Enterochromaffin cell
Enterochromaffin (EC) cells (also known as Kulchitsky cells) are a type of enteroendocrine cell, and neuroendocrine cell. They reside alongside the epithelium lining the lumen of the digestive tract and play a crucial role in gastrointestinal regulation, particularly intestinal motility and secretion.[1] They were discovered by Nikolai Kulchitsky.[2]
| Enterochromaffin cell | |
|---|---|
 | |
| Details | |
| Identifiers | |
| Latin | endocrinocytus EC |
| MeSH | D004759 |
| TH | H3.04.02.0.00029 |
| Anatomical terms of microanatomy | |
EC cells modulate neuron signalling in the enteric nervous system (ENS) via the secretion of the neurotransmitter serotonin and other peptides. As enteric afferent and efferent nerves do not protrude into the intestinal lumen, EC cells act as a form of sensory transduction.[1] Serotonin in the ENS acts in synergy with other digestive hormones to regulate sensory and motor gastrointestinal reflexes. EC cells respond to both chemical and neurological stimuli. They are also reactive to mechanosensation, which is the case in the peristaltic reflex of the gut, and can be stimulated by a bolus moving through the bowel. Upon activation, EC cells release serotonin to act upon serotonin receptors on ENS neurons. Dependent on concentration, serotonin can then modulate peristaltic contraction and secretion through activation of smooth muscle and glands, respectively.[3]
Pulmonary neuroendocrine cells in the respiratory tract are known as bronchial Kulchitsky cells.[4]
Structure
EC cells are small polygonal cells located in the crypts between intestinal villi. They are discriminated from other cells of the gastrointestinal epithelial crypts by the presence of basally located granulations that contain serotonin and other peptides. Ultrastructurally, these granules are reported to vary in size and shape and are considered pleomorphic.[5]
Most EC cells communicate with the lumen of intestinal crypts through apical microvilli (protrusions) and are referred to as “open”. A proportion of EC cells do not protrude into the crypt lumen and are subsequently referred to as ‘closed’.[6] EC cells typically extend down to the basal lamina with cytoplasmic extensions known to pass through the connective tissue and neighbouring glands. Tissue beneath EC cells typically contains abundant fenestrated capillaries, lymph vessels and small unmyelinated nerve fibres. Secreted serotonin can either be taken up into residing vessels (transported in the blood by platelets) or act on nerve synaptic terminals.[5]
Distribution
EC cells are found aggregated in specific locations throughout the gastrointestinal tract, predominantly in the small intestine, colon and appendix.[7] The proportion of high-density cell populations varies between species attributed to differences in dietary requirements and physiological characteristics.[8]
Enterochromaffin-like cells
Enterochromaffin-like (ECL) cells are a population of cells that are found in the gastric pits of the stomach luminal epithelium and secrete histamine. In response to gastrin released by neighbouring G-cells, secreted histamine from ECL cells acts on parietal cells to stimulate the release of gastric acid. The presence of ECL cells is critical in regulation of endocrine-induced gastric acid secretion.[9] ECL cells histologically appear similar to EC cells and are hence named as such. They are however a different cell type and do not possess any serotonin synthesizing mechanisms.
Development
In developing chick embryos, EC cells have been found in biopsies of developing GIT tissue before the migration of neural crest cells. Whilst EC cells have neuroendocrine properties and are similar chemically and histologically to cells of the adrenal medulla they are not derivatives of the neural crest and do not share a similar cell progenitor.[10] EC cells are believed to be derived from endodermal origins and are descended from the stem cells that form other epithelial cell types of the gastrointestinal lumen.[11]
Function
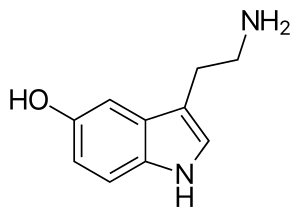
The primary function of EC cells is to synthesise and secrete serotonin for modulation of gastrointestinal neurons. Serotonin, also named 5-hydroxytryptamine (5HT), can be classified as a hormone, neurotransmitter and a mitogen. It is primarily known for its role in the central nervous system but plays an important role in the periphery, with the largest endogenous pool of serotonin residing in the gut (90% of endogenous store). In the ENS, serotonin is an essential modulator of sensory transduction and mucus secretion. Release of serotonin from EC cells can be triggered by a multitude of stimuli, particularly luminal distension, parasympathetic innervation or changes in osmotic concentrations in intestinal contents.[12]
Serotonin synthesis
The synthesis of 5-HT, in EC cells, is catalyzed by the enzyme tryptophan hydroxylase 1 (TpH1) from the amino acid L-tryptophan. The reaction proceeds in two stages with an initial rate limiting step involving the conversion of L-tryptophan to 5-hydroxytryptophan (5-HTP). Following conversion to 5-HTP, the non-rate limiting L-amino acid decarboxylase converts 5-HTP to 5-HT by decarboxylation. Following synthesis, 5-HT is then stored in vesicles by vesicular monoamine transporter 1 close to the basal margin of the cell for eventual secretion.[1]
Release of the vesicles occurs after chemical, neurological or mechanical stimulation of the EC cells and is predominantly calcium dependent, suggesting excretion via exocytosis. The combined effect of increased calcium flux and a liberation of stored calcium within the cell changes the cell potential triggering release of the 5-HT vesicles.[13] The vesicles pass from the basal margin into the surrounding lamina propria for interaction with nearby nerve synapses, lymph and blood vessels.
The serotonin synthesised by EC cells is predominately exocytosed from the basal border, but is also known to be apically secreted into the lumen of the gut and can be present in faecal samples. Secreted 5-HT acts on different receptor subtypes found localised in cells in the gastrointestinal epithelium, smooth muscle and connective tissue with responsiveness dependent on the concentration of the secreted hormone.[5]
The primary effect of serotonin involves the increase in peristaltic contraction through its effects on both ENS neurons and smooth muscle. 5-HT also activates a neural secretory response, whereby binding at 5-HT1P receptors on myenteric neurons triggers a signalling cascade in the submucosal plexus. This results in the release of acetylcholine to initiate secretion from the gut mucosa via release of chloride ions.[14]
Clinical significance
Irritable bowel syndrome
Irritable bowel syndrome (IBS) is a diverse condition associated with chronic bowel discomfort and abdominal pain that ranges in severity between patients. Abnormal concentrations of serotonin have been associated with IBS, predominantly increased concentrations intensifying gastrointestinal motility and mucosal secretions from the gut mucosa. Severe IBS often manifests as either chronic constipation or chronic diarrhoea, and abnormal EC cell populations have been correlated with both conditions. In patients suffering post-infectious IBS, rectal biopsies have shown a dramatic increase in populations of EC cells associated with diarrhoeal symptoms.[15]
Likewise, reduced populations of EC cells in patients suffering chronic constipation have been observed, indicating a lack of 5-HT, and therefore decreased GI motility and secretion. Ongoing research indicates that abnormal EC cell populations, and therefore 5-HT signalling, may significantly contribute to gastrointestinal dysfunction. Treatment using 5-HT-receptor agonists for patients with functional constipation have shown some effectiveness in achieving normal GI functionality.[16]
Carcinoid syndrome
Carcinoid syndrome is a rare condition characterized by an abnormal increase in circulating biologically active hormones, largely serotonin, with early symptoms involving diarrhea, abdominal cramping and episodic flushing.[6] Excess circulating serotonin is usually manufactured by EC-cell-originated carcinoid tumors in the small bowel or appendix. Tumors are slow growing, but can metastasise to the liver if aggressive. They can also be present at other sites, particularly the lung and stomach.[17]
History
The name ‘enterochromaffin’ comes from the Greek word “enteron” (ἔντερον), in relation to intestines, and “chromaffin” as a grouping of the words chromium and affinity, as they can be visualised by staining with chromium salts. Similarly named, chromaffin cells (of the adrenal medulla) share this characteristic and are histologically similar to EC cells. Their embryological origins, however, are quite different, nor do they possess similar functions.
References
- Bertrand, Paul P.; Bertrand, Rebecca L. (2010). "Serotonin release and uptake in the gastrointestinal tract". Autonomic Neuroscience. 153 (1–2): 47–57. doi:10.1016/j.autneu.2009.08.002. PMID 19729349. S2CID 25355291.
- Drozdov, I.; Modlin, I. M; Kidd, M.; Goloubinov, V. V (2009). "Nikolai Konstantinovich Kulchitsky (1856-1925)". Journal of Medical Biography. 17 (1): 47–54. doi:10.1258/jmb.2008.008038. PMID 19190200. S2CID 25289790.
- Mawe, Gary M.; Hoffman, Jill M. (2013). "Serotonin signalling in the gut—functions, dysfunctions and therapeutic targets". Nature Reviews Gastroenterology & Hepatology. 10 (8): 473–86. doi:10.1038/nrgastro.2013.105. PMC 4048923. PMID 23797870.
- Becker, KL; Silva, OL (July 1981). "Hypothesis: the bronchial Kulchitsky (K) cell as a source of humoral biologic activity". Medical Hypotheses. 7 (7): 943–9. doi:10.1016/0306-9877(81)90049-9. PMID 6270516.
- Wade, P.R.; Westfall, Janea. (1985). "Ultrastructure of enterochromaffin cells and associated neural and vascular elements in the mouse duodenum". Cell and Tissue Research. 241 (3): 557–63. doi:10.1007/BF00214576. PMID 4028141. S2CID 33773512.
- Gustafsson, Björn I (2005). The Serotonin Producing Enterochromaffin Cell, and Effects of Hyperserotoninemia on Heart and Bone (PhD Thesis). Det medisinske fakultet. hdl:11250/263149. ISBN 9788247173121.
- Graeme-Cook, Fiona (2009). "Neuroendocrine Tumors of the GI Tract and Appendix". In Odze, Robert D.; Goldblum, John R. (eds.). Surgical Pathology of the GI Tract, Liver, Biliary Tract, and Pancreas. pp. 653–80. doi:10.1016/B978-141604059-0.50028-X. ISBN 978-1-4160-4059-0.
- Al-Tikriti, M. S.; Khamas, W.; Chebolu, S.; Darmani, N. A. (2012). "Distribution of Serotonin-Immunoreactive Enterochromaffin Cells in the Gastrointestinal Tract of the Least Shrew" (PDF). Int. J. Morphol. 30 (3): 916–23. doi:10.4067/s0717-95022012000300025.
- Prinz, C; Zanner, R; Gerhard, M; Mahr, S; Neumayer, N; Höhne-Zell, B; Gratzl, M (1999). "The mechanism of histamine secretion from gastric enterochromaffin-like cells". The American Journal of Physiology. 277 (5 Pt 1): C845–55. doi:10.1152/ajpcell.1999.277.5.C845. PMID 10564076.
- Andrew, A (1974). "Further evidence that enterochromaffin cells are not derived from the neural crest". Journal of Embryology and Experimental Morphology. 31 (3): 589–98. PMID 4448939.
- Thompson, M; Fleming, K. A.; Evans, D. J.; Fundele, R; Surani, M. A.; Wright, N. A. (1990). "Gastric endocrine cells share a clonal origin with other gut cell lineages". Development. 110 (2): 477–81. PMID 2133551.
- Manocha, Marcus; Khan, Waliul I (2012). "Serotonin and GI Disorders: An Update on Clinical and Experimental Studies". Clinical and Translational Gastroenterology. 3 (4): e13. doi:10.1038/ctg.2012.8. PMC 3365677. PMID 23238212.
- Racké, K.; Reimann, A.; Schwörer, H.; Kilbinger, H. (1995). "Regulation of 5-HT release from enterochromaffin cells". Behavioural Brain Research. 73 (1–2): 83–7. doi:10.1016/0166-4328(96)00075-7. PMID 8788482. S2CID 3948073.
- Sidhu, M; Cooke, H. J. (1995). "Role for 5-HT and ACh in submucosal reflexes mediating colonic secretion". The American Journal of Physiology. 269 (3 Pt 1): G346–51. doi:10.1152/ajpgi.1995.269.3.G346. PMID 7573444.
- Lee, Kwang Jae; Kim, Yeong Bae; Kim, Jang Hee; Kwon, Hoek Chun; Kim, Dong Kyu; Cho, Sung Won (2008). "The alteration of enterochromaffin cell, mast cell, and lamina propria T lymphocyte numbers in irritable bowel syndrome and its relationship with psychological factors". Journal of Gastroenterology and Hepatology. 23 (11): 1689–94. doi:10.1111/j.1440-1746.2008.05574.x. PMID 19120860. S2CID 205463559.
- m. El-Salhy, ö. Norrgård; Norrgård, O; Spinnell, S (2009). "Abnormal Colonic Endocrine Cells in Patients with Chronic Idiopathic Slow-Transit Constipation". Scandinavian Journal of Gastroenterology. 34 (10): 1007–11. doi:10.1080/003655299750025110. PMID 10563671.
- Berge, Thorbjörn; Linell, Folke (2009). "Carcinoid Tumours: Frequency in a Defined Population During a 12-Year-Period". APMIS. 84A (4): 322–30. doi:10.1111/j.1699-0463.1976.tb00122.x. PMID 961424.