics". Journal of Proteome Research. 16 (2): 447–458. doi:10.1021/acs.jproteome.6b00564. PMID 28152590.
- Cleland TP (November 2018). "Human Bone Paleoproteomics Utilizing the Single-Pot, Solid-Phase-Enhanced Sample Preparation Method to Maximize Detected Proteins and Reduce Humics". Journal of Proteome Research. 17 (11): 3976–3983. doi:10.1021/acs.jproteome.8b00637. PMID 30336043. S2CID 53018221.
- Perdigão N, Heinrich J, Stolte C, Sabir KS, Buckley MJ, Tabor B, et al. (December 2015). "Unexpected features of the dark proteome". Proceedings of the National Academy of Sciences of the United States of America. 112 (52): 15898–15903. Bibcode:2015PNAS..11215898P. doi:10.1073/pnas.1508380112. PMC 4702990. PMID 26578815.
- Ankeny RA, Leonelli S (2020). "Model Organisms". Elements in the Philosophy of Biology. doi:10.1017/9781108593014. ISBN 9781108593014.
- Schroeter ER, Cleland TP (January 2016). "Glutamine deamidation: an indicator of antiquity, or preservational quality?". Rapid Communications in Mass Spectrometry. 30 (2): 251–255. Bibcode:2016RCMS...30..251S. doi:10.1002/rcm.7445. PMID 26689157.
- Brown S, Kozlikin M, Shunkov M, Derevianko A, Higham T, Douka K, Richter KK (September 2021). "Examining collagen preservation through glutamine deamidation at Denisova Cave". Journal of Archaeological Science. 133: 105454. Bibcode:2021JArSc.133j5454B. doi:10.1016/j.jas.2021.105454. ISSN 0305-4403.
- Asara JM, Schweitzer MH, Freimark LM, Phillips M, Cantley LC (April 2007). "Protein sequences from mastodon and Tyrannosaurus rex revealed by mass spectrometry". Science. 316 (5822): 280–285. Bibcode:2007Sci...316..280A. doi:10.1126/science.1137614. PMID 17431180. S2CID 85299309.
- Schweitzer MH, Zheng W, Organ CL, Avci R, Suo Z, Freimark LM, et al. (May 2009). "Biomolecular characterization and protein sequences of the Campanian hadrosaur B. canadensis". Science. 324 (5927): 626–631. Bibcode:2009Sci...324..626S. doi:10.1126/science.1165069. PMID 19407199. S2CID 5358680.
- Buckley M, Warwood S, van Dongen B, Kitchener AC, Manning PL (May 2017). "A fossil protein chimera; difficulties in discriminating dinosaur peptide sequences from modern cross-contamination". Proceedings. Biological Sciences. 284 (1855): 20170544. doi:10.1098/rspb.2017.0544. PMC 5454271. PMID 28566488.
- Pal Chowdhury M, Makarewicz C, Piezonka H, Buckley M (November 2022). "Novel Deep Eutectic Solvent-Based Protein Extraction Method for Pottery Residues and Archeological Implications". Journal of Proteome Research. 21 (11): 2619–2634. doi:10.1021/acs.jproteome.2c00340. PMC 9639204. PMID 36268809.
- Beslic D, Tscheuschner G, Renard BY, Weller MG, Muth T (January 2023). "Comprehensive evaluation of peptide de novo sequencing tools for monoclonal antibody assembly". Briefings in Bioinformatics. 24 (1). doi:10.1093/bib/bbac542. PMC 9851299. PMID 36545804.
| Genomics | |
|---|---|
| Bioinformatics | |
| Structural biology | |
| Research tools | |
| Organizations | |
| |