Ubiquitin-like protein
Ubiquitin-like proteins (UBLs) are a family of small proteins involved in post-translational modification of other proteins in a cell, usually with a regulatory function. The UBL protein family derives its name from the first member of the class to be discovered, ubiquitin (Ub), best known for its role in regulating protein degradation through covalent modification of other proteins. Following the discovery of ubiquitin, many additional evolutionarily related members of the group were described, involving parallel regulatory processes and similar chemistry. UBLs are involved in a widely varying array of cellular functions including autophagy, protein trafficking, inflammation and immune responses, transcription, DNA repair, RNA splicing, and cellular differentiation.[1][2][3]
| Ubiquitin family | |||||||||
|---|---|---|---|---|---|---|---|---|---|
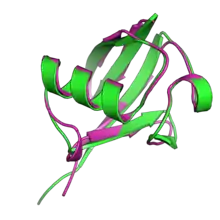 | |||||||||
| Identifiers | |||||||||
| Symbol | Ubiquitin | ||||||||
| Pfam | PF00240 | ||||||||
| InterPro | IPR029071 | ||||||||
| SMART | SM00213 | ||||||||
| |||||||||
Discovery
Ubiquitin itself was first discovered in the 1970s and originally named "ubiquitous immunopoietic polypeptide".[4] Subsequently, other proteins with sequence similarity to ubiquitin were occasionally reported in the literature, but the first shown to share the key feature of covalent protein modification was ISG15, discovered in 1987.[5] A succession of reports in the mid 1990s is recognized as a turning point in the field,[6] with the discovery of SUMO (small ubiquitin-like modifier, also known as Sentrin or SENP1) reported around the same time by a variety of investigators in 1996,[7] NEDD8 in 1997,[8] and Apg12 in 1998.[9] A systematic survey has since identified over 10,000 distinct genes for ubiquitin or ubiquitin-like proteins represented in eukaryotic genomes.[10]
Structure and classification
Members of the UBL family are small, non-enzymatic proteins that share a common structure exemplified by ubiquitin, which has 76 amino acid residues arranged into a "beta-grasp" protein fold consisting of a five-strand antiparallel beta sheet surrounding an alpha helix.[1][11][12] The beta-grasp fold is widely distributed in other proteins of both eukaryotic and prokaryotic origin.[13] Collectively, ubiquitin and ubiquitin-like proteins are sometimes referred to as "ubiquitons".[3]
UBLs can be divided into two categories depending on their ability to be covalently conjugated to other molecules. UBLs that are capable of conjugation (sometimes known as Type I) have a characteristic sequence motif consisting of one to two glycine residues at the C-terminus, through which covalent conjugation occurs. Typically, UBLs are expressed as inactive precursors and must be activated by proteolysis of the C-terminus to expose the active glycine.[1][12] Almost all such UBLs are ultimately linked to another protein, but there is at least one exception; ATG8 is linked to phosphatidylethanolamine.[1] UBLs that do not exhibit covalent conjugation (Type II) often occur as protein domains genetically fused to other domains in a single larger polypeptide chain, and may be proteolytically processed to release the UBL domain[1] or may function as protein-protein interaction domains.[11] UBL domains of larger proteins are sometimes known as UBX domains.[14]
Distribution
Ubiquitin is, as its name suggests, ubiquitous in eukaryotes; it is traditionally considered to be absent in bacteria and archaea,[11] though a few examples have been described in archaea.[15] UBLs are also widely distributed in eukaryotes, but their distribution varies among lineages; for example, ISG15, involved in the regulation of the immune system, is not present in lower eukaryotes.[1] Other families exhibit diversification in some lineages; a single member of the SUMO family is found in the yeast genome, but there are at least four in vertebrate genomes, which show some functional redundancy,[1][2] and there are at least eight in the genome of the model plant Arabidopsis thaliana.[16]
In humans
The human genome encodes at least eight families of UBLs, not including ubiquitin itself, that are considered Type I UBLs and are known to covalently modify other proteins: SUMO, NEDD8, ATG8, ATG12, URM1, UFM1, FAT10, and ISG15.[1] One additional protein, known as FUBI, is encoded as a fusion protein in the FAU gene, and is proteolytically processed to generate a free glycine C-terminus, but has not been experimentally demonstrated to form covalent protein modifications.[1]
In plants
Plant genomes are known to encode at least seven families of UBLs in addition to ubiquitin: SUMO, RUB (the plant homolog of NEDD8), ATG8, ATG12, MUB, UFM1, and HUB1, as well as a number of Type II UBLs.[17] Some UBL families and their associated regulatory proteins in plants have undergone dramatic expansion, likely due to both whole genome duplication and other forms of gene duplication; the ubiquitin, SUMO, ATG8, and MUB families have been estimated to account for almost 90% of plants' UBL genes.[18] Proteins associated with ubiquitin and SUMO signaling are highly enriched in the genomes of embryophytes.[15]
In prokaryotes
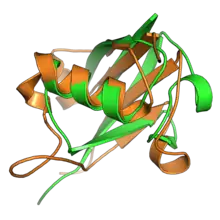
In comparison to eukaryotes, prokaryotic proteins with relationships to UBLs are phylogenetically restricted.[19][20] Prokaryotic ubiquitin-like protein (Pup) occurs in some actinobacteria and has functions closely analogous to ubiquitin in labeling proteins for proteasomal degradation; however it is intrinsically disordered and its evolutionary relationship to UBLs is unclear.[19] A related protein UBact in some Gram-negative lineages has recently been described.[21] By contrast, the protein TtuB in bacteria of the genus Thermus does share the beta-grasp fold with eukaryotic UBLs; it is reported to have dual functions as both a sulfur carrier protein and a covalently conjugated protein modification.[19] In archaea, the small archaeal modifier proteins (SAMPs) share the beta-grasp fold and have been shown to play a ubiquitin-like role in protein degradation.[19][20] Recently, a seemingly complete set of genes corresponding to a eukaryote-like ubiquitin pathway was identified in an uncultured archaeon in 2011,[22][23][24] and at least three lineages of archaea—"Euryarchaeota", Thermoproteota (formerly Crenarchaeota), and "Aigarchaeota"—are believed to possess such systems.[15][25][26] In addition, some pathogenic bacteria have evolved proteins that mimic those in eukaryotic UBL pathways and interact with UBLs in the host cell, interfering with their signaling function.[27][28]
Regulation
Regulation of UBLs that are capable of covalent conjugation in eukaryotes is elaborate but typically parallel for each member of the family, best characterized for ubiquitin itself. The process of ubiquitination is a tightly regulated three-step sequence: activation, performed by ubiquitin-activating enzymes (E1); conjugation, performed by ubiquitin-conjugating enzymes (E2); and ligation, performed by ubiquitin ligases (E3). The result of this process is the formation of a covalent bond between the C-terminus of ubiquitin and a residue (typically a lysine) on the target protein. Many UBL families have a similar three-step process catalyzed by a distinct set of enzymes specific to that family.[1][29][30] Deubiquitination or deconjugation - that is, removal of ubiquitin from a protein substrate - is performed by deubiquitinating enzymes (DUBs); UBLs can also be degraded through the action of ubiquitin-specific proteases (ULPs).[31] The range of UBLs on which these enzymes can act is variable and can be difficult to predict. Some UBLs, such as SUMO and NEDD8, have family-specific DUBs and ULPs.[32]
Ubiquitin is capable of forming polymeric chains, with additional ubiquitin molecules covalently attached to the first, which in turn is attached to its protein substrate. These chains may be linear or branched, and different regulatory signals may be sent by differences in the length and branching of the ubiquitin chain.[31] Although not all UBL families are known to form chains, SUMO, NEDD8, and URM1 chains have all been experimentally detected.[1] Additionally, ubiquitin can itself be modified by UBLs, known to occur with SUMO and NEDD8.[31][33] The best-characterized intersections between distinct UBL families involve ubiquitin and SUMO.[34][35]
Cellular functions
UBLs as a class are involved in a very large variety of cellular processes. Furthermore, individual UBL families vary in the scope of their activities and the diversity of the proteins to which they are conjugated.[1] The best known function of ubiquitin is identifying proteins to be degraded by the proteasome, but ubiquitination can play a role in other processes such as endocytosis and other forms of protein trafficking, transcription and transcription factor regulation, cell signaling, histone modification, and DNA repair.[11][12][36] Most other UBLs have similar roles in regulating cellular processes, usually with a more restricted known range than that of ubiquitin itself. SUMO proteins have the widest variety of cellular protein targets after ubiquitin[1] and are involved in processes including transcription, DNA repair, and the cellular stress response.[33] NEDD8 is best known for its role in regulating cullin proteins, which in turn regulate ubiquitin-mediated protein degradation,[2] though it likely also has other functions.[37] Two UBLs, ATG8 and ATG12, are involved in the process of autophagy;[38] both are unusual in that ATG12 has only two known protein substrates and ATG8 is conjugated not to a protein but to a phospholipid, phosphatidylethanolamine.[1]
Evolution
The evolution of UBLs and their associated suites of regulatory proteins has been of interest since shortly after they were recognized as a family.[39] Phylogenetic studies of the beta-grasp protein fold superfamily suggest that eukaryotic UBLs are monophyletic, indicating a shared evolutionary origin.[13] UBL regulatory systems - including UBLs themselves and the cascade of enzymes that interact with them - are believed to share a common evolutionary origin with prokaryotic biosynthesis pathways for the cofactors thiamine and molybdopterin; the bacterial sulfur transfer proteins ThiS and MoaD from these pathways share the beta-grasp fold with UBLs, while sequence similarity and a common catalytic mechanism link pathway members ThiF and MoeB to ubiquitin-activating enzymes.[13][17][11] Interestingly, the eukaryotic protein URM1 functions as both a UBL and a sulfur-carrier protein, and has been described as a molecular fossil establishing this evolutionary link.[11][40]
Comparative genomics surveys of UBL families and related proteins suggest that UBL signaling was already well-developed in the last eukaryotic common ancestor and ultimately originates from ancestral archaea,[15] a theory supported by the observation that some archaeal genomes possess the necessary genes for a fully functioning ubiquitination pathway.[25][18] Two different diversification events within the UBL family have been identified in eukaryotic lineages, corresponding to the origin of multicellularity in both animal and plant lineages.[15]
References
- Cappadocia L, Lima CD (February 2018). "Ubiquitin-like Protein Conjugation: Structures, Chemistry, and Mechanism". Chemical Reviews. 118 (3): 889–918. doi:10.1021/acs.chemrev.6b00737. PMC 5815371. PMID 28234446.
- van der Veen AG, Ploegh HL (7 July 2012). "Ubiquitin-like proteins". Annual Review of Biochemistry. 81 (1): 323–57. doi:10.1146/annurev-biochem-093010-153308. PMID 22404627.
- Welchman RL, Gordon C, Mayer RJ (August 2005). "Ubiquitin and ubiquitin-like proteins as multifunctional signals". Nature Reviews. Molecular Cell Biology. 6 (8): 599–609. doi:10.1038/nrm1700. PMID 16064136. S2CID 7373421.
- Goldstein G, Scheid M, Hammerling U, Schlesinger DH, Niall HD, Boyse EA (January 1975). "Isolation of a polypeptide that has lymphocyte-differentiating properties and is probably represented universally in living cells". Proceedings of the National Academy of Sciences of the United States of America. 72 (1): 11–5. Bibcode:1975PNAS...72...11G. doi:10.1073/pnas.72.1.11. PMC 432229. PMID 1078892.
- Haas AL, Ahrens P, Bright PM, Ankel H (August 1987). "Interferon induces a 15-kilodalton protein exhibiting marked homology to ubiquitin". The Journal of Biological Chemistry. 262 (23): 11315–23. doi:10.1016/S0021-9258(18)60961-5. PMID 2440890.
- Yeh ET, Gong L, Kamitani T (May 2000). "Ubiquitin-like proteins: new wines in new bottles". Gene. 248 (1–2): 1–14. doi:10.1016/S0378-1119(00)00139-6. PMID 10806345.
- Saitoh, Hisato; Pu, Robert T.; Dasso, Mary (October 1997). "SUMO-1: wrestling with a new ubiquitin-related modifier". Trends in Biochemical Sciences. 22 (10): 374–376. doi:10.1016/S0968-0004(97)01102-X. PMID 9357311.
- Kamitani T, Kito K, Nguyen HP, Yeh ET (November 1997). "Characterization of NEDD8, a developmentally down-regulated ubiquitin-like protein". The Journal of Biological Chemistry. 272 (45): 28557–62. doi:10.1074/jbc.272.45.28557. PMID 9353319.
- Mizushima N, Noda T, Yoshimori T, Tanaka Y, Ishii T, George MD, Klionsky DJ, Ohsumi M, Ohsumi Y (September 1998). "A protein conjugation system essential for autophagy". Nature. 395 (6700): 395–8. Bibcode:1998Natur.395..395M. doi:10.1038/26506. PMID 9759731. S2CID 204997310.
- Zhou J, Xu Y, Lin S, Guo Y, Deng W, Zhang Y, Guo A, Xue Y (January 2018). "iUUCD 2.0: an update with rich annotations for ubiquitin and ubiquitin-like conjugations". Nucleic Acids Research. 46 (D1): D447–D453. doi:10.1093/nar/gkx1041. PMC 5753239. PMID 29106644.
- Hochstrasser M (March 2009). "Origin and function of ubiquitin-like proteins". Nature. 458 (7237): 422–9. Bibcode:2009Natur.458..422H. doi:10.1038/nature07958. PMC 2819001. PMID 19325621.
- Kerscher O, Felberbaum R, Hochstrasser M (November 2006). "Modification of proteins by ubiquitin and ubiquitin-like proteins". Annual Review of Cell and Developmental Biology. 22 (1): 159–80. doi:10.1146/annurev.cellbio.22.010605.093503. PMID 16753028.
- Burroughs AM, Balaji S, Iyer LM, Aravind L (July 2007). "Small but versatile: the extraordinary functional and structural diversity of the beta-grasp fold". Biology Direct. 2 (1): 18. doi:10.1186/1745-6150-2-18. PMC 1949818. PMID 17605815.
- Buchberger A, Howard MJ, Proctor M, Bycroft M (March 2001). "The UBX domain: a widespread ubiquitin-like module". Journal of Molecular Biology. 307 (1): 17–24. doi:10.1006/jmbi.2000.4462. PMID 11243799.
- Grau-Bové X, Sebé-Pedrós A, Ruiz-Trillo I (March 2015). "The eukaryotic ancestor had a complex ubiquitin signaling system of archaeal origin". Molecular Biology and Evolution. 32 (3): 726–39. doi:10.1093/molbev/msu334. PMC 4327156. PMID 25525215.
- Miura K, Hasegawa PM (April 2010). "Sumoylation and other ubiquitin-like post-translational modifications in plants". Trends in Cell Biology. 20 (4): 223–32. doi:10.1016/j.tcb.2010.01.007. PMID 20189809.
- Vierstra RD (September 2012). "The expanding universe of ubiquitin and ubiquitin-like modifiers". Plant Physiology. 160 (1): 2–14. doi:10.1104/pp.112.200667. PMC 3440198. PMID 22693286.
- Hua Z, Doroodian P, Vu W (July 2018). "Contrasting duplication patterns reflect functional diversities of ubiquitin and ubiquitin-like protein modifiers in plants". The Plant Journal. 95 (2): 296–311. doi:10.1111/tpj.13951. PMID 29738099.
- Maupin-Furlow JA (2014). "Prokaryotic ubiquitin-like protein modification". Annual Review of Microbiology. 68: 155–75. doi:10.1146/annurev-micro-091313-103447. PMC 4757901. PMID 24995873.
- Ganguli, S; Ratna Prabha, C (2017). "Pups, SAMPs, and Prokaryotic Proteasomes". In Chakraborti, S; Dhalla, N (eds.). Proteases in physiology and pathology. Springer. ISBN 978-981-10-2512-9.
- Lehmann G, Udasin RG, Livneh I, Ciechanover A (February 2017). "Identification of UBact, a ubiquitin-like protein, along with other homologous components of a conjugation system and the proteasome in different gram-negative bacteria". Biochemical and Biophysical Research Communications. 483 (3): 946–950. doi:10.1016/j.bbrc.2017.01.037. PMID 28087277.
- Nunoura T, Takaki Y, Kakuta J, Nishi S, Sugahara J, Kazama H, Chee GJ, Hattori M, Kanai A, Atomi H, Takai K, Takami H (April 2011). "Insights into the evolution of Archaea and eukaryotic protein modifier systems revealed by the genome of a novel archaeal group". Nucleic Acids Research. 39 (8): 3204–23. doi:10.1093/nar/gkq1228. PMC 3082918. PMID 21169198.
- Hennell James R, Caceres EF, Escasinas A, Alhasan H, Howard JA, Deery MJ, Ettema TJ, Robinson NP (October 2017). "Functional reconstruction of a eukaryotic-like E1/E2/(RING) E3 ubiquitylation cascade from an uncultured archaeon". Nature Communications. 8 (1): 1120. Bibcode:2017NatCo...8.1120H. doi:10.1038/s41467-017-01162-7. PMC 5654768. PMID 29066714.
- Fuchs AC, Maldoner L, Wojtynek M, Hartmann MD, Martin J (July 2018). "Rpn11-mediated ubiquitin processing in an ancestral archaeal ubiquitination system". Nature Communications. 9 (1): 2696. Bibcode:2018NatCo...9.2696F. doi:10.1038/s41467-018-05198-1. PMC 6043591. PMID 30002364.
- Zaremba-Niedzwiedzka K, Caceres EF, Saw JH, Bäckström D, Juzokaite L, Vancaester E, Seitz KW, Anantharaman K, Starnawski P, Kjeldsen KU, Stott MB, Nunoura T, Banfield JF, Schramm A, Baker BJ, Spang A, Ettema TJ (January 2017). "Asgard archaea illuminate the origin of eukaryotic cellular complexity". Nature. 541 (7637): 353–358. Bibcode:2017Natur.541..353Z. doi:10.1038/nature21031. OSTI 1580084. PMID 28077874. S2CID 4458094.
- Hua ZS, Qu YN, Zhu Q, Zhou EM, Qi YL, Yin YR, Rao YZ, Tian Y, Li YX, Liu L, Castelle CJ, Hedlund BP, Shu WS, Knight R, Li WJ (July 2018). "Genomic inference of the metabolism and evolution of the archaeal phylum Aigarchaeota". Nature Communications. 9 (1): 2832. Bibcode:2018NatCo...9.2832H. doi:10.1038/s41467-018-05284-4. PMC 6053391. PMID 30026532.
- Zhou Y, Zhu Y (January 2015). "Diversity of bacterial manipulation of the host ubiquitin pathways". Cellular Microbiology. 17 (1): 26–34. doi:10.1111/cmi.12384. PMID 25339545. S2CID 33328949.
- Ribet D, Cossart P (November 2018). "Ubiquitin, SUMO, and NEDD8: Key Targets of Bacterial Pathogens" (PDF). Trends in Cell Biology. 28 (11): 926–940. doi:10.1016/j.tcb.2018.07.005. PMC 7028394. PMID 30107971.
- Streich FC, Lima CD (6 May 2014). "Structural and functional insights to ubiquitin-like protein conjugation". Annual Review of Biophysics. 43 (1): 357–79. doi:10.1146/annurev-biophys-051013-022958. PMC 4118471. PMID 24773014.
- Schulman BA, Harper JW (May 2009). "Ubiquitin-like protein activation by E1 enzymes: the apex for downstream signalling pathways". Nature Reviews. Molecular Cell Biology. 10 (5): 319–31. doi:10.1038/nrm2673. PMC 2712597. PMID 19352404.
- Mevissen TE, Komander D (June 2017). "Mechanisms of Deubiquitinase Specificity and Regulation". Annual Review of Biochemistry. 86 (1): 159–192. doi:10.1146/annurev-biochem-061516-044916. PMID 28498721.
- Ronau JA, Beckmann JF, Hochstrasser M (April 2016). "Substrate specificity of the ubiquitin and Ubl proteases". Cell Research. 26 (4): 441–56. doi:10.1038/cr.2016.38. PMC 4822132. PMID 27012468.
- Swatek KN, Komander D (April 2016). "Ubiquitin modifications". Cell Research. 26 (4): 399–422. doi:10.1038/cr.2016.39. PMC 4822133. PMID 27012465.
- Denuc A, Marfany G (February 2010). "SUMO and ubiquitin paths converge". Biochemical Society Transactions. 38 (Pt 1): 34–9. doi:10.1042/BST0380034. PMID 20074031.
- Wilkinson KA, Henley JM (May 2010). "Mechanisms, regulation and consequences of protein SUMOylation". The Biochemical Journal. 428 (2): 133–45. doi:10.1042/BJ20100158. PMC 3310159. PMID 20462400.
- Mukhopadhyay D, Riezman H (January 2007). "Proteasome-independent functions of ubiquitin in endocytosis and signaling". Science. 315 (5809): 201–5. Bibcode:2007Sci...315..201M. doi:10.1126/science.1127085. PMID 17218518. S2CID 35434448.
- Enchev RI, Schulman BA, Peter M (January 2015). "Protein neddylation: beyond cullin-RING ligases". Nature Reviews. Molecular Cell Biology. 16 (1): 30–44. doi:10.1038/nrm3919. PMC 5131867. PMID 25531226.
- Shpilka T, Mizushima N, Elazar Z (May 2012). "Ubiquitin-like proteins and autophagy at a glance". Journal of Cell Science. 125 (Pt 10): 2343–8. doi:10.1242/jcs.093757. PMID 22736434.
- Hochstrasser M (August 2000). "Evolution and function of ubiquitin-like protein-conjugation systems". Nature Cell Biology. 2 (8): E153-7. doi:10.1038/35019643. PMID 10934491. S2CID 29557235.
- Wang F, Liu M, Qiu R, Ji C (August 2011). "The dual role of ubiquitin-like protein Urm1 as a protein modifier and sulfur carrier". Protein & Cell. 2 (8): 612–9. doi:10.1007/s13238-011-1074-6. PMC 4875326. PMID 21904977.