Biochemical cascade
A biochemical cascade, also known as a signaling cascade or signaling pathway, is a series of chemical reactions that occur within a biological cell when initiated by a stimulus. This stimulus, known as a first messenger, acts on a receptor that is transduced to the cell interior through second messengers which amplify the signal and transfer it to effector molecules, causing the cell to respond to the initial stimulus.[1] Most biochemical cascades are series of events, in which one event triggers the next, in a linear fashion. At each step of the signaling cascade, various controlling factors are involved to regulate cellular actions, in order to respond effectively to cues about their changing internal and external environments.[1]
An example would be the coagulation cascade of secondary hemostasis which leads to fibrin formation, and thus, the initiation of blood coagulation. Another example, sonic hedgehog signaling pathway, is one of the key regulators of embryonic development and is present in all bilaterians.[2] Signaling proteins give cells information to make the embryo develop properly. When the pathway malfunctions, it can result in diseases like basal cell carcinoma.[3] Recent studies point to the role of hedgehog signaling in regulating adult stem cells involved in maintenance and regeneration of adult tissues. The pathway has also been implicated in the development of some cancers. Drugs that specifically target hedgehog signaling to fight diseases are being actively developed by a number of pharmaceutical companies.
Introduction
Signaling cascades
Cells require a full and functional cellular machinery to live. When they belong to complex multicellular organisms, they need to communicate among themselves and work for symbiosis in order to give life to the organism. These communications between cells triggers intracellular signaling cascades, termed signal transduction pathways, that regulate specific cellular functions. Each signal transduction occurs with a primary extracellular messenger that binds to a transmembrane or nuclear receptor, initiating intracellular signals. The complex formed produces or releases second messengers that integrate and adapt the signal, amplifying it, by activating molecular targets, which in turn trigger effectors that will lead to the desired cellular response.[4]
Transductors and effectors
Signal transduction is realized by activation of specific receptors and consequent production/delivery of second messengers, such as Ca2+ or cAMP. These molecules operate as signal transducers, triggering intracellular cascades and in turn amplifying the initial signal.[4] Two main signal transduction mechanisms have been identified, via nuclear receptors, or via transmembrane receptors. In the first one, first messenger cross through the cell membrane, binding and activating intracellular receptors localized at nucleus or cytosol, which then act as transcriptional factors regulating directly gene expression. This is possible due to the lipophilic nature of those ligands, mainly hormones. In the signal transduction via transmembrane receptors, the first messenger binds to the extracellular domain of transmembrane receptor, activating it. These receptors may have intrinsic catalytic activity or may be coupled to effector enzymes, or may also be associated to ionic channels. Therefore, there are four main transmembrane receptor types: G protein coupled receptors (GPCRs), tyrosine kinase receptors (RTKs), serine/threonine kinase receptors (RSTKs), and ligand-gated ion channels (LGICs).[1][4] Second messengers can be classified into three classes:
- Hydrophilic/cytosolic – are soluble in water and are localized at the cytosol, including cAMP, cGMP, IP3, Ca2+, cADPR and S1P. Their main targets are protein kinases as PKA and PKG, being then involved in phosphorylation mediated responses.[4]
- Hydrophobic/membrane-associated – are insoluble in water and membrane-associated, being localized at intermembrane spaces, where they can bind to membrane-associated effector proteins. Examples: PIP3, DAG, phosphatidic acid, arachidonic acid and ceramide. They are involved in regulation of kinases and phosphatases, G protein associated factors and transcriptional factors.[4]
- Gaseous – can be widespread through cell membrane and cytosol, including nitric oxide and carbon monoxide. Both of them can activate cGMP and, besides of being capable of mediating independent activities, they also can operate in a coordinated mode.[4]
Cellular response
The cellular response in signal transduction cascades involves alteration of the expression of effector genes or activation/inhibition of targeted proteins. Regulation of protein activity mainly involves phosphorylation/dephosphorylation events, leading to its activation or inhibition. It is the case for the vast majority of responses as a consequence of the binding of the primary messengers to membrane receptors. This response is quick, as it involves regulation of molecules that are already present in the cell. On the other hand, the induction or repression of the expression of genes requires the binding of transcriptional factors to the regulatory sequences of these genes. The transcriptional factors are activated by the primary messengers, in most cases, due to their function as nuclear receptors for these messengers. The secondary messengers like DAG or Ca2+ could also induce or repress gene expression, via transcriptional factors. This response is slower than the first because it involves more steps, like transcription of genes and then the effect of newly formed proteins in a specific target. The target could be a protein or another gene.[1][4][5]
Examples of biochemical cascades
In biochemistry, several important enzymatic cascades and signal transduction cascades participate in metabolic pathways or signaling networks, in which enzymes are usually involved to catalyze the reactions. For example, the tissue factor pathway in the coagulation cascade of secondary hemostasis is the primary pathway leading to fibrin formation, and thus, the initiation of blood coagulation. The pathways are a series of reactions, in which a zymogen (inactive enzyme precursor) of a serine protease and its glycoprotein co-factors are activated to become active components that then catalyze the next reaction in the cascade, ultimately resulting in cross-linked fibrin.[6]
Another example, sonic hedgehog signaling pathway, is one of the key regulators of embryonic development and is present in all bilaterians.[2] Different parts of the embryo have different concentrations of hedgehog signaling proteins, which give cells information to make the embryo develop properly and correctly into a head or a tail. When the pathway malfunctions, it can result in diseases like basal cell carcinoma.[3] Recent studies point to the role of hedgehog signaling in regulating adult stem cells involved in maintenance and regeneration of adult tissues. The pathway has also been implicated in the development of some cancers. Drugs that specifically target hedgehog signaling to fight diseases are being actively developed by a number of pharmaceutical companies.[7] Most biochemical cascades are series of events, in which one event triggers the next, in a linear fashion.
Biochemical cascades include:
- The Complement system
- The Insulin Signaling Pathway
- The Sonic hedgehog Signaling Pathway
- The Wnt signaling pathway
- The JAK-STAT signaling pathway
- The Adrenergic receptor Pathways
- The Acetylcholine receptor Pathways
- The Mitogen-activated protein kinase cascade
Conversely, negative cascades include events that are in a circular fashion, or can cause or be caused by multiple events.[8] Negative cascades include:
Cell-specific biochemical cascades
Epithelial cells
Adhesion is an essential process to epithelial cells so that epithelium can be formed and cells can be in permanent contact with extracellular matrix and other cells. Several pathways exist to accomplish this communication and adhesion with environment. But the main signalling pathways are the cadherin and integrin pathways.[9] The cadherin pathway is present in adhesion junctions or in desmosomes and it is responsible for epithelial adhesion and communication with adjacent cells. Cadherin is a transmembrane glycoprotein receptor that establishes contact with another cadherin present in the surface of a neighbour cell forming an adhesion complex.[10] This adhesion complex is formed by β-catenin and α-catenin, and p120CAS is essential for its stabilization and regulation. This complex then binds to actin, leading to polymerization. For actin polymerization through the cadherin pathway, proteins of the Rho GTPases family are also involved. This complex is regulated by phosphorylation, which leads to downregulation of adhesion. Several factors can induce the phosphorylation, like EGF, HGF or v-Src. The cadherin pathway also has an important function in survival and proliferation because it regulates the concentration of cytoplasmic β-catenin. When β-catenin is free in the cytoplasm, normally it is degraded, however if the Wnt signalling is activated, β-catenin degradation is inhibited and it is translocated to the nucleus where it forms a complex with transcription factors. This leads to activation of genes responsible for cell proliferation and survival. So the cadherin-catenin complex is essential for cell fate regulation.[11][12] Integrins are heterodimeric glycoprotein receptors that recognize proteins present in the extracellular matrix, like fibronectin and laminin. In order to function, integrins have to form complexes with ILK and Fak proteins. For adhesion to the extracellular matrix, ILK activate the Rac and Cdc42 proteins and leading to actin polymerization. ERK also leads to actin polymerization through activation of cPLA2. Recruitment of FAK by integrin leads to Akt activation and this inhibits pro-apoptotic factors like BAD and Bax. When adhesion through integrins do not occur the pro-apoptotic factors are not inhibited and resulting in apoptosis.[13][14]
Hepatocytes
The hepatocyte is a complex and multifunctional differentiated cell whose cell response will be influenced by the zone in hepatic lobule, because concentrations of oxygen and toxic substances present in the hepatic sinusoids change from periportal zone to centrilobular zone10. The hepatocytes of the intermediate zone have the appropriate morphological and functional features since they have the environment with average concentrations of oxygen and other substances.[15] This specialized cell is capable of:[16]
- Regulate glucose metabolism[4][5][17]
- Via cAMP/PKA/TORC (transducers of regulated CREB)/CRE, PIP3 /PKB and PLC /IP3
- Expression of enzymes for synthesis, storage and distribution of glucose
- Synthesis of acute phase proteins[18][19][20]
- Via JAK /STAT /APRE (acute phase response element)
- Expression of C-reactive protein, globulin protease inhibitors, complement, coagulation and fibrinolytic systems and iron homeostasis
- Regulate iron homeostasis (acute phase independent)[4][20][21]
- Regulate lipid metabolism[4][20][22][23]
- Exocrine production of bile salts and other compounds[4][20][22][23]
- Via LXR /LXRE
- Expression of CYP7A1 and ABC transporters
- Via LXR /LXRE
- Expression of ABC transporters
- Endocrine production
- Angiotensinogen expression
- Via STAT and Gab1: RAS/MAPK, PLC/IP3 and PI3K/FAK
- Cell growth, proliferation, survival, invasion and motility
The hepatocyte also regulates other functions for constitutive synthesis of proteins (albumin, ALT and AST) that influences the synthesis or activation of other molecules (synthesis of urea and essential amino acids), activate vitamin D, utilization of vitamin K, transporter expression of vitamin A and conversion of thyroxine.[15][30]
Neurons
Purinergic signalling has an essential role at interactions between neurons and glia cells, allowing these to detect action potentials and modulate neuronal activity, contributing for intra and extracellular homeostasis regulation. Besides purinergic neurotransmitter, ATP acts as a trophic factor at cellular development and growth, being involved on microglia activation and migration, and also on axonal myelination by oligodendrocytes. There are two main types of purinergic receptors, P1 binding to adenosine, and P2 binding to ATP or ADP, presenting different signalling cascades.[31][32] The Nrf2/ARE signalling pathway has a fundamental role at fighting against oxidative stress, to which neurons are especially vulnerable due to its high oxygen consumption and high lipid content. This neuroprotective pathway involves control of neuronal activity by perisynaptic astrocytes and neuronal glutamate release, with the establishment of tripartite synapses. The Nrf2/ARE activation leads to a higher expression of enzymes involved in glutathione syntheses and metabolism, that have a key role in antioxidant response.[33][34][35][36] The LKB1/NUAK1 signalling pathway regulates terminal axon branching at cortical neurons, via local immobilized mitochondria capture. Besides NUAK1, LKB1 kinase acts under other effectors enzymes as SAD-A/B and MARK, therefore regulating neuronal polarization and axonal growth, respectively. These kinase cascades implicates also Tau and others MAP.[37][38][39] An extended knowledge of these and others neuronal pathways could provide new potential therapeutic targets for several neurodegenerative chronic diseases as Alzheimer's, Parkinson's and Huntington's disease, and also amyotrophic lateral sclerosis.[31][32][33]
Blood cells
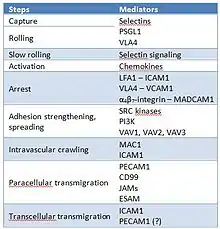
The blood cells (erythrocytes, leukocytes and platelets) are produced by hematopoiesis. The erythrocytes have as main function the O2 delivery to the tissues, and this transfer occurs by diffusion and is determined by the O2 tension (PO2). The erythrocyte is able to feel the tissue need for O2 and cause a change in vascular caliber, through the pathway of ATP release, which requires an increase in cAMP, and are regulated by the phosphodiesterase (PDE). This pathway can be triggered via two mechanisms: physiological stimulus (like reduced O2 tension) and activation of the prostacyclin receptor (IPR). This pathway includes heterotrimeric G proteins, adenylyl cyclase (AC), protein kinase A (PKA), cystic fibrosis transmembrane conductance regulator (CFTR), and a final conduit that transport ATP to vascular lumen (pannexin 1 or voltage-dependent anion channel (VDAC)). The released ATP acts on purinergic receptors on endothelial cells, triggering the synthesis and release of several vasodilators, like nitric oxide (NO) and prostacyclin (PGI2).[40][41] The current model of leukocyte adhesion cascade includes many steps mentioned in Table 1.[42] The integrin-mediated adhesion of leukocytes to endothelial cells is related with morphological changes in both leukocytes and endothelial cells, which together support leukocyte migration through the venular walls. Rho and Ras small GTPases are involved in the principal leukocyte signaling pathways underlying chemokine-stimulated integrin-dependent adhesion, and have important roles in regulating cell shape, adhesion and motility.[43]
After a vascular injury occurs, platelets are activated by locally exposed collagen (glycoprotein (GP) VI receptor), locally generated thrombin (PAR1 and PAR4 receptors), platelet-derived thromboxane A2 (TxA2) (TP receptor) and ADP (P2Y1 and P2Y12 receptors) that is either released from damaged cells or secreted from platelet dense granules. The von Willebrand factor (VWF) serves as an essential accessory molecule. In general terms, platelet activation initiated by agonist takes to a signaling cascade that leads to an increase of the cytosolic calcium concentration. Consequently, the integrin αIIbβ3 is activated and the binding to fibrinogen allows the aggregation of platelets to each other. The increase of cytosolic calcium also leads to shape change and TxA2 synthesis, leading to signal amplification.
Lymphocytes
The main goal of biochemical cascades in lymphocytes is the secretion of molecules that can suppress altered cells or eliminate pathogenic agents, through proliferation, differentiation and activation of these cells. Therefore, the antigenic receptors play a central role in signal transduction in lymphocytes, because when antigens interact with them lead to a cascade of signal events. These receptors, that recognize the antigen soluble (B cells) or linked to a molecule on Antigen Presenting Cells (T cells), do not have long cytoplasm tails, so they are anchored to signal proteins, which contain a long cytoplasmic tails with a motif that can be phosphorylated (ITAM – immunoreceptor tyrosine-based activation motif) and resulting in different signal pathways. The antigen receptor and signal protein form a stable complex, named BCR or TCR, in B or T cells, respectively. The family Src is essential for signal transduction in these cells, because it is responsible for phosphorylation of ITAMs. Therefore, Lyn and Lck, in lymphocytes B and T, respectively, phosphorylate immunoreceptor tyrosine-based activation motifs after the antigen recognition and the conformational change of the receptor, which leads to the binding of Syk/Zap-70 kinases to ITAM and its activation. Syk kinase is specific of lymphocytes B and Zap-70 is present in T cells. After activation of these enzymes, some adaptor proteins are phosphorylated, like BLNK (B cells) and LAT (T cells). These proteins after phosphorylation become activated and allow binding of others enzymes that continue the biochemical cascade.[4][44][45][46] One example of a protein that binds to adaptor proteins and become activated is PLC that is very important in the lymphocyte signal pathways. PLC is responsible for PKC activation, via DAG and Ca2+, which leads to phosphorylation of CARMA1 molecule, and formation of CBM complex. This complex activates Iκκ kinase, which phosphorylates I-κB, and then allows the translocation of NF-κB to the nucleus and transcription of genes encoding cytokines, for example. Others transcriptional factors like NFAT and AP1 complex are also important for transcription of cytokines.[45][47][48][49] The differentiation of B cells to plasma cells is also an example of a signal mechanism in lymphocytes, induced by a cytokine receptor. In this case, some interleukins bind to a specific receptor, which leads to activation of MAPK/ERK pathway. Consequently, the BLIMP1 protein is translated and inhibits PAX5, allowing immunoglobulin genes transcription and activation of XBP1 (important for the secretory apparatus formation and enhancing of protein synthesis).[50][51][52] Also, the coreceptors (CD28/CD19) play an important role because they can improve the antigen/receptor binding and initiate parallel cascade events, like activation o PI3 Kinase. PIP3 then is responsible for activation of several proteins, like vav (leads to activation of JNK pathway, which consequently leads to activation of c-Jun) and btk (can also activate PLC).[45][53]
Wnt signaling pathway
The Wnt signaling pathway can be divided in canonical and non-canonical. The canonical signaling involves binding of Wnt to Frizzled and LRP5 co-receptor, leading to GSK3 phosphorylation and inhibition of β-catenin degradation, resulting in its accumulation and translocation to the nucleus, where it acts as a transcription factor. The non-canonical Wnt signaling can be divided in planar cell polarity (PCP) pathway and Wnt/calcium pathway. It is characterized by binding of Wnt to Frizzled and activation of G proteins and to an increase of intracellular levels of calcium through mechanisms involving PKC 50.[54] The Wnt signaling pathway plays a significant role in osteoblastogenesis and bone formation, inducing the differentiation of mesenquimal pluripotent cells in osteoblasts and inhibiting the RANKL/RANK pathway and osteoclastogenesis.[55]
RANKL/RANK signaling pathway
RANKL is a member of the TNF superfamily of ligands. Through binding to the RANK receptor it activates various molecules, like NF-kappa B, MAPK, NFAT and PI3K52. The RANKL/RANK signaling pathway regulates osteoclastogenesis, as well as, the survival and activation of osteoclasts.[56][57]
Adenosine signaling pathway
Adenosine is very relevant in bone metabolism, as it plays a role in formation and activation of both osteoclasts and osteoblasts. Adenosine acts by binding to purinergic receptors and influencing adenylyl cyclase activity and the formation of cAMP and PKA 54.[58] Adenosine may have opposite effects on bone metabolism, because while certain purinergic receptors stimulate adenylyl cyclase activity, others have the opposite effect.[58][59] Under certain circumstances adenosine stimulates bone destruction and in other situations it promotes bone formation, depending on the purinergic receptor that is being activated.
Stem cells
Self-renewal and differentiation abilities are exceptional properties of stem cells. These cells can be classified by their differentiation capacity, which progressively decrease with development, in totipotents, pluripotents, multipotents and unipotents.[60]
Self-renewal process is highly regulated from cell cycle and genetic transcription control. There are some signaling pathways, such as LIF/JAK/STAT3 (Leukemia inhibitory factor/Janus kinase/Signal transducer and activator of transcription 3) and BMP/SMADs/Id (Bone morphogenetic proteins/ Mothers against decapentaplegic/ Inhibitor of differentiation), mediated by transcription factors, epigenetic regulators and others components, and they are responsible for self-renewal genes expression and inhibition of differentiation genes expression, respectively.[61]
At cell cycle level there is an increase of complexity of the mechanisms in somatic stem cells. However, it is observed a decrease of self-renewal potential with age. These mechanisms are regulated by p16Ink4a-CDK4/6-Rb and p19Arf-p53-P21Cip1 signaling pathways. Embryonic stem cells have constitutive cyclin E-CDK2 activity, which hyperphosphorylates and inactivates Rb. This leads to a short G1 phase of the cell cycle with rapid G1-S transition and little dependence on mitogenic signals or D cyclins for S phase entry. In fetal stem cells, mitogens promote a relatively rapid G1-S transition through cooperative action of cyclin D-CDK4/6 and cyclin E-CDK2 to inactivate Rb family proteins. p16Ink4a and p19Arf expression are inhibited by Hmga2-dependent chromatin regulation. Many young adult stem cells are quiescent most of the time. In the absence of mitogenic signals, cyclin-CDKs and the G1-S transition are suppressed by cell cycle inhibitors including Ink4 and Cip/Kip family proteins. As a result, Rb is hypophosphorylated and inhibits E2F, promoting quiescence in G0-phase of the cell cycle. Mitogen stimulation mobilizes these cells into cycle by activating cyclin D expression. In old adult stem cells, let-7 microRNA expression increases, reducing Hmga2 levels and increasing p16Ink4a and p19Arf levels. This reduces the sensitivity of stem cells to mitogenic signals by inhibiting cyclin-CDK complexes. As a result, either stem cells cannot enter the cell cycle, or cell division slows in many tissues.[62]
Extrinsic regulation is made by signals from the niche, where stem cells are found, which is able to promote quiescent state and cell cycle activation in somatic stem cells.[63] Asymmetric division is characteristic of somatic stem cells, maintaining the reservoir of stem cells in the tissue and production of specialized cells of the same.[64]
Stem cells show an elevated therapeutic potential, mainly in hemato-oncologic pathologies, such as leukemia and lymphomas. Little groups of stem cells were found into tumours, calling cancer stem cells. There are evidences that these cells promote tumor growth and metastasis.[65]
Oocytes
The oocyte is the female cell involved in reproduction.[66] There is a close relationship between the oocyte and the surrounding follicular cells which is crucial to the development of both.[67] GDF9 and BMP15 produced by the oocyte bind to BMPR2 receptors on follicular cells activating SMADs 2/3, ensuring follicular development.[68] Concomitantly, oocyte growth is initiated by binding of KITL to its receptor KIT in the oocyte, leading to the activation of PI3K/Akt pathway, allowing oocyte survival and development.[69] During embryogenesis, oocytes initiate meiosis and stop in prophase I. This arrest is maintained by elevated levels of cAMP within the oocyte.[70] It was recently suggested that cGMP cooperates with cAMP to maintain the cell cycle arrest.[70][71] During meiotic maturation, the LH peak that precedes ovulation activates MAPK pathway leading to gap junction disruption and breakdown of communication between the oocyte and the follicular cells. PDE3A is activated and degrades cAMP, leading to cell cycle progression and oocyte maturation.[72][73] The LH surge also leads to the production of progesterone and prostaglandins that induce the expression of ADAMTS1 and other proteases, as well as their inhibitors. This will lead to degradation of the follicular wall, but limiting the damage and ensuring that the rupture occurs in the appropriate location, releasing the oocyte into the Fallopian tubes.[74][75] Oocyte activation depends on fertilization by sperm.[76] It is initiated with sperm's attraction induced by prostaglandins produced by the oocyte, which will create a gradient that will influence the sperm's direction and velocity.[77] After fusion with the oocyte, PLC ζ of the spermatozoa is released into the oocyte leading to an increase in Ca2+ levels that will activate CaMKII which will degrade MPF, leading to the resumption of meiosis.[78][79] The increased Ca2+ levels will induce the exocytosis of cortical granules that degrade ZP receptors, used by sperm to penetrate the oocyte, blocking polyspermy.[80] Deregulation of these pathways will lead to several diseases like, oocyte maturation failure syndrome which results in infertility.[81] Increasing our molecular knowledge of oocyte development mechanisms could improve the outcome of assisted reproduction procedures, facilitating conception.
Spermatozoon
Spermatozoon is the male gamete. After ejaculation this cell is not mature, so it can not fertilize the oocyte. To have the ability to fertilize the female gamete, this cell suffers capacitation and acrosome reaction in female reproductive tract. The signaling pathways best described for spermatozoon involve these processes. The cAMP/PKA signaling pathway leads to sperm cells capacitation; however, adenylyl cyclase in sperm cells is different from the somatic cells. Adenylyl cyclase in spermatozoon does not recognize G proteins, so it is stimulated by bicarbonate and Ca2+ ions. Then, it converts adenosine triphosphate into cyclic AMP, which activates Protein kinase A. PKA leads to protein tyrosine phosphorylation.[82][83][84] Phospholipase C (PLC) is involved in acrosome reaction. ZP3 is a glycoprotein present in zona pelucida and it interacts with receptors in spermatozoon. So, ZP3 can activate G protein coupled receptors and tyrosine kinase receptors, that leads to production of PLC. PLC cleaves the phospholipid phosphatidylinositol 4,5-bisphosphate (PIP2) into diacyl glycerol (DAG) and inositol 1,4,5-trisphosphate. IP3 is released as a soluble structure into the cytosol and DAG remains bound to the membrane. IP3 binds to IP3 receptors, present in acrosome membrane. In addition, calcium and DAG together work to activate protein kinase C, which goes on to phosphorylate other molecules, leading to altered cellular activity. These actions cause an increase in cytosolic concentration of Ca2+ that leads to dispersion of actin and consequently promotes plasmatic membrane and outer acrosome membrane fusion.[85][86] Progesterone is a steroid hormone produced in cumulus oophorus. In somatic cells it binds to receptors in nucleus; however, in spermatozoon its receptors are present in plasmatic membrane. This hormone activates AKT that leads to activation of other protein kinases, involved in capacitation and acrosome reaction.[87][88] When ROS (reactive oxygen species) are present in high concentration, they can affect the physiology of cells, but when they are present in moderated concentration they are important for acrosome reaction and capacitation. ROS can interact with cAMP/PKA and progesterone pathway, stimulating them. ROS also interacts with ERK pathway that leads to activation of Ras, MEK and MEK-like proteins. These proteins activate protein tyrosine kinase (PTK) that phosphorylates various proteins important for capacitation and acrosome reaction.[89][90]
Embryos
Various signalling pathways, as FGF, WNT and TGF-β pathways, regulate the processes involved in embryogenesis.
FGF (Fibroblast Growth Factor) ligands bind to receptors tyrosine kinase, FGFR (Fibroblast Growth Factor Receptors), and form a stable complex with co-receptors HSPG (Heparan Sulphate Proteoglycans) that will promote autophosphorylation of the intracellular domain of FGFR and consequent activation of four main pathways: MAPK/ERK, PI3K, PLCγ and JAK/STAT.[91][92][93]
- MAPK/ERK (Mitogen-Activated Protein Kinase/Extracellular Signal-Regulated Kinase) regulates gene transcription through successive kinase phosphorylation and in human embryonic stem cells it helps maintaining pluripotency.[93][94] However, in the presence of Activin A, a TGF-β ligand, it causes the formation of mesoderm and neuroectoderm.[95]
- Phosphorylation of membrane phospholipids by PI3K (Phosphatidylinositol 3-Kinase) results in activation of AKT/PKB (Protein Kinase B). This kinase is involved in cell survival and inhibition of apoptosis, cellular growth and maintenance of pluripotency, in embryonic stem cells.[93][96][97]
- PLCγ (Phosphoinositide Phospholipase C γ) hydrolyzes membrane phospholipids to form IP3 (Inositoltriphosphate) and DAG (Diacylglycerol), leading to activation of kinases and regulating morphogenic movements during gastrulation and neurulation.[91][92][98]
- STAT (Signal Trandsducer and Activator of Transcription) is phosphorylated by JAK (Janus Kinase) and regulates gene transcription, determining cell fates. In mouse embryonic stem cells, this pathway helps maintaining pluripotency.[92][93]
The WNT pathway allows β-catenin function in gene transcription, once the interaction between WNT ligand and G protein-coupled receptor Frizzled inhibits GSK-3 (Glycogen Synthase Kinase-3) and thus formation of β-catenin destruction complex.[93][99][100] Although there is some controversy about the effects of this pathway in embryogenesis, it is thought that WNT signalling induces primitive streak, mesoderm and endoderm formation.[100] In TGF-β (Transforming Growth Factor β) pathway, BMP (Bone Morphogenic Protein), Activin and Nodal ligands bind to their receptors and activate Smads that bind to DNA and promote gene transcription.[93][101][102] Activin is necessary for mesoderm and specially endoderm differentiation, and Nodal and BMP are involved in embryo patterning. BMP is also responsible for formation of extra-embryonic tissues before and during gastrulation, and for early mesoderm differentiation, when Activin and FGF pathways are activated.[101][102][103]
Pathway construction
Pathway building has been performed by individual groups studying a network of interest (e.g., immune signaling pathway) as well as by large bioinformatics consortia (e.g., the Reactome Project) and commercial entities (e.g., Ingenuity Systems). Pathway building is the process of identifying and integrating the entities, interactions, and associated annotations, and populating the knowledge base. Pathway construction can have either a data-driven objective (DDO) or a knowledge-driven objective (KDO). Data-driven pathway construction is used to generate relationship information of genes or proteins identified in a specific experiment such as a microarray study.[104] Knowledge-driven pathway construction entails development of a detailed pathway knowledge base for particular domains of interest, such as a cell type, disease, or system. The curation process of a biological pathway entails identifying and structuring content, mining information manually and/or computationally, and assembling a knowledgebase using appropriate software tools.[105] A schematic illustrating the major steps involved in the data-driven and knowledge-driven construction processes.[104]
For either DDO or KDO pathway construction, the first step is to mine pertinent information from relevant information sources about the entities and interactions. The information retrieved is assembled using appropriate formats, information standards, and pathway building tools to obtain a pathway prototype. The pathway is further refined to include context-specific annotations such as species, cell/tissue type, or disease type. The pathway can then be verified by the domain experts and updated by the curators based on appropriate feedback.[106] Recent attempts to improve knowledge integration have led to refined classifications of cellular entities, such as GO, and to the assembly of structured knowledge repositories.[107] Data repositories, which contain information regarding sequence data, metabolism, signaling, reactions, and interactions are a major source of information for pathway building.[108] A few useful databases are described in the following table.[104]
| Database | Curation Type | GO Annotation (Y/N) | Description | |
|---|---|---|---|---|
| 1. Protein-protein interactions databases | ||||
| BIND | Manual Curation | N | 200,000 documented biomolecular interactions and complexes | |
| MINT | Manual Curation | N | Experimentally verified interactions | |
| HPRD | Manual Curation | N | Elegant and comprehensive presentation of the interactions, entities and evidences | |
| MPact | Manual and Automated Curation | N | Yeast interactions. A part of MIPS | |
| DIP | Manual and Automated Curation | Y | Experimentally determined interactions | |
| IntAct | Manual Curation | Y | Database and analysis system of binary and multi-protein interactions | |
| PDZBase | Manual Curation | N | PDZ Domain containing proteins | |
| GNPV | Manual and Automated Curation | Y | Based on specific experiments and literature | |
| BioGrid | Manual Curation | Y | Physical and genetic interactions | |
| UniHi | Manual and Automated Curation | Y | Comprehensive human protein interactions | |
| OPHID | Manual Curation | Y | Combines PPI from BIND, HPRD, and MINT | |
| 2. Metabolic Pathway databases | ||||
| EcoCyc | Manual and Automated Curation | Y | Entire genome and biochemical machinery of E. Coli | |
| MetaCyc | Manual Curation | N | Pathways of over 165 species | |
| HumanCyc | Manual and Automated Curation | N | Human metabolic pathways and the human genome | |
| BioCyc | Manual and Automated Curation | N | Collection of databases for several organism | |
| 3. Signaling Pathway databases | ||||
| KEGG | Manual Curation | Y | Comprehensive collection of pathways such as human disease, signaling, genetic information processing pathways. Links to several useful databases | |
| PANTHER | Manual Curation | N | Compendium of metabolic and signaling pathways built using CellDesigner. Pathways can be downloaded in SBML format | |
| Reactome | Manual Curation | Y | Hierarchical layout. Extensive links to relevant databases such as NCBI, ENSEMBL, UNIPROT, HAPMAP, KEGG, CHEBI, PubMed, GO. Follows PSI-MI standards | |
| Biomodels | Manual Curation | Y | Domain experts curated biological connection maps and associated mathematical models | |
| STKE | Manual Curation | N | Repository of canonical pathways | |
| Ingenuity Systems | Manual Curation | Y | Commercial mammalian biological knowledgebase about genes, drugs, chemical, cellular and disease processes, and signaling and metabolic pathways | |
| Human signaling network | Manual Curation | Y | Literature-curated human signaling network, the largest human signaling network database | |
| PID | Manual Curation | Y | Compendium of several highly structured, assembled signaling pathways | |
| BioPP | Manual and Automated Curation | Y | Repository of biological pathways built using CellDesigner |
Legend: Y – Yes, N – No; BIND – Biomolecular Interaction Network Database, DIP – Database of Interacting Proteins, GNPV – Genome Network Platform Viewer, HPRD = Human Protein Reference Database, MINT – Molecular Interaction database, MIPS – Munich Information center for Protein Sequences, UNIHI – Unified Human Interactome, OPHID – Online Predicted Human Interaction Database, EcoCyc – Encyclopaedia of E. Coli Genes and Metabolism, MetaCyc – aMetabolic Pathway database, KEGG – Kyoto Encyclopedia of Genes and Genomes, PANTHER – Protein Analysis Through Evolutionary Relationship database, STKE – Signal Transduction Knowledge Environment, PID – The Pathway Interaction Database, BioPP – Biological Pathway Publisher. A comprehensive list of resources can be found at http://www.pathguide.org.
Pathway-related databases and tools
KEGG
The increasing amount of genomic and molecular information is the basis for understanding higher-order biological systems, such as the cell and the organism, and their interactions with the environment, as well as for medical, industrial and other practical applications. The KEGG resource[109] provides a reference knowledge base for linking genomes to biological systems, categorized as building blocks in the genomic space (KEGG GENES), the chemical space (KEGG LIGAND), wiring diagrams of interaction networks and reaction networks (KEGG PATHWAY), and ontologies for pathway reconstruction (BRITE database).[110] The KEGG PATHWAY database is a collection of manually drawn pathway maps for metabolism, genetic information processing, environmental information processing such as signal transduction, ligand–receptor interaction and cell communication, various other cellular processes and human diseases, all based on extensive survey of published literature.[111]
GenMAPP
Gene Map Annotator and Pathway Profiler (GenMAPP)[112] a free, open-source, stand-alone computer program is designed for organizing, analyzing, and sharing genome scale data in the context of biological pathways. GenMAPP database support multiple gene annotations and species as well as custom species database creation for a potentially unlimited number of species.[113] Pathway resources are expanded by utilizing homology information to translate pathway content between species and extending existing pathways with data derived from conserved protein interactions and coexpression. A new mode of data visualization including time-course, single nucleotide polymorphism (SNP), and splicing, has been implemented with GenMAPP database to support analysis of complex data. GenMAPP also offers innovative ways to display and share data by incorporating HTML export of analyses for entire sets of pathways as organized web pages.[114] In short, GenMAPP provides a means to rapidly interrogate complex experimental data for pathway-level changes in a diverse range of organisms.
Reactome
Given the genetic makeup of an organism, the complete set of possible reactions constitutes its reactome. Reactome, located at http://www.reactome.org is a curated, peer-reviewed resource of human biological processes/pathway data. The basic unit of the Reactome database is a reaction; reactions are then grouped into causal chains to form pathways[115] The Reactome data model allows us to represent many diverse processes in the human system, including the pathways of intermediary metabolism, regulatory pathways, and signal transduction, and high-level processes, such as the cell cycle.[116] Reactome provides a qualitative framework, on which quantitative data can be superimposed. Tools have been developed to facilitate custom data entry and annotation by expert biologists, and to allow visualization and exploration of the finished dataset as an interactive process map.[117] Although the primary curational domain is pathways from Homo sapiens, electronic projections of human pathways onto other organisms are regularly created via putative orthologs, thus making Reactome relevant to model organism research communities. The database is publicly available under open source terms, which allows both its content and its software infrastructure to be freely used and redistributed. Studying whole transcriptional profiles and cataloging protein–protein interactions has yielded much valuable biological information, from the genome or proteome to the physiology of an organism, an organ, a tissue or even a single cell. The Reactome database containing a framework of possible reactions which, when combined with expression and enzyme kinetic data, provides the infrastructure for quantitative models, therefore, an integrated view of biological processes, which links such gene products and can be systematically mined by using bioinformatics applications.[118] Reactome data available in a variety of standard formats, including BioPAX, SBML and PSI-MI, and also enable data exchange with other pathway databases, such as the Cycs, KEGG and amaze, and molecular interaction databases, such as BIND and HPRD. The next data release will cover apoptosis, including the death receptor signaling pathways, and the Bcl2 pathways, as well as pathways involved in hemostasis. Other topics currently under development include several signaling pathways, mitosis, visual phototransduction and hematopoeisis.[119] In summary, Reactome provides high-quality curated summaries of fundamental biological processes in humans in a form of biologist-friendly visualization of pathways data, and is an open-source project.
Pathway-oriented approaches
In the post-genomic age, high-throughput sequencing and gene/protein profiling techniques have transformed biological research by enabling comprehensive monitoring of a biological system, yielding a list of differentially expressed genes or proteins, which is useful in identifying genes that may have roles in a given phenomenon or phenotype.[120] With DNA microarrays and genome-wide gene engineering, it is possible to screen global gene expression profiles to contribute a wealth of genomic data to the public domain. With RNA interference, it is possible to distill the inferences contained in the experimental literature and primary databases into knowledge bases that consist of annotated representations of biological pathways. In this case, individual genes and proteins are known to be involved in biological processes, components, or structures, as well as how and where gene products interact with each other.[121][122] Pathway-oriented approaches for analyzing microarray data, by grouping long lists of individual genes, proteins, and/or other biological molecules according to the pathways they are involved in into smaller sets of related genes or proteins, which reduces the complexity, have proven useful for connecting genomic data to specific biological processes and systems. Identifying active pathways that differ between two conditions can have more explanatory power than a simple list of different genes or proteins. In addition, a large number of pathway analytic methods exploit pathway knowledge in public repositories such as Gene Ontology (GO) or Kyoto Encyclopedia of Genes and Genomes (KEGG), rather than inferring pathways from molecular measurements.[123][124] Furthermore, different research focuses have given the word "pathway" different meanings. For example, 'pathway' can denote a metabolic pathway involving a sequence of enzyme-catalyzed reactions of small molecules, or a signaling pathway involving a set of protein phosphorylation reactions and gene regulation events. Therefore, the term "pathway analysis" has a very broad application. For instance, it can refer to the analysis physical interaction networks (e.g., protein–protein interactions), kinetic simulation of pathways, and steady-state pathway analysis (e.g., flux-balance analysis), as well as its usage in the inference of pathways from expression and sequence data. Several functional enrichment analysis tools[125][126][127][128] and algorithms[129] have been developed to enhance data interpretation. The existing knowledge base–driven pathway analysis methods in each generation have been summarized in recent literature.[130]
Applications of pathway analysis in medicine
Colorectal cancer (CRC)
A program package MatchMiner was used to scan HUGO names for cloned genes of interest are scanned, then are input into GoMiner, which leveraged the GO to identify the biological processes, functions and components represented in the gene profile. Also, Database for Annotation, Visualization, and Integrated Discovery (DAVID) and KEGG database can be used for the analysis of microarray expression data and the analysis of each GO biological process (P), cellular component (C), and molecular function (F) ontology. In addition, DAVID tools can be used to analyze the roles of genes in metabolic pathways and show the biological relationships between genes or gene-products and may represent metabolic pathways. These two databases also provide bioinformatics tools online to combine specific biochemical information on a certain organism and facilitate the interpretation of biological meanings for experimental data. By using a combined approach of Microarray-Bioinformatic technologies, a potential metabolic mechanism contributing to colorectal cancer (CRC) has been demonstrated[131] Several environmental factors may be involved in a series of points along the genetic pathway to CRC. These include genes associated with bile acid metabolism, glycolysis metabolism and fatty acid metabolism pathways, supporting a hypothesis that some metabolic alternations observed in colon carcinoma may occur in the development of CRC.[131]
Parkinson's disease (PD)
Cellular models are instrumental in dissecting a complex pathological process into simpler molecular events. Parkinson's disease (PD) is multifactorial and clinically heterogeneous; the aetiology of the sporadic (and most common) form is still unclear and only a few molecular mechanisms have been clarified so far in the neurodegenerative cascade. In such a multifaceted picture, it is particularly important to identify experimental models that simplify the study of the different networks of proteins and genes involved. Cellular models that reproduce some of the features of the neurons that degenerate in PD have contributed to many advances in our comprehension of the pathogenic flow of the disease. In particular, the pivotal biochemical pathways (i.e. apoptosis and oxidative stress, mitochondrial impairment and dysfunctional mitophagy, unfolded protein stress and improper removal of misfolded proteins) have been widely explored in cell lines, challenged with toxic insults or genetically modified. The central role of a-synuclein has generated many models aiming to elucidate its contribution to the dysregulation of various cellular processes. Classical cellular models appear to be the correct choice for preliminary studies on the molecular action of new drugs or potential toxins and for understanding the role of single genetic factors. Moreover, the availability of novel cellular systems, such as cybrids or induced pluripotent stem cells, offers the chance to exploit the advantages of an in vitro investigation, although mirroring more closely the cell population being affected.[132]
Alzheimer's disease (AD)
Synaptic degeneration and death of nerve cells are defining features of Alzheimer's disease (AD), the most prevalent age-related neurodegenerative disorders. In AD, neurons in the hippocampus and basal forebrain (brain regions that subserve learning and memory functions) are selectively vulnerable. Studies of postmortem brain tissue from AD people have provided evidence for increased levels of oxidative stress, mitochondrial dysfunction and impaired glucose uptake in vulnerable neuronal populations. Studies of animal and cell culture models of AD suggest that increased levels of oxidative stress (membrane lipid peroxidation, in particular) may disrupt neuronal energy metabolism and ion homeostasis, by impairing the function of membrane ion-motive ATPases, glucose and glutamate transporters. Such oxidative and metabolic compromise may thereby render neurons vulnerable to excitotoxicity and apoptosis. Recent studies suggest that AD can manifest systemic alterations in energy metabolism (e.g., increased insulin resistance and dysregulation of glucose metabolism). Emerging evidence that dietary restriction can forestall the development of AD is consistent with a major "metabolic" component to these disorders, and provides optimism that these devastating brain disorders of aging may be largely preventable.[133]
References
- Bastien D. Gomperts; Peter E.R. Tatham; Ijsbrand M. Kramer (2004). Signal transduction (Pbk. ed., [Nachdr.]. ed.). Amsterdam [u.a.]: Elsevier Academic Press. ISBN 978-0122896323.
- Ingham, P.W.; Nakano, Y.; Seger, C. (2011). "Mechanisms and functions of Hedgehog signalling across the metazoa". Nature Reviews Genetics. 12 (6): 393–406. doi:10.1038/nrg2984. PMID 21502959. S2CID 33769324.
- Antoniotti, M., Park, F., Policriti, A., Ugel, N., Mishra, B. (2003) Foundations of a query and simulation system for the modeling of biochemical and biological processes. In Pacific Symposium on Biocomputing 2003 (PSB 2003), pp. 116–127.
- Fardilha, Margarida (2012). O eSsencial em… Sinalização Celular. Edições Afrontamento. ISBN 9789723612530.
- Jeremy M. Berg; John L. Tymoczko; Lubert Stryer (2007). Biochemistry (6. ed., 3. print. ed.). New York: Freeman. ISBN 978-0716787242.
- Mishra, B. (2002) A symbolic approach to modelling cellular behaviour. In Prasanna, V., Sahni, S. and Shukla, U. (eds), High Performance Computing—HiPC 2002. LNCS 2552. Springer-Verlag, pp. 725–732.
- de Jong, H. (2002) Modeling and simulation of genetic regulatory systems: a literature review. J. Comput. Biol., 9(1), 67–103.
- Hinkle JL, Bowman L (2003) Neuroprotection for ischemic stroke. J Neurosci Nurs 35 (2): 114–8.
- Carneiro, Luiz Carlos; Junqueira, José (2005). Basic histology text & atlas (11th ed.). New York, N.Y., [etc.]: McGraw-Hill. ISBN 978-0071440912.
- Tian, Xinrui; Liu, Z; Niu, B; Zhang, J; Tan, T. K.; Lee, S. R.; Zhao, Y; Harris, D. C.; Zheng, G (2011). "E-Cadherin/β-Catenin Complex and the Epithelial Barrier". Journal of Biomedicine and Biotechnology. 2011: 1–6. doi:10.1155/2011/567305. PMC 3191826. PMID 22007144.
- Barth, Angela IM; Näthke, Inke S; Nelson, W James (October 1997). "Cadherins, catenins and APC protein: interplay between cytoskeletal complexes and signaling pathways". Current Opinion in Cell Biology. 9 (5): 683–690. doi:10.1016/S0955-0674(97)80122-6. PMID 9330872.
- Conacci-Sorrell, Maralice; Zhurinsky, Jacob; Ben-Ze'ev, Avri (15 April 2002). "The cadherin-catenin adhesion system in signaling and cancer". Journal of Clinical Investigation. 109 (8): 987–991. doi:10.1172/JCI15429. PMC 150951. PMID 11956233.
- Gilcrease, Michael Z. (March 2007). "Integrin signaling in epithelial cells". Cancer Letters. 247 (1): 1–25. doi:10.1016/j.canlet.2006.03.031. PMID 16725254.
- Campbell, I. D.; Humphries, M. J. (19 January 2011). "Integrin Structure, Activation, and Interactions". Cold Spring Harbor Perspectives in Biology. 3 (3): a004994. doi:10.1101/cshperspect.a004994. PMC 3039929. PMID 21421922.
- Eugene R. Schiff; Willis C. Maddrey; Michael F. Sorrell, eds. (12 December 2011). Schiff's diseases of the liver (11th ed.). Chichester, West Sussex, UK: John Wiley & Sons. ISBN 978-0-470-65468-2.
- Pawlina, Michael H. Ross, Wojciech (23 April 2011). Histology : a text and atlas : with correlated cell and molecular biology (6th ed.). Philadelphia: Wolters Kluwer/Lippincott Williams & Wilkins Health. ISBN 978-0781772006.
{{cite book}}: CS1 maint: multiple names: authors list (link) - Berridge, Michael J. (10 April 2012). "Cell Signalling Biology: Module 1 - Introduction". Biochemical Journal. 6: csb0001001. doi:10.1042/csb0001001.
- Bode, Johannes G.; Albrecht, Ute; Häussinger, Dieter; Heinrich, Peter C.; Schaper, Fred (June 2012). "Hepatic acute phase proteins – Regulation by IL-6- and IL-1-type cytokines involving STAT3 and its crosstalk with NF-κB-dependent signaling". European Journal of Cell Biology. 91 (6–7): 496–505. doi:10.1016/j.ejcb.2011.09.008. PMID 22093287.
- Wang, Hua (2011). "Signal Transducer and Activator of Transcription 3 in Liver Diseases: A Novel Therapeutic Target". International Journal of Biological Sciences. 7 (5): 536–550. doi:10.7150/ijbs.7.536. PMC 3088876. PMID 21552420.
- Irwin M. Arias; Harvey J. Alter (2009). The liver : biology and pathobiology (5th ed.). Chichester, UK: Wiley-Blackwell. ISBN 978-0470723135.
- Tolosano, Emanuela; Altruda, Fiorella (April 2002). "Hemopexin: Structure, Function, and Regulation". DNA and Cell Biology. 21 (4): 297–306. doi:10.1089/104454902753759717. PMID 12042069.
- Jean-Francois Dufour; Pierre-Alain Clavien (2010). Signaling pathways in liver diseases (2. ed.). Berlin: Springer. ISBN 978-3-642-00149-9.
- Edwards, Peter A; Kennedy, Matthew A; Mak, Puiying A (April 2002). "LXRs;". Vascular Pharmacology. 38 (4): 249–256. doi:10.1016/S1537-1891(02)00175-1. PMID 12449021.
- Dzau, VJ; Herrmann, HC (15–22 February 1982). "Hormonal control of angiotensinogen production". Life Sciences. 30 (7–8): 577–84. doi:10.1016/0024-3205(82)90272-7. PMID 7040893.
- Chi, Hsiang Cheng; Chen, Cheng-Yi; Tsai, Ming-Ming; Tsai, Chung-Ying; Lin, Kwang-Huei (2013). Tsai, Ming-Ming; Tsai, Chung-Ying; Lin, Kwang-Huei. "Molecular Functions of Thyroid Hormones and Their Clinical Significance in Liver-Related Diseases". BioMed Research International. 2013: 1–16. doi:10.1155/2013/601361. PMC 3708403. PMID 23878812.
- Lai, Hong-Shiee; Lin, Wen-Hsi (3 July 2013). Lai, Shuo-Lun; Lin, Hao-Yu; Hsu, Wen-Ming; Chou, Chia-Hung; Lee, Po-Huang; Rishi, Arun. "Interleukin-6 Mediates Angiotensinogen Gene Expression during Liver Regeneration". PLOS ONE. 8 (7): e67868. Bibcode:2013PLoSO...867868L. doi:10.1371/journal.pone.0067868. PMC 3700864. PMID 23844114.
- Nakamura, T; Mizuno, S (2010). "The discovery of hepatocyte growth factor (HGF) and its significance for cell biology, life sciences and clinical medicine". Proceedings of the Japan Academy, Series B. 86 (6): 588–610. Bibcode:2010PJAB...86..588N. doi:10.2183/pjab.86.588. PMC 3081175. PMID 20551596.
- Blumenschein GR, Jr; Mills, GB; Gonzalez-Angulo, AM (10 September 2012). "Targeting the hepatocyte growth factor-cMET axis in cancer therapy". Journal of Clinical Oncology. 30 (26): 3287–96. doi:10.1200/JCO.2011.40.3774. PMC 3434988. PMID 22869872.
- Organ, SL; Tsao, MS (November 2011). "An overview of the c-MET signaling pathway". Therapeutic Advances in Medical Oncology. 3 (1 Suppl): S7–S19. doi:10.1177/1758834011422556. PMC 3225017. PMID 22128289.
- Dufour, Jean-François (2005). Signaling pathways in liver diseases : with 15 tables. Berlin [u.a.]: Springer. ISBN 978-3540229346.
- Fields, RD; Burnstock, G (June 2006). "Purinergic signalling in neuron-glia interactions". Nature Reviews Neuroscience. 7 (6): 423–36. doi:10.1038/nrn1928. PMC 2062484. PMID 16715052.
- Abbracchio, Maria P.; Burnstock, Geoffrey; Verkhratsky, Alexei; Zimmermann, Herbert (January 2009). "Purinergic signalling in the nervous system: an overview". Trends in Neurosciences. 32 (1): 19–29. doi:10.1016/j.tins.2008.10.001. PMID 19008000. S2CID 7653609.
- Vargas, MR; Johnson, JA (3 June 2009). "The Nrf2-ARE cytoprotective pathway in astrocytes". Expert Reviews in Molecular Medicine. 11: e17. doi:10.1017/S1462399409001094. PMC 5563256. PMID 19490732.
- Habas, A.; Hahn, J.; Wang, X.; Margeta, M. (21 October 2013). "Neuronal activity regulates astrocytic Nrf2 signaling". Proceedings of the National Academy of Sciences. 110 (45): 18291–18296. Bibcode:2013PNAS..11018291H. doi:10.1073/pnas.1208764110. PMC 3831500. PMID 24145448.
- Escartin, C; Won, SJ (18 May 2011). Malgorn, C; Auregan, G; Berman, AE; Chen, PC; Déglon, N; Johnson, JA; Suh, SW; Swanson, RA. "Nuclear factor erythroid 2-related factor 2 facilitates neuronal glutathione synthesis by upregulating neuronal excitatory amino acid transporter 3 expression". The Journal of Neuroscience. 31 (20): 7392–401. doi:10.1523/JNEUROSCI.6577-10.2011. PMC 3339848. PMID 21593323.
- Johnson, JA; Johnson, DA; Kraft, A. D.; Calkins, M. J.; Jakel, R. J.; Vargas, M. R.; Chen, P. C. (December 2008). Kraft, AD; Calkins, MJ; Jakel, RJ; Vargas, MR; Chen, PC. "The Nrf2-ARE pathway: an indicator and modulator of oxidative stress in neurodegeneration". Annals of the New York Academy of Sciences. 1147: 61–9. doi:10.1196/annals.1427.036. PMC 2605641. PMID 19076431.
- Lewis, T. L.; Courchet, J.; Polleux, F. (16 September 2013). "Cell biology in neuroscience: Cellular and molecular mechanisms underlying axon formation, growth, and branching". The Journal of Cell Biology. 202 (6): 837–848. doi:10.1083/jcb.201305098. PMC 3776347. PMID 24043699.
- Courchet, Julien; Lewis, Tommy L. (June 2013). Lee, Sohyon; Courchet, Virginie; Liou, Deng-Yuan; Aizawa, Shinichi; Polleux, Franck. "Terminal Axon Branching Is Regulated by the LKB1-NUAK1 Kinase Pathway via Presynaptic Mitochondrial Capture". Cell. 153 (7): 1510–1525. doi:10.1016/j.cell.2013.05.021. PMC 3729210. PMID 23791179.
- Satoh, Daisuke; Arber, Silvia (June 2013). "Carving Axon Arbors to Fit: Master Directs One Kinase at a Time". Cell. 153 (7): 1425–1426. doi:10.1016/j.cell.2013.05.047. PMID 23791171.
- Ellsworth, ML; Ellis, CG; Goldman, D; Stephenson, A. H.; Dietrich, H. H.; Sprague, R. S. (April 2009). Goldman, D; Stephenson, AH; Dietrich, HH; Sprague, RS. "Erythrocytes: oxygen sensors and modulators of vascular tone". Physiology. 24 (2): 107–16. doi:10.1152/physiol.00038.2008. PMC 2725440. PMID 19364913.
- Sprague, RS; Ellsworth, ML (July 2012). "Erythrocyte-derived ATP and perfusion distribution: role of intracellular and intercellular communication". Microcirculation. 19 (5): 430–9. doi:10.1111/j.1549-8719.2011.00158.x. PMC 3324633. PMID 22775760.
- Ley, K; Laudanna, C; Cybulsky, MI; Nourshargh, S (September 2007). "Getting to the site of inflammation: the leukocyte adhesion cascade updated". Nature Reviews. Immunology. 7 (9): 678–89. doi:10.1038/nri2156. PMID 17717539. S2CID 1871230.
- Nourshargh, S; Hordijk, PL; Sixt, M (May 2010). "Breaching multiple barriers: leukocyte motility through venular walls and the interstitium". Nature Reviews Molecular Cell Biology. 11 (5): 366–78. doi:10.1038/nrm2889. PMID 20414258. S2CID 9669661.
- Roitt, Ivan M (2013). Fundamentos de Imunologia. GUANABARA KOOGAN. ISBN 978-8527721424.
- Baker, Abul (2012). Cellular and molecular immunology. K. Abbas, Andrew H. Lichtman, Shiv Pillai; illustrations by David L. Baker, Alexandra (7th ed.). Philadelphia: Elsevier/Saunders. ISBN 978-1437715286.
- Cox, Michael (2005). Encyclopedia of life sciences. Hoboken, NJ [u.a.]: Wiley [Online-Anbieter]. ISBN 9780470015902.
- Macian, F (June 2005). "NFAT proteins: key regulators of T-cell development and function". Nature Reviews. Immunology. 5 (6): 472–84. doi:10.1038/nri1632. PMID 15928679. S2CID 2460785.
- Mercedes Rincón; Richard A Flavell & Roger J Davis (2001). "Signal transduction by MAP kinases in T lymphocytes". Oncogene. 20 (19): 2490–2497. doi:10.1038/sj.onc.1204382. PMID 11402343.
- Weiss, Arthur. "Signal Transduction Events Involved in Lymphocyte Activation and Differentiation". Retrieved 8 January 2014.
- Le Gallou, S; Caron, G (1 July 2012). Delaloy, C; Rossille, D; Tarte, K; Fest, T. "IL-2 requirement for human plasma cell generation: coupling differentiation and proliferation by enhancing MAPK-ERK signaling". Journal of Immunology. 189 (1): 161–73. doi:10.4049/jimmunol.1200301. PMID 22634617.
- Shaffer, AL; Shapiro-Shelef, M (July 2004). Iwakoshi, NN; Lee, AH; Qian, SB; Zhao, H; Yu, X; Yang, L; Tan, BK; Rosenwald, A; Hurt, EM; Petroulakis, E; Sonenberg, N; Yewdell, JW; Calame, K; Glimcher, LH; Staudt, LM. "XBP1, downstream of Blimp-1, expands the secretory apparatus and other organelles, and increases protein synthesis in plasma cell differentiation". Immunity. 21 (1): 81–93. doi:10.1016/j.immuni.2004.06.010. PMID 15345222.
- Crotty, Shane; Johnston, Robert J; Schoenberger, Stephen P (19 January 2010). "Effectors and memories: Bcl-6 and Blimp-1 in T and B lymphocyte differentiation". Nature Immunology. 11 (2): 114–120. doi:10.1038/ni.1837. PMC 2864556. PMID 20084069.
- Michael Cox (2005). Encyclopedia of life sciences. Hoboken, NJ [u.a.]: Wiley [Online-Anbieter]. ISBN 9780470015902.
- Cruciat, CM.; Niehrs, C. (19 October 2012). "Secreted and Transmembrane Wnt Inhibitors and Activators". Cold Spring Harbor Perspectives in Biology. 5 (3): a015081. doi:10.1101/cshperspect.a015081. PMC 3578365. PMID 23085770.
- Kobayashi, Yasuhiro; Maeda, Kazuhiro; Takahashi, Naoyuki (July 2008). "Roles of Wnt signaling in bone formation and resorption". Japanese Dental Science Review. 44 (1): 76–82. doi:10.1016/j.jdsr.2007.11.002.
- Raju, R; Balakrishnan, L; Nanjappa, V; Bhattacharjee, M; Getnet, D; Muthusamy, B; Kurian Thomas, J; Sharma, J; Rahiman, B. A.; Harsha, H. C.; Shankar, S; Prasad, T. S.; Mohan, S. S.; Bader, G. D.; Wani, M. R.; Pandey, A (2011). "A comprehensive manually curated reaction map of RANKL/RANK-signaling pathway". Database (Oxford). 2011: bar021. doi:10.1093/database/bar021. PMC 3170171. PMID 21742767.
- Boyce, BF; Xing, L (2007). "Biology of RANK, RANKL, and osteoprotegerin". Arthritis Research & Therapy. 9 (Suppl 1): S1. doi:10.1186/ar2165. PMC 1924516. PMID 17634140.
- Mediero, Aránzazu; Cronstein, Bruce N. (June 2013). "Adenosine and bone metabolism". Trends in Endocrinology & Metabolism. 24 (6): 290–300. doi:10.1016/j.tem.2013.02.001. PMC 3669669. PMID 23499155.
- Ham, J; Evans, BA (2012). "An emerging role for adenosine and its receptors in bone homeostasis". Frontiers in Endocrinology. 3: 113. doi:10.3389/fendo.2012.00113. PMC 3444801. PMID 23024635.
- Watt, F. M.; Driskell, R. R. (24 November 2009). "The therapeutic potential of stem cells". Philosophical Transactions of the Royal Society B: Biological Sciences. 365 (1537): 155–163. doi:10.1098/rstb.2009.0149. PMC 2842697. PMID 20008393.
- Ying, QL; Nichols, J; Chambers, I; Smith, A (31 October 2003). "BMP induction of Id proteins suppresses differentiation and sustains embryonic stem cell self-renewal in collaboration with STAT3". Cell. 115 (3): 281–92. doi:10.1016/S0092-8674(03)00847-X. PMID 14636556. S2CID 7201396.
- Nishino, J; Kim, I; Chada, K; Morrison, SJ (17 October 2008). "Hmga2 promotes neural stem cell self-renewal in young but not old mice by reducing p16Ink4a and p19Arf Expression". Cell. 135 (2): 227–39. doi:10.1016/j.cell.2008.09.017. PMC 2582221. PMID 18957199.
- Morrison, SJ; Spradling, AC (22 February 2008). "Stem cells and niches: mechanisms that promote stem cell maintenance throughout life". Cell. 132 (4): 598–611. doi:10.1016/j.cell.2008.01.038. PMC 4505728. PMID 18295578.
- Fuchs, E; Tumbar, T; Guasch, G (19 March 2004). "Socializing with the neighbors: stem cells and their niche". Cell. 116 (6): 769–78. doi:10.1016/s0092-8674(04)00255-7. PMID 15035980. S2CID 18494303.
- Clarke, MF; Dick, JE (1 October 2006). Dirks, PB; Eaves, CJ; Jamieson, CH; Jones, DL; Visvader, J; Weissman, IL; Wahl, GM. "Cancer stem cells--perspectives on current status and future directions: AACR Workshop on cancer stem cells". Cancer Research. 66 (19): 9339–44. doi:10.1158/0008-5472.CAN-06-3126. PMID 16990346. S2CID 8791540.
- Jones, GM; Cram, DS (May 2008). Song, B; Magli, MC; Gianaroli, L; Lacham-Kaplan, O; Findlay, JK; Jenkin, G; Trounson, AO. "Gene expression profiling of human oocytes following in vivo or in vitro maturation". Human Reproduction. 23 (5): 1138–44. doi:10.1093/humrep/den085. PMID 18346995.
- Kidder, GM; Vanderhyden, BC (April 2010). "Bidirectional communication between oocytes and follicle cells: ensuring oocyte developmental competence". Canadian Journal of Physiology and Pharmacology. 88 (4): 399–413. doi:10.1139/y10-009. PMC 3025001. PMID 20555408.
- Peng, J.; Li, Q. (4 February 2013). Wigglesworth, K.; Rangarajan, A.; Kattamuri, C.; Peterson, R. T.; Eppig, J. J.; Thompson, T. B.; Matzuk, M. M. "Growth differentiation factor 9:bone morphogenetic protein 15 heterodimers are potent regulators of ovarian functions". Proceedings of the National Academy of Sciences. 110 (8): E776–E785. doi:10.1073/pnas.1218020110. PMC 3581982. PMID 23382188.
- McGinnis, LK; Carroll, DJ; Kinsey, WH (October–November 2011). "Protein tyrosine kinase signaling during oocyte maturation and fertilization". Molecular Reproduction and Development. 78 (10–11): 831–45. doi:10.1002/mrd.21326. PMC 3186829. PMID 21681843.
- Norris, RP; Ratzan, WJ (June 2009). Freudzon, M; Mehlmann, LM; Krall, J; Movsesian, MA; Wang, H; Ke, H; Nikolaev, VO; Jaffe, LA. "Cyclic GMP from the surrounding somatic cells regulates cyclic AMP and meiosis in the mouse oocyte". Development. 136 (11): 1869–78. doi:10.1242/dev.035238. PMC 2680110. PMID 19429786.
- Vaccari, S; Weeks JL, 2nd (September 2009). Hsieh, M; Menniti, FS; Conti, M. "Cyclic GMP signaling is involved in the luteinizing hormone-dependent meiotic maturation of mouse oocytes". Biology of Reproduction. 81 (3): 595–604. doi:10.1095/biolreprod.109.077768. PMC 2731981. PMID 19474061.
- Sela-Abramovich, S; Edry, I; Galiani, D; Nevo, N; Dekel, N (May 2006). "Disruption of gap junctional communication within the ovarian follicle induces oocyte maturation". Endocrinology. 147 (5): 2280–6. doi:10.1210/en.2005-1011. PMID 16439460.
- Sela-Abramovich, S; Chorev, E; Galiani, D; Dekel, N (March 2005). "Mitogen-activated protein kinase mediates luteinizing hormone-induced breakdown of communication and oocyte maturation in rat ovarian follicles". Endocrinology. 146 (3): 1236–44. doi:10.1210/en.2004-1006. PMID 15576461.
- Kim, J; Bagchi, IC; Bagchi, MK (December 2009). "Control of ovulation in mice by progesterone receptor-regulated gene networks". Molecular Human Reproduction. 15 (12): 821–8. doi:10.1093/molehr/gap082. PMC 2776476. PMID 19815644.
- Fortune, JE; Willis, EL; Bridges, PJ; Yang, CS (January 2009). "The periovulatory period in cattle: progesterone, prostaglandins, oxytocin and ADAMTS proteases". Animal Reproduction. 6 (1): 60–71. PMC 2853051. PMID 20390049.
- Geldziler, BD; Marcello, MR; Shakes, D. C.; Singson, A (2011). The genetics and cell biology of fertilization. Methods in Cell Biology. Vol. 106. pp. 343–75. doi:10.1016/B978-0-12-544172-8.00013-X. ISBN 9780125441728. PMC 3275088. PMID 22118284.
- Han, SM; Cottee, PA; Miller, MA (May 2010). "Sperm and oocyte communication mechanisms controlling C. elegans fertility". Developmental Dynamics. 239 (5): 1265–81. doi:10.1002/dvdy.22202. PMC 2963114. PMID 20034089.
- Miao, YL; Williams, CJ (November 2012). "Calcium signaling in mammalian egg activation and embryo development: the influence of subcellular localization". Molecular Reproduction and Development. 79 (11): 742–56. doi:10.1002/mrd.22078. PMC 3502661. PMID 22888043.
- Swann, K; Windsor, S (March 2012). Campbell, K; Elgmati, K; Nomikos, M; Zernicka-Goetz, M; Amso, N; Lai, FA; Thomas, A; Graham, C. "Phospholipase C-ζ-induced Ca2+ oscillations cause coincident cytoplasmic movements in human oocytes that failed to fertilize after intracytoplasmic sperm injection". Fertility and Sterility. 97 (3): 742–7. doi:10.1016/j.fertnstert.2011.12.013. PMC 3334266. PMID 22217962.
- Mio, Y; Iwata, K (September 2012). Yumoto, K; Kai, Y; Sargant, HC; Mizoguchi, C; Ueda, M; Tsuchie, Y; Imajo, A; Iba, Y; Nishikori, K. "Possible mechanism of polyspermy block in human oocytes observed by time-lapse cinematography". Journal of Assisted Reproduction and Genetics. 29 (9): 951–6. doi:10.1007/s10815-012-9815-x. PMC 3463667. PMID 22695746.
- Beall, S; Brenner, C; Segars, J (December 2010). "Oocyte maturation failure: a syndrome of bad eggs". Fertility and Sterility. 94 (7): 2507–13. doi:10.1016/j.fertnstert.2010.02.037. PMC 2946974. PMID 20378111.
- Abou-haila, A; Tulsiani, DR (1 May 2009). "Signal transduction pathways that regulate sperm capacitation and the acrosome reaction". Archives of Biochemistry and Biophysics. 485 (1): 72–81. doi:10.1016/j.abb.2009.02.003. PMID 19217882.
- Visconti, PE; Westbrook, VA (January 2002). Chertihin, O; Demarco, I; Sleight, S; Diekman, AB. "Novel signaling pathways involved in sperm acquisition of fertilizing capacity". Journal of Reproductive Immunology. 53 (1–2): 133–50. doi:10.1016/S0165-0378(01)00103-6. PMID 11730911.
- Salicioni, AM; Platt, MD; Wertheimer, E. V.; Arcelay, E; Allaire, A; Sosnik, J; Visconti, P. E. (2007). Wertheimer, EV; Arcelay, E; Allaire, A; Sosnik, J; Visconti, PE. "Signalling pathways involved in sperm capacitation". Society of Reproduction and Fertility Supplement. 65: 245–59. PMID 17644966.
- Breitbart, H (22 February 2002). "Intracellular calcium regulation in sperm capacitation and acrosomal reaction". Molecular and Cellular Endocrinology. 187 (1–2): 139–44. doi:10.1016/s0303-7207(01)00704-3. PMID 11988321. S2CID 24124381.
- Gupta, SK; Bhandari, B (January 2011). "Acrosome reaction: relevance of zona pellucida glycoproteins". Asian Journal of Andrology. 13 (1): 97–105. doi:10.1038/aja.2010.72. PMC 3739397. PMID 21042299.
- Sagare-Patil, V; Vernekar, M; Galvankar, M; Modi, D (15 July 2013). "Progesterone utilizes the PI3K-AKT pathway in human spermatozoa to regulate motility and hyperactivation but not acrosome reaction". Molecular and Cellular Endocrinology. 374 (1–2): 82–91. doi:10.1016/j.mce.2013.04.005. PMID 23623968. S2CID 25689637.
- Publicover, S; Barratt, C (17 March 2011). "Reproductive biology: Progesterone's gateway into sperm". Nature. 471 (7338): 313–4. Bibcode:2011Natur.471..313P. doi:10.1038/471313a. PMID 21412330. S2CID 205062974.
- Ashok Agarwal; R. John Aitken; Juan G. Alvarez (17 March 2012). Studies on men's health and fertility. New York: Humana Press. ISBN 978-1-61779-775-0.
- O'Flaherty, C; de Lamirande, E; Gagnon, C (15 August 2006). "Positive role of reactive oxygen species in mammalian sperm capacitation: triggering and modulation of phosphorylation events". Free Radical Biology & Medicine. 41 (4): 528–40. doi:10.1016/j.freeradbiomed.2006.04.027. PMID 16863985.
- Dorey, K; Amaya, E (November 2010). "FGF signalling: diverse roles during early vertebrate embryogenesis". Development. 137 (22): 3731–42. doi:10.1242/dev.037689. PMC 3747497. PMID 20978071.
- Lanner, F; Rossant, J (October 2010). "The role of FGF/Erk signaling in pluripotent cells". Development. 137 (20): 3351–60. doi:10.1242/dev.050146. PMID 20876656. S2CID 1380227.
- Dreesen, O; Brivanlou, AH (January 2007). "Signaling pathways in cancer and embryonic stem cells". Stem Cell Reviews. 3 (1): 7–17. doi:10.1007/s12015-007-0004-8. PMID 17873377. S2CID 25311665.
- Li, J; Wang, G (April 2007). Wang, C; Zhao, Y; Zhang, H; Tan, Z; Song, Z; Ding, M; Deng, H. "MEK/ERK signaling contributes to the maintenance of human embryonic stem cell self-renewal". Differentiation; Research in Biological Diversity. 75 (4): 299–307. doi:10.1111/j.1432-0436.2006.00143.x. PMID 17286604.
- Sui, Lina; Bouwens, Luc; Mfopou, Josué K. (2013). "Signaling pathways during maintenance and definitive endoderm differentiation of embryonic stem cells". The International Journal of Developmental Biology. 57 (1): 1–12. doi:10.1387/ijdb.120115ls. PMID 23585347. S2CID 38544740.
- Manning, BD; Cantley, LC (29 June 2007). "AKT/PKB signaling: navigating downstream". Cell. 129 (7): 1261–74. doi:10.1016/j.cell.2007.06.009. PMC 2756685. PMID 17604717.
- Song, G; Ouyang, G; Bao, S (January–March 2005). "The activation of Akt/PKB signaling pathway and cell survival". Journal of Cellular and Molecular Medicine. 9 (1): 59–71. doi:10.1111/j.1582-4934.2005.tb00337.x. PMC 6741304. PMID 15784165.
- Dailey, L; Ambrosetti, D; Mansukhani, A; Basilico, C (April 2005). "Mechanisms underlying differential responses to FGF signaling". Cytokine & Growth Factor Reviews. 16 (2): 233–47. doi:10.1016/j.cytogfr.2005.01.007. PMID 15863038.
- Kelleher, FC; Fennelly, D; Rafferty, M (2006). "Common critical pathways in embryogenesis and cancer". Acta Oncologica. 45 (4): 375–88. doi:10.1080/02841860600602946. PMID 16760173. S2CID 24282171.
- Wang, J; Wynshaw-Boris, A (October 2004). "The canonical Wnt pathway in early mammalian embryogenesis and stem cell maintenance/differentiation". Current Opinion in Genetics & Development. 14 (5): 533–9. doi:10.1016/j.gde.2004.07.013. PMID 15380245.
- Wu, MY; Hill, CS (March 2009). "Tgf-beta superfamily signaling in embryonic development and homeostasis". Developmental Cell. 16 (3): 329–43. doi:10.1016/j.devcel.2009.02.012. PMID 19289080.
- Kishigami, S; Mishina, Y (June 2005). "BMP signaling and early embryonic patterning". Cytokine & Growth Factor Reviews. 16 (3): 265–78. doi:10.1016/j.cytogfr.2005.04.002. PMID 15871922.
- Lifantseva, N. V.; Koltsova, A. M.; Poljanskaya, G. G.; Gordeeva, O. F. (23 January 2013). "Expression of TGFβ family factors and FGF2 in mouse and human embryonic stem cells maintained in different culture systems". Russian Journal of Developmental Biology. 44 (1): 7–18. doi:10.1134/S1062360413010050. PMID 23659078. S2CID 8167222.
- Viswanathan, G. A.; Seto, J.; Patil, S.; Nudelman, G.; Sealfon, S. C. (2008). "Getting Started in Biological Pathway Construction and Analysis". PLOS Comput Biol. 4 (2): e16. Bibcode:2008PLSCB...4...16V. doi:10.1371/journal.pcbi.0040016. PMC 2323403. PMID 18463709.
- Stromback L., Jakoniene V., Tan H., Lambrix P. (2006) Representing, storing and accessing. The MIT Press.
- Brazma, A.; Krestyaninova, M.; Sarkans, U. (2006). "Standards for systems biology". Nat Rev Genet. 7 (8): 593–605. doi:10.1038/nrg1922. PMID 16847461. S2CID 35398897.
- Baclawski K., Niu T. (2006) Ontologies for bioinformatics. Cambridge (Massachusetts): Boca Raton (Florida): Chapman & Hall/CRC.
- Kashtan, N.; Itzkovitz, S.; Milo, R.; Alon, U. (2004). "Efficient sampling algorithm for estimating subgraph concentrations and detecting network motifs". Bioinformatics. 20 (11): 1746–1758. doi:10.1093/bioinformatics/bth163. PMID 15001476.
- "KEGG: Kyoto Encyclopedia of Genes and Genomes".
- Kanehisa, M.; Goto, S.; Hattori, M.; Aoki-Kinoshita, K.F.; Itoh, M.; Kawashima, S. (2006). "From genomics to chemical genomics: new developments in KEGG". Nucleic Acids Res. 34 (Database issue): D354–D357. doi:10.1093/nar/gkj102. PMC 1347464. PMID 16381885.
- Minoru K., Susumu G., Miho F., Mao T., Mika H. (2010) KEGG for representation and analysis of molecular networks involving diseases and drugs Nucleic Acids Res. 38(1): D355-D360.
- "Home". genmapp.org.
- Dahlquist, K. D.; Salomonis, N.; Vranizan, K.; Lawlor, S. C.; Conklin, B. R. (2002). "GenMAPP, a new tool for viewing and analyzing microarray data on biological pathways". Nat. Genet. 31 (1): 19–20. doi:10.1038/ng0502-19. PMID 11984561.
- "Archived copy" (PDF). www.genmapp.org. Archived from the original (PDF) on 3 February 2013. Retrieved 12 January 2022.
{{cite web}}: CS1 maint: archived copy as title (link) - Vastrik, I.; D'Eustachio, P.; Schmidt, E.; Joshi-Tope, G.; Gopinath, G.; Croft, D.; de Bono, B.; Gillespie, M.; Jassal, B.; Lewis, S.; Matthews, L.; Wu, G.; Birney, E.; Stein, L. (2007). "Reactome: a knowledgebase of biological pathways and processes". Genome Biol. 8 (3): R39. doi:10.1186/gb-2007-8-3-r39. PMC 1868929. PMID 17367534.
- Joshi-Tope, G.; Gillespie, M.; Vastrik, I.; D'Eustachio, P.; Schmidt, E.; de Bono, B.; Jassal, B.; Gopinath, G. R.; Wu, G. R.; Matthews, L.; Lewis, S.; Birney, E.; Stein, L. (2005). "Reactome: a knowledgebase of biological pathways". Nucleic Acids Res. 33 (Database issue): D428–32. doi:10.1093/nar/gki072. PMC 540026. PMID 15608231.
- Matthews, L.; Gopinath, G.; Gillespie, M.; Caudy, M. (2009). "Reactome knowledge base of human biological pathways and processes". Nucleic Acids Res. 37 (Database issue): D619–D622. doi:10.1093/nar/gkn863. PMC 2686536. PMID 18981052.
- Croft, D.; O'Kelly, G.; Wu, G.; Haw, R. (2011). "Reactome: a database of reactions, pathways and biological processes". Nucleic Acids Res. 39 (Database issue): D691–D697. doi:10.1093/nar/gkq1018. PMC 3013646. PMID 21067998.
- Haw, R.; Hermjakob, H.; D'Eustachio, P.; Stein, L. (2011). "Reactome pathway analysis to enrich biological discovery in proteomics data sets". Proteomics. 11 (18): 3598–3613. doi:10.1002/pmic.201100066. PMC 4617659. PMID 21751369.
- Priami, C. (ed.) (2003) Computational Methods in Systems Biology. LNCS 2602. Springer Verlag.
- Karp, P. D.; Riley, M.; Saier, M.; Paulsen, I. T.; Paley, S. M.; Pellegrini-Toole, A. (2000). "The ecocyc and metacyc databases". Nucleic Acids Res. 28 (1): 56–59. doi:10.1093/nar/28.1.56. PMC 102475. PMID 10592180.
- Ogata, H.; Goto, S.; Sato, K.; Fujibuchi, W.; Bono, H.; Kanehisa, M. (1999). "Kegg: Kyoto encyclopedia of genes and genomes". Nucleic Acids Res. 27 (1): 29–34. doi:10.1093/nar/27.1.29. PMC 148090. PMID 9847135.
- Ashburner, M (2000). "Gene ontology: tool for the unification of biology. The Gene Ontology Consortium". Nat. Genet. 25 (1): 25–29. doi:10.1038/75556. PMC 3037419. PMID 10802651.
- Kanehisa, M (2002). "The KEGG databases at GenomeNet". Nucleic Acids Res. 30 (1): 42–46. doi:10.1093/nar/30.1.42. PMC 99091. PMID 11752249.
- Boyle, E. I. (2004). "GO::TermFinder–open source software for accessing Gene Ontology information and finding significantly enriched gene ontology terms associated with a list of genes". Bioinformatics. 20 (18): 3710–3715. doi:10.1093/bioinformatics/bth456. PMC 3037731. PMID 15297299.
- Huang, D. W. (2007). "The DAVID Gene Functional Classification Tool: a novel biological module-centric algorithm to functionally analyze large gene lists". Genome Biol. 8 (9): R183. doi:10.1186/gb-2007-8-9-r183. PMC 2375021. PMID 17784955.
- Maere, S (2005). "BiNGO: a Cytoscape plugin to assess overrepresentation of Gene Ontology categories in biological networks". Bioinformatics. 21 (16): 3448–3449. doi:10.1093/bioinformatics/bti551. PMID 15972284.
- Ramos, H (2008). "The protein information and property explorer: an easy-to-use, rich-client web application for the management and functional analysis of proteomic data". Bioinformatics. 24 (18): 2110–2111. doi:10.1093/bioinformatics/btn363. PMC 2638980. PMID 18635572.
- Li, Y (2008). "A global pathway crosstalk network". Bioinformatics. 24 (12): 1442–1447. doi:10.1093/bioinformatics/btn200. PMID 18434343.
- Khatri, P.; Sirota, M.; Butte, A. J. (2012). "Ten Years of Pathway Analysis: Current Approaches and Outstanding Challenges". PLOS Comput. Biol. 8 (2): e1002375. Bibcode:2012PLSCB...8E2375K. doi:10.1371/journal.pcbi.1002375. PMC 3285573. PMID 22383865.
- Yeh, C. S.; Wang, J. Y.; Cheng, T. L.; Juan, C. H.; Wu, C. H.; Lin, S. R. (2006). "Fatty acid metabolism pathway play an important role in carcinogenesis of human colorectal cancers by Microarray-Bioinformatics analysis". Cancer Letters. 233 (2): 297–308. doi:10.1016/j.canlet.2005.03.050. PMID 15885896.
- Alberio, T.; Lopiano, L.; Fasano, M. (2012). "Cellular models to investigate biochemical pathways in Parkinson's disease". FEBS Journal. 279 (7): 1146–1155. doi:10.1111/j.1742-4658.2012.08516.x. PMID 22314200. S2CID 22244998.
- Mattson, M. P.; Pedersen, W. A.; Duan, W.; Culmsee, C.; Camandola, S. (1999). "Cellular and Molecular Mechanisms Underlying Perturbed Energy Metabolism and Neuronal Degeneration in Alzheimer's and Parkinson's Diseases". Annals of the New York Academy of Sciences (Submitted manuscript). 893 (1): 154–175. Bibcode:1999NYASA.893..154M. doi:10.1111/j.1749-6632.1999.tb07824.x. PMID 10672236. S2CID 23438312.
External links
- KEGG resource
- DAVID tools Archived 16 September 2014 at the Wayback Machine
- GenMAPP
- GoMiner Archived 1 January 2016 at the Wayback Machine
- Pathguide