Lokiarchaeota
Lokiarchaeota is a proposed phylum of the Archaea.[1] The phylum includes all members of the group previously named Deep Sea Archaeal Group (DSAG), also known as Marine Benthic Group B (MBG-B). Lokiarchaeota is part of the superphylum Asgard containing the phyla: Lokiarchaeota, Thorarchaeota, Odinarchaeota, Heimdallarchaeota, and Helarchaeota.[2] A phylogenetic analysis disclosed a monophyletic grouping of the Lokiarchaeota with the eukaryotes. The analysis revealed several genes with cell membrane-related functions. The presence of such genes support the hypothesis of an archaeal host for the emergence of the eukaryotes; the eocyte-like scenarios.
| Lokiarchaeota | |
|---|---|
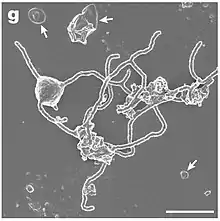 | |
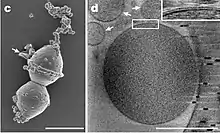
| SEM image of Candidatus Prometheoarchaeum syntrophicum | |
| Scientific classification | |
| Domain: | |
| Kingdom: | |
| Superphylum: | |
| Phylum: | "Lokiarchaeota" Spang et al. 2015 |
| Class: | "Lokiarchaeia" corrig. Spang et al. 2015 |
| Order: | "Lokiarchaeales" Spang et al. 2015 |
| Family: | "Lokiarchaeaceae" Vanwonterghem et al. 2016 |
| Type species | |
| "Ca. Prometheoarchaeum syntrophicum" Imachi et al. 2020 | |
| Genus | |
| |
Lokiarchaeota was introduced in 2015 after the identification of a candidate genome in a metagenomic analysis of a mid-oceanic sediment sample. This analysis suggests the existence of a genus of unicellular life dubbed Lokiarchaeum. The sample was taken near a hydrothermal vent at a vent field known as Loki's Castle located at the bend between Mohns/Knipovich ridge in the Arctic Ocean.[3]
Discovery
Sediments from a gravity core taken in 2010 in the rift valley on the Knipovich ridge in the Arctic Ocean, near the so-called Loki's Castle hydrothermal vent site, were analysed. Specific sediment horizons, previously shown to contain high abundances of novel archaeal lineages[4][5] were subjected to metagenomic analysis. Due to the low density of cells in the sediment, the resulting genetic sequence does not come from an isolated cell, as would be the case in conventional analysis, but is rather a combination of genetic fragments.[6] The result was a 92% complete, 1.4 fold-redundant composite genome named Lokiarchaeum.[3]
The metagenomic analysis determined the presence of an organism's genome in the sample.[3] However, the organism itself was not cultured until years later, with a Japanese group first reporting isolation and cultivation of a Lokiarchaeota strain in 2019.[7] Since this initial cultivation of Lokiarchaeota, members of the phylum have been reported in a diverse range of habitats. Advances in both long and short-read technologies for DNA sequencing have also aided in the recovery and identification of Lokiarchaeota from microbial samples.[2]
The Lokiarchaeota phylum was proposed based on phylogenetic analyses using a set of highly conserved protein-coding genes.[3] Through a reference to the hydrothermal vent complex from which the first genome sample originated, the name refers to Loki, the Norse shape-shifting god.[6] The Loki of literature has been described as "a staggeringly complex, confusing, and ambivalent figure who has been the catalyst of countless unresolved scholarly controversies",[8] an analogy to the role of Lokiarchaeota in debates about the origin of eukaryotes.[3]
Description
The Lokiarchaeum composite genome consists of 5,381 protein coding genes. Of these, roughly 32% do not correspond to any known protein, 26% closely resemble archaeal proteins, and 29% correspond to bacterial proteins. This situation is consistent with: (i) proteins from a novel phylum (with few close relatives, or none) being difficult to assign to their correct domain; and (ii) existing research that suggests there has been significant inter-domain gene transfer between bacteria and Archaea.
A small, but significant portion of the proteins (175, 3.3%) that the recovered genes code for are very similar to eukaryotic proteins. These proteins included homologs of cytoskeleton proteins, GTPases, and the oligosaccharyltransferase (OST) protein complex. Homologues for components of the endosomal sorting complex required for transport and the ubiquitin protein modifier system were also identified in Lokiarchaeota genome analysis.[2] Sample contamination is an unlikely explanation for the unusual proteins because the recovered genes were always flanked by prokaryotic genes and no genes of known eukaryotic origin were detected in the metagenome from which the composite genome was extracted. Further, previous phylogenetic analysis suggested the genes in question had their origin at the base of the eukaryotic clades.[3]
In eukaryotes, the function of these shared proteins include cell membrane deformation, cell shape formation, and a dynamic protein cytoskeleton.[3][9][10] Eukaryotic protein functions found in Lokiarchaeota also include intracellular transport mechanisms.[11] It is inferred then that Lokiarchaeum may have some of these abilities.[3] Another shared protein, actin, is essential for phagocytosis in eukaryotes.[6][9] Phagocytosis is the ability to engulf and consume another particle; such ability would facilitate the endosymbiotic origin of mitochondria and chloroplasts, which is a key difference between prokaryotes and eukaryotes.[3] The presence of actin proteins and intracellular transport mechanisms provides evidence for the common ancestry between ancient Lokiarchaeota and eukarya.[11]
Evolutionary significance

A comparative analysis of the Lokiarchaeum genome against known genomes resulted in a phylogenetic tree that showed a monophyletic group composed of the Lokiarchaeota and the eukaryotes,[12] supporting an archaeal host or eocyte-like scenarios for the emergence of the eukaryotes.[13][14][15] The repertoire of membrane-related functions of Lokiarchaeum suggests that the common ancestor to the eukaryotes might be an intermediate step between the prokaryotic cells, devoid of subcellular structures, and the eukaryotic cells, which harbor many organelles.[3]
Carl Woese's three-domain system classifies cellular life into three domains: archaea, bacteria, and eukaryotes; the last being characterised by large, highly evolved cells, containing mitochondria, which help the cells produce ATP (adenosine triphosphate, the energy currency of the cell), and a membrane-bound nucleus containing nucleic acids. Protozoa and all multicellular organisms such as animals, fungi, and plants are eukaryotes.
The bacteria and archaea are thought to be the most ancient of lineages,[16] as fossil strata bearing the chemical signature of archaeal lipids have been dated back to 3.8 billion years ago.[17] The eukaryotes include all complex cells and almost all multicellular organisms. They are thought to have evolved between 1.6 and 2.1 billion years ago.[18] While the evolution of eukaryotes is considered to be an event of great evolutionary significance, no intermediate forms or "missing links" had been discovered previously. In this context, the discovery of Lokiarchaeum, with some but not all of the characteristics of eukaryotes, provides evidence on the transition from archaea to eukaryotes.[19] Lokiarchaeota and the eukaryotes probably share a common ancestor, and if so, diverged roughly two billion years ago. Evidence for common ancestry, rather than an evolutionary shift from Lokiarchaeota to eukaryotes, is found in analysis of fold superfamilies (FSFs). Fold super families are evolutionarily defined domains of protein structure. It is estimated that there are around 2500 total FSFs found in nature.[11] Utilization of Venn diagrams allowed researchers to depict distributions of FSFs of those that were shared by Archaea and Eukarya, as well as those unique to their respective kingdoms. The addition of Lokiarchaeum into the Venn groups created from an initial genomic census only added 10 FSFs to Archaea. The addition of Lokiarchaeum also only contributed to a decrease of two FSFs previously unique to Eukarya. There were still 284 FSFs found exclusively in Eukarya. Lokiarchaeota’s limited impact in changing the Venn distribution of FSFs demonstrates the lack of genes that could be traced to a common ancestor with Eukaryotes. Rather, Eukaryotic genes present in bacterial and archaeal organisms are hypothesized to be from horizontal transfer from an early ancestor of modern eukaryotes.[11] This putative ancestor possessed crucial "starter" genes that enabled increased cellular complexity. This common ancestor, or a relative, eventually led to the evolution of eukaryotes.[6]
In 2020, a Japanese research group reported culturing a strain of Lokiarchaeota in the laboratory.[20][21][7] This strain, currently named Candidatus Prometheoarchaeum syntrophicum strain MK-D1, was observed in syntrophic association with two hydrogen-consuming microbes: a sulfate-reducing bacteria of the genus Halodesulfovibrio and a methanogen of the genus Methanogenium. The MK-D1 organism produces hydrogen as a metabolic byproduct, which is then consumed by the symbiotic syntrophs. MK-D1 also seems to organize its external membrane into complex structures using genes shared with eukaryotes. While association with alphaproteobacteria (from which mitochondria are thought to descend) was not observed, these features suggest that MK-D1 and its syntrophs may represent an extant example of archaea-bacteria symbiosis similar to that which gave rise to eukaryotes. To date, this is the only Asgard archaeon for which a co-culture is available.
Physiological Traits
Lokiarachaeota is known to have a tetrahydromethanopterin-dependent Wood-Ljundahl (H4MPT-WL) pathway. This pathway contains a series of biochemical reactions aiding in inorganic carbon utilization. In Lokiarchaeota, the WLP is thought to be acetogenic, due to lacking the gene methyl-CoM reductase necessary for methanogenesis.[2]
Analysis of Lokiarchaeon genes also showed the expression of protein-encoding open reading frames (ORFs) involving the metabolism of sugars and proteins. However, these metabolic activities vary between subgroups of Lokiarchaeota. While Lokiarchaeota subgroups have similar genetic information, differences in metabolic abilities explain their respective ecological niches.[22]
Two major subgroups of the Lokiarachaeota phylum are Loki-2 and Loki-3. Incubations of these two subgroups from Helgoland mud sediments were analyzed through RNA and DNA stable isotope probing to understand their respective carbon metabolisms.[22] Loki-3 were found to be active in both organic carbon utilization and the degradation of aromatic compounds. The Loki-3 subgroup was not found to utilize proteins or short chain fatty acids, even though genes for amino acid degradation were present in both subgroups. Loki-2 was found to utilize protein, as seen through activity in when proteins were provided in Loki-2 incubations. Due to the greater carbon utilization pathways of Loki-3, the subgroup is found in a more diverse range of marine sediments than Loki-2.[22]
See also
References
- Lambert, Jonathan (9 August 2019). "Scientists glimpse oddball microbe that could help explain rise of complex life - 'Lokiarchaea', previously known only from DNA, is isolated and grown in culture". Nature. 572 (7769): 294. Bibcode:2019Natur.572..294L. doi:10.1038/d41586-019-02430-w. PMID 31409927.
- Caceres, Eva F.; Lewis, William H.; Homa, Felix; Martin, Tom; Schramm, Andreas; Kjeldsen, Kasper U.; Ettema, Thijs J. G. (2019-12-18). "Near-complete Lokiarchaeota genomes from complex environmental samples using long and short read metagenomic analyses": 2019.12.17.879148. doi:10.1101/2019.12.17.879148. S2CID 213982145.
{{cite journal}}: Cite journal requires|journal=(help) - Spang, Anja; Saw, Jimmy H.; Jørgensen, Steffen L.; Zaremba-Niedzwiedzka, Katarzyna; Martijn, Joran; Lind, Anders E.; van Eijk, Roel; Schleper, Christa; Guy, Lionel; Ettema, Thijs J. G. (2015). "Complex archaea that bridge the gap between prokaryotes and eukaryotes". Nature. 521 (7551): 173–179. Bibcode:2015Natur.521..173S. doi:10.1038/nature14447. ISSN 0028-0836. PMC 4444528. PMID 25945739.
- Jørgensen, Steffen Leth; Hannisdal, Bjarte; Lanzen, Anders; Baumberger, Tamara; Flesland, Kristin; Fonseca, Rita; Øvreås, Lise; Steen, Ida H; Thorseth, Ingunn H; Pedersen, Rolf B; Schleper, Christa (September 5, 2012). "Correlating microbial community profiles with geochemical data in highly stratified sediments from the Arctic Mid-Ocean Ridge". PNAS. 109 (42): E2846–55. doi:10.1073/pnas.1207574109. PMC 3479504. PMID 23027979.
- Jørgensen, Steffen Leth; Thorseth, Ingunn H; Pedersen, Rolf B; Baumberger, Tamara; Schleper, Christa (October 4, 2013). "Quantitative and phylogenetic study of the Deep Sea Archaeal Group in sediments of the Arctic mid-ocean spreading ridge". Frontiers in Microbiology. 4: 299. doi:10.3389/fmicb.2013.00299. PMC 3790079. PMID 24109477.
- Rincon, Paul (May 6, 2015). "Newly found microbe is close relative of complex life". BBC. Retrieved May 9, 2015.
- Imachi, Hiroyuki; et al. (15 January 2020). "Isolation of an archaeon at the prokaryote–eukaryote interface". Nature. 577 (7791): 519–525. Bibcode:2020Natur.577..519I. bioRxiv 10.1101/726976. doi:10.1038/s41586-019-1916-6. PMC 7015854. PMID 31942073.
- von Schnurbein, Stefanie (November 2000). "The Function of Loki in Snorri Sturluson's "Edda"". History of Religions. 40 (2): 109–124. doi:10.1086/463618.
- Ghoshdastider U, Jiang S, Popp D, Robinson RC (2015). "In search of the primordial actin filament". Proc Natl Acad Sci U S A. 112 (30): 9150–1. doi:10.1073/pnas.1511568112. PMC 4522752. PMID 26178194.
{{cite journal}}: CS1 maint: uses authors parameter (link) - Khan, Amina (May 6, 2015). "Meet Loki, your closest-known prokaryote relative". LA Times. Retrieved May 9, 2015.
- Nasir, Arshan; Kim, Kyung Mo; Caetano-Anollés, Gustavo (2015-08-01). "Lokiarchaeota: eukaryote-like missing links from microbial dark matter?". Trends in Microbiology. 23 (8): 448–450. doi:10.1016/j.tim.2015.06.001. ISSN 0966-842X. PMID 26112912.
- Maximillan, Ludwig (27 December 2019). "Evolution: A revelatory relationship". Phys.org. Retrieved 28 December 2019.
- Embley, T. Martin; Martin, William (2006). "Eukaryotic evolution, changes and challenges". Nature. 440 (7084): 623–630. Bibcode:2006Natur.440..623E. doi:10.1038/nature04546. ISSN 0028-0836. PMID 16572163. S2CID 4396543.
- Lake, James A. (1988). "Origin of the eukaryotic nucleus determined by rate-invariant analysis of rRNA sequences". Nature. 331 (6152): 184–186. Bibcode:1988Natur.331..184L. doi:10.1038/331184a0. ISSN 0028-0836. PMID 3340165. S2CID 4368082.
- Guy, Lionel; Ettema, Thijs J.G. (2011). "The archaeal 'TACK' superphylum and the origin of eukaryotes". Trends in Microbiology. 19 (12): 580–587. doi:10.1016/j.tim.2011.09.002. ISSN 0966-842X. PMID 22018741.
- Wang, Minglei; Yafremava, Liudmila S.; Caetano-Anollés, Derek; Mittenthal, Jay E.; Caetano-Anollés, Gustavo (2007). "Reductive evolution of architectural repertoires in proteomes and the birth of the tripartite world". Genome Research. 17 (11): 1572–1585. doi:10.1101/gr.6454307. PMC 2045140. PMID 17908824.
- Hahn, Jürgen; Haug, Pat (1986). "Traces of Archaebacteria in ancient sediments". Systematic and Applied Microbiology. 7 (Archaebacteria '85 Proceedings): 178–83. doi:10.1016/S0723-2020(86)80002-9.
- Knoll, Andrew H.; Javaux, E. J.; Hewitt, D.; Cohen, P. (29 June 2006). "Eukaryotic organisms in Proterozoic oceans". Philosophical Transactions of the Royal Society B. 361 (1470): 1023–38. doi:10.1098/rstb.2006.1843. PMC 1578724. PMID 16754612.
- Zimmer, Carl (6 May 2015). "Under the Sea, a Missing Link in the Evolution of Complex Cells". New York Times. Retrieved 8 May 2015.
- Timmer, John (2019-08-07). "We've finally gotten a look at the microbe that might have been our ancestor". Ars Technica. Retrieved 2019-08-08.
- Zimmer, Carl (2020-01-15). "This Strange Microbe May Mark One of Life's Great Leaps". The New York Times. Retrieved 2020-01-15.
- Yin, Xiuran; Cai, Mingwei; Liu, Yang; Zhou, Guowei; Richter-Heitmann, Tim; Aromokeye, David A.; Kulkarni, Ajinkya C.; Nimzyk, Rolf; Cullhed, Henrik; Zhou, Zhichao; Pan, Jie (Mar 2021). "Subgroup level differences of physiological activities in marine Lokiarchaeota". The ISME Journal. 15 (3): 848–861. doi:10.1038/s41396-020-00818-5. ISSN 1751-7370. PMC 8027215. PMID 33149207.