Purinosome
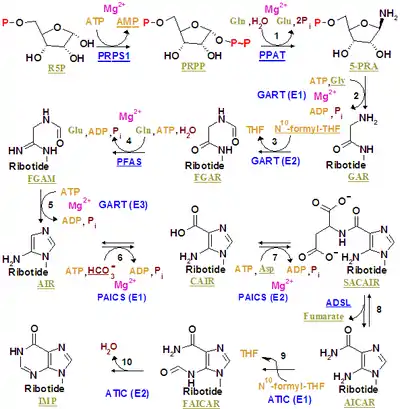
The purinosome is a putative multi-enzyme complex that carries out de novo purine biosynthesis within the cell. It is postulated to include all six of the human enzymes identified as direct participants in this ten-step biosynthetic pathway converting phosphoribosyl pyrophosphate to inosine monophosphate:
| Step(s) | Symbol | Description |
|---|---|---|
| 1 | PPAT | phosphoribosylpyrophosphate amidotransferase |
| 2,3,5 | GART | trifunctional phosphoribosylglycinamide formyltransferase/phosphoribosylglycinamide synthetase/phosphoribosylaminoimidazole synthetase |
| 4 | PFAS | phosphoribosylformylglycinamidine synthase |
| 6,7 | PAICS | bifunctional phosphoribosylaminoimidazole carboxylase |
| 8 | ADSL | adenylosuccinate lyase |
| 9,10 | ATIC | bifunctional 5-aminoimidazole-4-carboxamide ribonucleotide formyltransferase/IMP cyclohydrolase |
History
Hypothesis
The enzymes of the multi-step de novo purine biosynthesis pathway have been postulated to form a multi-enzyme complex to facilitate substrate channeling between each enzyme of the pathway. Slight variations of the pathway exists between phyla; however, there are 13 enzymes that can be considered part of this biosynthetic pathway.[1] Several individual enzymatic functions have consolidated onto single bifunctional or trifunctional polypeptide chains in higher organisms, suggesting stable physical interactions exist between enzymes.[2][3] The functional consolidation of steps 2,3, and 5 of the pathway into a single enzyme in higher organisms such as humans suggests physical local proximity of the enzyme for step 4 to the trifunctional enzyme.[2][4][5]
Evidence for a complex
The purine biosynthesis enzymes can be co-purified under certain conditions.[6][7] A complex of two particular pathway enzymes GART and ATIC can be isolated with cofactor production enzyme C1THF synthase and SHMT1.[8] Kinetic studies show evidence of substrate channeling between PPAT and GART, but evidence could not be obtained for their physical protein-protein interaction.[9] Thus far, isolation of a multienzyme complex inclusive of all purine biosynthesis enzymes has not been achieved.
Purinosome macrobodies

Purinosome macrobodies (also may be referred to as bodies, clusters, foci, puncta) describe the assembly of fluorescent-tagged human purine biosynthetic enzymes into bodies visible by fluorescence microscopy. The purinosome body theory states that purinosome bodies are assembled from proteins normally dispersed in the cell, and this assembly manifests when the demand for purines exceeds the amount supplied by the purine salvage pathway, such as when the extracellular medium is depleted of purines. In addition to the 6 purine biosynthesis pathway proteins, purinosome macrobodies are composed of at least 10 additional proteins not involved in purine biosynthesis. Due to the nature of their expression and association with cellular stress response proteins, purinosome macrobodies may actually be aggregated protein bodies.
Initial discovery
The human purinosome was thought to have been identified in 2008 by the observation that transiently expressed GFP fusion constructs of purine biosynthesis proteins form macrobodies.[11][12] A folate enzyme not directly involved in the purine biosynthesis pathway, 5,10-methenyltetrahydrofolate synthase (MTHFS), was later found to be part of purinosome macrobodies by the same approach.[13] The biological relevance of this folate enzyme's inclusion to the purinosome macrobody is unclear: while it provides substrate for a trifunctional folate enzyme C1THF synthase to generate a key cofactor for purine biosynthesis, C1THF synthase is not a part of purinosome macrobodies.[11] Curiously, hypoxanthine levels do not alter purinosome macrobodies,[11] but adenosine or guanosine addition suppresses formation of macromolecular bodies formed by the folate enzyme.[13]
Aggregation
Later studies in 2013 support the interpretation that those macrobodies could be artifacts of aggregated proteins that commonly result from fusion protein expression.[10] Characteristics of purinosome bodies were found to be shared between those of canonical protein aggregates, such as induction by peroxide. While purinosome bodies were also found to be associated with early cell death, it is unclear whether the bodies were a cause of that stress or rather an indicator of stressed cells.
Discrepancies
Inhibition of microtubule polymerization with nocodazole blocks formation of the purinosome macrobodies, and reduces the flux of de novo purine biosynthesis.[14] However, nocodazole also blocks formation of aggresomes, complicating interpretation of these observations. Partial inhibition of casein kinase 2 by small molecule inhibitors - 4,5,6,7-tetrabromo-1H-benzimidazole (TBI), 2-dimethylamino-4,5,6,7-tetrabromo-1H-benzimidazole (DMAT), tetrabromocinammic acid (TBCA) or ellagic acid - was found to induce purinosome macrobody formation, while another inhibitor, 4,5,6,7-tetrabromobenzotriazole (TBB) induced purinosome macrobody formation at low concentration but not at high concentration, and caused the dissociation of the bodies formed in response to DMAT.[15] Complicating the interpretation of these data, inhibition of casein kinase 2 is also known to disrupt hundreds of cellular processes, among them being protein homeostasis which regulates protein aggregation.
Additional members of purinosome macrobodies
References
- Kornberg, A. (1982). "Supplement to DNA Replication". San Francisco:Freeman.
- Henikoff, S.; Keene, M. A.; Sloan, S.; Bleskan, J.; Hards, R.; Patterson, D. (1986). "Multiple purine pathway enzyme activities are encoded at a single genetic locus in Drosophila". Proc. Natl. Acad. Sci. USA. 83 (3): 720–24. doi:10.1073/pnas.83.3.720. PMC 322936. PMID 3080748.
- Marcotte EM; Pellegrini M; Ng HL; Rice DW; Yeates TO; Eisenberg D. (1999). "Detecting protein function and protein-protein interactions from genome sequences". Science. 285 (5428): 751–3. CiteSeerX 10.1.1.535.9650. doi:10.1126/science.285.5428.751. PMID 10427000.
- Patterson, D.; Graw, S.; Jones, C. (1981). "Demonstration by somatic cell genetics, of coordinate regulation of genes for two enzymes of purine synthesis assigned to human chromosome 21". Proceedings of the National Academy of Sciences of the United States of America. 78 (1): 405–409. Bibcode:1981PNAS...78..405P. doi:10.1073/pnas.78.1.405. PMC 319062. PMID 6941256.
- Hard, R. G.; Benkovic, S. J.; Van Keuren, M. L.; Graw, S. L.; Drabkin, H. A.; Patterson, D. (1986). "Assignment of a third purine biosynthetic gene (glycinamide ribonucleotide transformylase) to human chromosome 21". American Journal of Human Genetics. 39 (2): 179–185. PMC 1683921. PMID 3529945.
- Rowe, P. B.; McCaims, E.; Madsen, G.; Sauer, D.; Elliott, H. (1978). "De novo purine synthesis in avian liver. Co-purification of the enzymes and properties of the pathway". J. Biol. Chem. 253 (21): 7711–21. doi:10.1016/S0021-9258(17)34428-9. PMID 701284.
- McCairns, E.; Fahey, D.; Sauer, D.; Rowe, P. B. (1983). "De novo purine synthesis in human lymphocytes. Partial co-purification of the enzymes and some properties of the pathway". J. Biol. Chem. 258 (3): 1851–56. doi:10.1016/S0021-9258(18)33066-7. PMID 6296113.
- Smith, G. K.; Mueller, W. T.; Wasserman, G. F.; Taylor, W. D.; Benkovic, S. J. (1980). "Characterization of the enzyme complex involving the folate-requiring enzymes of de novo purine biosynthesis". Biochemistry. 19 (18): 4313–21. doi:10.1021/bi00559a026. PMID 7417406.
- J. Rudolph; J. Stubbe (1995). "Investigation of the Mechanism of Phosphoribosylamine Transfer from Glutamine Phosphoribosylpyrophosphate Amidotransferase to Glycinamide Ribonucleotide Synthetase". Biochemistry. 34 (7): 2241–2250. doi:10.1021/bi00007a019. PMID 7532005.
- Alice Zhao; Mark Tsechansky; Jagannath Swaminathan; Lindsey Cook; Andrew D. Ellington; Edward M. Marcotte (2013-02-06). "Transiently Transfected Purine Biosynthetic Enzymes Form Stress Bodies". PLOS ONE. 8 (2): e56203. Bibcode:2013PLoSO...856203Z. doi:10.1371/journal.pone.0056203. PMC 3566086. PMID 23405267.
- Songon An; et al. (2008). "Reversible Compartmentalization of de Novo Purine Biosynthetic Complexes in Living Cells". Science. 320 (5872): 103–106. Bibcode:2008Sci...320..103A. doi:10.1126/science.1152241. PMID 18388293. S2CID 24119538.
- Hong Zhao; Jarrod B. French; Ye Fang; Stephen J. Benkovic (2013-04-03). "The purinosome, a multi-protein complex involved in the de novo biosynthesis of purines in humans". Chem. Commun. 285 (15): 11093–11099. doi:10.1039/c3cc41437j. PMC 3877848. PMID 23575936.
- Martha S. Field; Donald D. Anderson; Patrick J. Stover (2011). "Mthfs is an essential gene in mice and a component of the purinosome". Front Genet. 2 (36): 36. doi:10.3389/fgene.2011.00036. PMC 3268590. PMID 22303332.
- Songon An; Yijun Deng; John W. Tomsho; Minjoung Kyoung; Stephen J. Benkovic (2010-07-20). "Microtubule-assisted mechanism for functional metabolic macromolecular complex formation". Proc. Natl. Acad. Sci. U.S.A. 107 (29): 12872–12876. Bibcode:2010PNAS..10712872A. doi:10.1073/pnas.1008451107. PMC 2919939. PMID 20615962.
- Songon An; Minjoung Kyoung; Jasmina J. Allen; Kevan M. Shokat; Stephen J. Benkovic (2010-04-09). "Dynamic regulation of a metabolic multi-enzyme complex by protein kinase CK2". J Biol Chem. 285 (15): 11093–11099. doi:10.1074/jbc.M110.101139. PMC 2856985. PMID 20157113.
- French, J. B.; et al. (2013-01-28). "Hsp70/Hsp90 chaperone machinery is involved in the assembly of the purinosome". Proceedings of the National Academy of Sciences. 110 (7): 2528–2533. Bibcode:2013PNAS..110.2528F. doi:10.1073/pnas.1300173110. ISSN 0027-8424. PMC 3574928. PMID 23359685.
- Alice Zhao; Mark Tsechansky; Jagannath Swaminathan; Lindsey Cook; Andrew Ellington; Edward Marcotte. "By the same standards, prior work may not either" (PDF). Archived from the original (PDF) on 4 February 2014. Retrieved 20 April 2013.