Three-toed sloth
The three-toed or three-fingered sloths are arboreal neotropical mammals.[2] They are the only members of the genus Bradypus (meaning "slow-footed") and the family Bradypodidae. The five living species of three-toed sloths are the brown-throated sloth, the maned sloth, the pale-throated sloth, the southern maned sloth, and the pygmy three-toed sloth. In complete contrast to past morphological studies, which tended to place Bradypus as the sister group to all other folivorans, molecular studies place them nested within the sloth superfamily Megatherioidea, making them the only surviving members of that radiation.[2][3]
| Three-toed sloths[1] | |
|---|---|
 | |
| Brown-throated three-toed sloth (Bradypus variegatus) | |
| Scientific classification | |
| Domain: | Eukaryota |
| Kingdom: | Animalia |
| Phylum: | Chordata |
| Class: | Mammalia |
| Order: | Pilosa |
| Superfamily: | Megatherioidea |
| Family: | Bradypodidae Gray, 1821 |
| Genus: | Bradypus Linnaeus, 1758 |
| Type species | |
| Bradypus tridactylus | |
| Species | |
|
4, see text | |
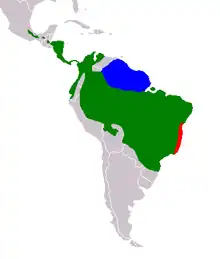 | |
| Green: B. variegatus, blue: B. tridactylus, red: B. torquatus | |
Extant species
| Image | Scientific name | Common name | Distribution |
|---|---|---|---|
 | B. pygmaeus | Pygmy three-toed sloth | Isla Escudos de Veraguas (Panama) |
.JPG.webp) | B. torquatus | Maned three-toed sloth | Atlantic coastal rainforest of southeastern Brazil |
 | B. tridactylus | Pale-throated three-toed sloth | Part of northern South America, including Guyana, Suriname, French Guiana, eastern Venezuela and Brazil north of the Amazon River |
 | B. variegatus | Brown-throated three-toed sloth | Central America and much of north and central South America, from Honduras through Colombia, Venezuela, Ecuador, eastern Peru, Bolivia and Brazil |
 | B. crinitus | Southern maned sloth | Coastal Brazil in Rio de Janeiro and Espirito Santo.[4] |
Evolution
A study of mitochondrial cytochrome b and 16S rRNA sequences suggests that B. torquatus diverged from B. variegatus and B. tridactylus about 12 million years ago, while the latter two split 5 to 6 million years ago. The diversification of B. variegatus lineages was estimated to have started 4 to 5 million years ago.[5]
Relation to the two-toed sloth
Both types of sloth tend to occupy the same forests; in most areas, a particular species of three-toed sloth and a single species of the somewhat larger and generally faster-moving two-toed sloth will jointly predominate. Although similar in overall appearance, the two genera are placed in different families. Recent phylogenetic analyses support the morphological data from the 1970s and 1980s that the two genera are not closely related and that each adopted their arboreal lifestyles independently.[6] From morphological studies it was unclear from which ground-dwelling sloth taxa the three-toed sloths evolved or whether they retained their arboreality from the last common ancestor of extant sloths. The two-toed sloths were thought on the basis of morphology to nest phylogenetically within one of the divisions of ground-dwelling Caribbean sloths.[7]
Understanding of sloth phylogeny has recently been greatly revised by molecular studies, based on collagen[3] and mitochondrial DNA[2] sequences. These investigations consistently place three-toed sloths within Megatherioidea, close to Megalonyx, megatheriids and nothrotheriids, and two-toed sloths close to mylodontids, while moving the Caribbean sloths to a separate, basal branch of the sloth evolutionary tree.[3][2] These results provide further strong support for the long-held belief that arboreality arose separately in the two genera via convergent paths.
The following sloth family phylogenetic tree is based on collagen and mitochondrial DNA sequence data (see Fig. 4 of Presslee et al., 2019).[3]
| Folivora |
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Characteristics
Famously slow-moving, a sloth travels at an average speed of 0.24 km/h (0.15 mph).[8] Three-toed sloths are about the size of a small dog or a large cat, with the head and body having a combined length of around 45 cm (18 in) and a weight of 3.5–4.5 kg (8–10 lb). Unlike the two-toed sloths, they also have short tails of 6–7 cm (2–3 in), and they have three clawed toes on each limb. All sloths have three digits on their hindlimbs; the difference is found in the number of digits on the forelimbs; thus they are sometimes referred to as three-fingered sloths.[9] However, sloths are generally regarded as quadrupeds.
Behavior
Like the two-toed sloth, three-toed sloths are agile swimmers. They are still slow in trees.[10] The muscles that sloths use to grip and produce a pulling motion are much more prominent than those that produce a pushing motion.[11] This means that they struggle to support their body weight when walking on all four limbs, so traveling on the ground is a dangerous and laborious process.[12]
Three-toed sloths are arboreal (tree-dwelling), with a body adapted to hang by their limbs. Large, curved claws and muscles specifically adapted for strength and stamina help sloths to keep a strong grip on tree branches.[13] The abdominal organs close to their diaphragm (such as their stomach, liver, and kidneys) are attached to their lower ribs (or pelvic girdle in the latter case) by fibrinous adhesions, which prevent the weight of these organs from compressing their lungs when hanging, making inhalation easier.[14]
They live high in the canopy but descend once a week to defecate on the forest floor. During this week-long interval, their feces and urine accumulate to about a third of their total body mass.[14] It takes about a month for a single leaf to pass through its four-chambered stomach and digestive tract.[15] Although they get most of their fluids from the leaves that they eat, they have been observed drinking directly from rivers.[16] Because of their slow metabolism, they do not need to ingest many leaves on a daily basis, but when ambient temperatures are high, the symbiotic microbes and bacteria present in their gut will break down and ferment food at a faster rate.[17] Conversely, when temperatures are lower, sloths will consume less, which is opposite to what has been observed in most other mammals.[15] Only very few species of sloths are found at higher altitudes, and these are found to have thicker coats than those living in lower altitudes. Some of the extinct species of sloths were able to tolerate cooler temperatures, but researchers believe this was probably due to thicker fur, larger size, larger muscle mass, and more access to a constant food supply.[18]
Their long, coarse fur often appears greenish, not due to pigment, but to algae growing on it. Sloths' greenish color and their sluggish habits provide an effective camouflage; hanging quietly, sloths resemble a bundle of leaves.
They move between different trees up to four times a day, although they prefer to keep to a particular type of tree, which varies between individuals, perhaps as a means of allowing multiple sloths to occupy overlapping home ranges without competing with each other.[19]
Three-toed sloths are predominantly diurnal, although they can be active at any hour of the day, while two-toed sloths are nocturnal.[20]
Lifecycle

Members of this genus tend to live around 25 to 30 years, reaching sexual maturation at three to five years of age.
Three-toed sloths do not have a mating season and breed year-round. Females give birth to a single young after a gestation period of around six months. The offspring cling to their mother's bellies for around nine months. They are weaned around nine months of age when the mother leaves her home territory to her offspring and moves elsewhere. Adults are solitary, and mark their territories using anal scent glands and dung middens.[19]
Male three-toed sloths are attracted to females in estrus by their screams echoing throughout the canopy. Sloth copulation lasts an average of 25 minutes.[21] Male three-toed sloths are strongly polygamous and exclude competitors from their territory. Males are also able to compete with one another within small habitable territories.[22]
The home ranges used by wild brown-throated three-toed sloths in Costa Rica include cacao, pasture, riparian forests, peri-urban areas, and living fence-rows.[23] For the first few months after giving birth, mothers remain at just one or two trees and guide their young. At about five to seven months of age, when the young have become more independent, mothers expand their resources and leave their young in new areas. During natal dispersion, three-toed sloths prefer tropical forests, often using riparian forest habitat to disperse while avoiding pastures and shade-grown cacao.[24] The home range for mothers is larger than those of young. After separation, only the mothers use the cacao-growing forest, but both use riparian forests. Different types of trees are used by both mother and young, which indicates that this agricultural matrix provides an important habitat type for these animals.[25]
Dentition and skeleton
Three-toed sloths have no incisor or canine teeth, just a set of peg-shaped cheek teeth that are not clearly divided into premolars and molars, and lack homology with those teeth in other mammals, and thus are referred to as molariforms. The molariform dentition in three-toed sloths is simple and can be characterized as dental formula of: 54-5.[26]
Three-toed sloths are unusual amongst mammals in possessing as many as nine cervical vertebrae, which may be due to mutations in the homeotic genes.[27] All other mammals have seven cervical vertebrae,[28] other than the two-toed sloth and the manatee, which have only six.
References
- Gardner, A. L. (2005). "Order Pilosa". In Wilson, D. E.; Reeder, D. M. (eds.). Mammal Species of the World: A Taxonomic and Geographic Reference (3rd ed.). Johns Hopkins University Press. pp. 100–101. ISBN 978-0-8018-8221-0. OCLC 62265494.
- Delsuc, F.; Kuch, M.; Gibb, G.C.; Karpinski, E.; Hackenberger, D.; Szpak, P.; et al. (2019). "Ancient Mitogenomes reveal the evolutionary history and biogeography of sloths". Current Biology. 29 (12): 2031–2042.e6. doi:10.1016/j.cub.2019.05.043. hdl:11336/136908. PMID 31178321.
- Presslee, S.; Slater, G.J.; Pujos, F.; Forasiepi, A.M.; Fischer, R.; Molloy, K.; et al. (2019). "Palaeoproteomics resolves sloth relationships". Nature Ecology & Evolution. 3 (7): 1121–1130. doi:10.1038/s41559-019-0909-z. PMID 31171860. S2CID 174813630.
- Miranda, Flavia (February 2023). "Taxonomic revision of maned sloths, subgenus Bradypus (Scaeopus), Pilosa, Bradypodidae, with revalidation of Bradypus crinitus Gray, 1850". Journal of Mammalogy. 104 (1): 86–103. doi:10.1093/jmammal/gyac059.
- de Moraes-Barros, N.; Silva, J. A. B.; Morgante, J. S. (2011-02-16). "Morphology, molecular phylogeny, and taxonomic inconsistencies in the study of Bradypus sloths (Pilosa: Bradypodidae)". Journal of Mammalogy. 92 (1): 86–100. doi:10.1644/10-MAMM-A-086.1.
- Hoss, Matthias; Dilling, Amrei; Currant, Andrew; Paabo, Svante (9 Jan 1996). "Molecular phylogeny of the extinct ground sloth Mylodon darwinii". Proceedings of the National Academy of Sciences. 93 (1): 181–185. Bibcode:1996PNAS...93..181H. doi:10.1073/pnas.93.1.181. PMC 40202. PMID 8552600.
- White, J.L.; MacPhee, R.D.E. (2001). "The sloths of the West Indies: a systematic and phylogenetic review". In Woods, C.A.; Sergile, F.E. (eds.). Biogeography of the West Indies: Patterns and perspectives. Boca Raton, FL / London, UK / New York, NY / Washington, DC: CRC Press. pp. 201–235. doi:10.1201/9781420039481-14. ISBN 978-0-8493-2001-9.
- "Speed of Animals". infoplease.com.
- Judy Avey-Arroyo for Gage, L. J. (2008). Hand-Rearing Wild and Domestic Mammals, Iowa State University Press, 2002, pages 81-89.doi:10.1002/9780470385005.ch12
- Three-toed Sloth, National Geographic.
- Olson, Rachel A.; Glenn, Zachary D.; Cliffe, Rebecca N.; Butcher, Michael T. (2018-12-01). "Architectural Properties of Sloth Forelimb Muscles (Pilosa: Bradypodidae)". Journal of Mammalian Evolution. 25 (4): 573–588. doi:10.1007/s10914-017-9411-z. ISSN 1573-7055. S2CID 22238582.
- Keating, Ian D. (2018-01-01), Audacious Sloth, retrieved 2020-03-30
- Olson, R.A.; Glenn, Z.D.; Cliffe, R.N.; Butcher, M.T. (2018-12-01). "Architectural Properties of Sloth Forelimb Muscles (Pilosa: Bradypodidae)". Journal of Mammalian Evolution. 25 (4): 573–588. doi:10.1007/s10914-017-9411-z. S2CID 22238582.
- Cliffe, R.N.; Avey-Arroyo, J.A.; Arroyo, F.J.; Holton, M.D.; Wilson, R.P. (2014-04-30). "Mitigating the squash effect: sloths breathe easily upside down". Biology Letters. 10 (4): 20140172. doi:10.1098/rsbl.2014.0172. PMC 4013704. PMID 24759371.
- Cliffe, Rebecca N.; Haupt, Ryan J.; Avey-Arroyo, Judy A.; Wilson, Rory P. (2015-04-02). "Sloths like it hot: ambient temperature modulates food intake in the brown-throated sloth (Bradypus variegatus)". PeerJ. 3: e875. doi:10.7717/peerj.875. ISSN 2167-8359. PMC 4389270. PMID 25861559.
- Cliffe, Rebecca N.; Haupt, Ryan J. (2018). "Hanging out for a drink". Frontiers in Ecology and the Environment. 16 (8): 446. doi:10.1002/fee.1955. ISSN 1540-9309.
- Cliffe, R.N.; Haupt, R.J.; Avey-Arroyo, J.A.; Wilson, R.P. (2015-04-02). "Sloths like it hot: ambient temperature modulates food intake in the brown-throated sloth (Bradypus variegatus)". PeerJ. 3: e875. doi:10.7717/peerj.875. PMC 4389270. PMID 25861559.
- Gilmore, D. P.; Da Costa, C. P.; Duarte, D. P. F. (January 2001). "Sloth biology: an update on their physiological ecology, behavior and role as vectors of arthropods and arboviruses". Brazilian Journal of Medical and Biological Research. 34 (1): 9–25. doi:10.1590/S0100-879X2001000100002. ISSN 0100-879X. PMID 11151024.
- Dickman, Christopher R. (1984). Macdonald, D. (ed.). The Encyclopedia of Mammals. New York: Facts on File. pp. 776–779. ISBN 0-87196-871-1.
- Eisenberg, John F.; Redford, Kent H. (May 15, 2000). Mammals of the Neotropics, Volume 3: The Central Neotropics: Ecuador, Peru, Bolivia, Brazil. University of Chicago Press. pp. 624 (see pp. 94–95, 97). ISBN 978-0-226-19542-1. OCLC 493329394.
- Gilmore, D.P.; et al. (2000). "An update on the physiology of two- and three-toed sloths". Brazilian Journal of Medical and Biological Research. 33 (2): 129–146. doi:10.1590/S0100-879X2000000200001. PMID 10657054.
- Pauli, J.N. & Peery, M.Z. (2012). "Unexpected strong polygyny in the brown-throated three-toed sloth". PLOS ONE. 7 (12): e51389. Bibcode:2012PLoSO...751389P. doi:10.1371/journal.pone.0051389. PMC 3526605. PMID 23284687.
- Garcés‐Restrepo, M.F.; Pauli, J.N.; Peery, M.Z. (2018). "Natal dispersal of tree sloths in a human-dominated landscape: Implications for tropical biodiversity conservation". Journal of Applied Ecology. 55 (5): 2253–2262. doi:10.1111/1365-2664.13138.
- Garcés‐Restrepo, Mario F.; Pauli, Jonathan N.; Peery, M. Zachariah (2018). "Natal dispersal of tree sloths in a human-dominated landscape: Implications for tropical biodiversity conservation". Journal of Applied Ecology. 55 (5): 2253–2262. doi:10.1111/1365-2664.13138. ISSN 1365-2664.
- Ramirez, O.; et al. (2011). "Temporal and spatial resource use by female three-toed sloths and their young in an agricultural landscape in Costa Rica". Revista de Biología Tropical. 59 (4): 1743–1755. PMID 22208090.
- Ungar, P.S. (2010). Mammal Teeth: Origin, Evolution, and Diversity. Baltimore, US: Johns Hopkins University Press. pp. 145–6. ISBN 9780801899515.
- "Sticking Their Necks out for Evolution: Why Sloths and Manatees Have Unusually Long (or Short) Necks". May 6th 2011. Science Daily. Retrieved 25 July 2013.
- Frietson Galis (1999). "Why do almost all mammals have seven cervical vertebrae? Developmental constraints, Hox genes and Cancer" (PDF). Journal of Experimental Zoology. 285 (1): 19–26. doi:10.1002/(SICI)1097-010X(19990415)285:1<19::AID-JEZ3>3.0.CO;2-Z. PMID 10327647. Archived from the original (PDF) on 2004-11-10.

