Synapsin I
Synapsin I, is the collective name for Synapsin Ia and Synapsin Ib, two nearly identical phosphoproteins that in humans are encoded by the SYN1 gene.[5][6] In its phosphorylated form, Synapsin I may also be referred to as phosphosynaspin I. Synapsin I is the first of the proteins in the synapsin family of phosphoproteins in the synaptic vesicles present in the central and peripheral nervous systems. Synapsin Ia and Ib are close in length and almost the same in make up, however, Synapsin Ib stops short of the last segment of the C-terminal in the amino acid sequence found in Synapsin Ia.

Protein
The synapsin I protein is a member of the synapsin family that are neuronal phosphoproteins which associate with the cytoplasmic surface of synaptic vesicles. Family members are characterized by common protein domains, and they are implicated in synaptogenesis and the modulation of neurotransmitter release, suggesting a potential role in several neuropsychiatric diseases.
The phosphoprotein plays a role in regulation of axonogenesis and synaptogenesis. The protein serves as a substrate for several different protein kinases and phosphorylation may function in the regulation of this protein in the nerve terminal.[6]
Synapsin I is found in two isoforms of the protein, Synapsin Ia and Synapsin Ib, with Synapsin Ib being a slightly shorter version of the protein. Both Synapsin I proteins are highly basic with a pI in the range of 10.3 and 10.2, respectively. Both isoforms are phosphorylated at identical locations within their protein sequences at the same three serine residues.
Synapsin I phosphoproteins make up approximately 6% of the total protein in synaptic vesicles.[7] Among bovine, rat, and human it has been shown to be 95% homologous, with the central 'C' domain evolutionarily conserved. This phosphoprotein is loosely associated with the vesicular membrance and is easily dissociated by treatment with a salt, versus a detergent being required for its removal from the membrane.
Structure
Synapsin I proteins are made up of a globular portion at the N-terminal and an elongated C-terminal domain, rendering them largely elongated. Synapsin Ib has the same protein domains as synapsin Ia, however synapsin Ib lacks the last C-terminal segment, making it slightly shorter in its elongated domain. 706 amino acids comprise synapsin Ia, and starting from the N-terminal, the same first 670 amino acids comprise synapsin Ib.
Rich in the amino acids proline and glycine, the compositional and structural natures of this protein are somewhat similar to collagen. This aided in the early determination of its structure using collagenase, which was later confirmed by amino acid sequencing and modern techniques. Cleavage of synapsin I by collagenase fragments the elongated C-terminal and leaves the globular N-terminal domain intact.[8]
Amino acid sequencing has shown that synapsin I has common N-terminals across both isoforms and shares the same N-terminal as synapsin II. Synapsin I isoforms differ from synapsin II isoforms in their C-terminal domains as well.[9] Further research has been done on the interactions of synapsin I, synapsin II, and synapsin III with each other to create heterodimers of the proteins in COS cells.[10]
Function
Synapsin I is present in the nerve terminal of axons, specifically in the membranes of synaptic vesicles based on immunocytochemistry.[11] This phosphoprotein is as an endogenous substrate bound to the vesicular membrane. It is phosphorylated by four known classes of protein kinases including those activated by cAMP,[12][13] calcium/calmodulin,[14] mitogen, and cyclin. Both isoforms have the same six phosphorylation sites:
The N-terminal globular domain contains three sites: the cAMP-dependent protein kinase-mediated phosphorylation site near the end in domain A, and two sites further in, in domain B, mediated by mitogen-activated protein kinase (MAP kinase). The tail portion of the protein, the C-terminal end, bears three phosphorylation sites: two sites at which calcium/calmodulin dependent protein kinase II acts, and a third site at which MAP kinase and cyclin-dependent protein kinase (CDK) acts. Specificity for calcium/calmodulin dependent protein kinase binding to Synapsin I is very high in comparison to other substrate proteins.[15] Cyclic AMP-dependent protein kinase is unique in its mechanism of activation. The protein kinase is composed of two regulatory (R) subunits and two catalytic (C) subunits, creating a tetrameric holoenzyme. Cyclic AMP binds to the regulatory subunits of cAMP-dependent protein kinase and causes the dissociation of its regulatory subunits from the catalytic subunits, generating the active form of the kinase. This active form of the protein kinase catalyses the phosphorylation of Synapsin I. The phosphorylated form of Synapsin I is referred to as phosphosynapsin I.
Depolarization of the presynaptic membrane induces a calcium ion influx into the axonal nerve terminal of neurons, and increases the intracellular concentration of calcium ions. Synapsin I was shown to be phosphorylated by this calcium influx.[16] The calcium ion, Ca2+, binds to calmodulin to form a calcium/calmodulin complex which then activates the calcium/calmodulin-dependent protein kinase, in turn triggering phosphorylation.[14] Calcium/calmodulin-dependent phosphorylation of synapsin I causes dissociation of synapsin I from the vesicular membrane.
In the nerve terminal ending, there are two pools of synaptic vesicles, the reserve pool and the ready-release pool. The reserve pool refers to the synaptic vesicles that are not ready to release neurotransmitters and the ready-release pool refers to the vesicles which are primed to release their neurotransmitters across the presynaptic cytoplasmic membrane and into the synaptic cleft. The removal of Synapsin I from synaptic vesicles is thought to mobilize synaptic vesicles from the reserve pool to the release-ready pool, thereby modulating neurotransmitter release. Since it is only present in the vesicles in the reserve pool, the non-phosphorylated form of Synapsin I is considered to be an inhibitory regulator of neurotransmission.
Clinical significance
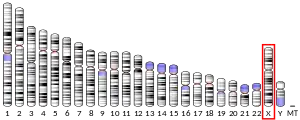
Mutations in the SYN1 gene may be associated with X-linked disorders with primary neuronal degeneration such as Rett syndrome.[6]
Discovery
The first member of the synapsin family, synapsin I was initially observed in 1973, as a neuronal membrane protein that was phosphorylated by membrane bound cAMP-dependent protein kinase. Synapsin I was detected by radioactive P-32 incorporated into the unknown protein through phosphorylation, using a then newly developed technique: a combination of SDS gel electrophoresis and autoradiography.[11] This groundbreaking technique allowed the advancement of the analysis of phosphorylated proteins, and introduced the identification of specific proteins. This was accomplished by the measuring the radioactivity through autoradiography of the individual protein bands phosphorylated by radioactive ATP, which is radio-labelled with P-32 at the gamma phosphate.
In 1977, at the same laboratory at Yale University, this first neuronal phosphoprotein was purified and initially characterized by Tetsufumi Ueda and Nobel Prize winner Paul Greengard. Originally named Protein I, it was found as an endogenous substrate for cAMP-dependent protein kinase in the synaptic membrane of the rat brain and was the first collagenous protein to be described in the nervous system.[13]
References
- GRCh38: Ensembl release 89: ENSG00000008056 - Ensembl, May 2017
- GRCm38: Ensembl release 89: ENSMUSG00000037217 - Ensembl, May 2017
- "Human PubMed Reference:". National Center for Biotechnology Information, U.S. National Library of Medicine.
- "Mouse PubMed Reference:". National Center for Biotechnology Information, U.S. National Library of Medicine.
- Südhof TC (May 1990). "The structure of the human synapsin I gene and protein". J. Biol. Chem. 265 (14): 7849–52. doi:10.1016/S0021-9258(19)39008-8. PMID 2110562.
- "Entrez Gene: SYN1 synapsin I".
- Huttner WB, Schiebler W, Greengard P, De Camilli P (May 1983). "Synapsin I (protein I), a nerve terminal-specific phosphoprotein. III. Its association with synaptic vesicles studied in a highly purified synaptic vesicle preparation". J. Cell Biol. 96 (5): 1374–88. doi:10.1083/jcb.96.5.1374. PMC 2112660. PMID 6404912.
- Ueda, T. & Greengard, P. (1977–1978). "Unpublished findings". Original Manuscript.
- Südhof TC, Czernik AJ, Kao HT, Takei K, Johnston PA, Horiuchi A, Kanazir SD, Wagner MA, Perin MS, De Camilli P (September 1989). "Synapsins: mosaics of shared and individual domains in a family of synaptic vesicle phosphoproteins". Science. 245 (4925): 1474–80. Bibcode:1989Sci...245.1474S. doi:10.1126/science.2506642. PMID 2506642.
- Hosaka M, Südhof TC (June 1999). "Homo- and heterodimerization of synapsins". J. Biol. Chem. 274 (24): 16747–53. doi:10.1074/jbc.274.24.16747. PMID 10358015.
- De Camilli P, Cameron R, Greengard P (May 1983). "Synapsin I (protein I), a nerve terminal-specific phosphoprotein. I. Its general distribution in synapses of the central and peripheral nervous system demonstrated by immunofluorescence in frozen and plastic sections". J. Cell Biol. 96 (5): 1337–54. doi:10.1083/jcb.96.5.1337. PMC 2112636. PMID 6404910.
- Ueda T, Maeno H, Greengard P (December 1973). "Regulation of endogenous phosphorylation of specific proteins in synaptic membrane fractions from rat brain by adenosine 3':5'-monophosphate". J. Biol. Chem. 248 (23): 8295–305. doi:10.1016/S0021-9258(19)43227-4. PMID 4356625.
- Ueda T, Greengard P (July 1977). "Adenosine 3':5'-monophosphate-regulated phosphoprotein system of neuronal membranes. I. Solubilization, purification, and some properties of an endogenous phosphoprotein". J. Biol. Chem. 252 (14): 5155–63. doi:10.1016/S0021-9258(17)40170-0. PMID 194903.
- Schulman H, Greengard P (February 1978). "Stimulation of brain membrane protein phosphorylation by calcium and an endogenous heat-stable protein". Nature. 271 (5644): 478–9. Bibcode:1978Natur.271..478S. doi:10.1038/271478a0. PMID 628428. S2CID 4288598.
- Kennedy MB, McGuinness T, Greengard P (April 1983). "A calcium/calmodulin-dependent protein kinase from mammalian brain that phosphorylates Synapsin I: partial purification and characterization". J. Neurosci. 3 (4): 818–31. doi:10.1523/JNEUROSCI.03-04-00818.1983. PMC 6564459. PMID 6403674.
- Krueger BK, Forn J, Greengard P (April 1977). "Depolarization-induced phosphorylation of specific proteins, mediated by calcium ion influx, in rat brain synaptosomes". J. Biol. Chem. 252 (8): 2764–73. doi:10.1016/S0021-9258(17)40523-0. PMID 323254.
- Jaffrey, Samie R; Benfenati Fabio; Snowman Adele M; Czernik Andrew J; Snyder Solomon H (Mar 2002). "Neuronal nitric-oxide synthase localization mediated by a ternary complex with synapsin and CAPON". Proc. Natl. Acad. Sci. U.S.A. United States. 99 (5): 3199–204. Bibcode:2002PNAS...99.3199J. doi:10.1073/pnas.261705799. ISSN 0027-8424. PMC 122496. PMID 11867766.
Further reading
- Krueger BK, Forn J, Greengard P (April 1977). "Depolarization-induced phosphorylation of specific proteins, mediated by calcium ion influx, in rat brain synaptosomes". J. Biol. Chem. 252 (8): 2764–73. doi:10.1016/S0021-9258(17)40523-0. PMID 323254.
- Greengard, Paul (1978). Cyclic nucleotides, phosphorylated proteins, and neuronal function. New York: Raven Press. ISBN 0-89004-281-0.
- Greengard, P. (1981). Intracellular Signals in the Brain. Harvey Lecture Series. Vol. 75. Academic Press. pp. 277–331.
PDB gallery | |
|---|---|
|