Origin of replication
The origin of replication (also called the replication origin) is a particular sequence in a genome at which replication is initiated.[1] Propagation of the genetic material between generations requires timely and accurate duplication of DNA by semiconservative replication prior to cell division to ensure each daughter cell receives the full complement of chromosomes.[2] This can either involve the replication of DNA in living organisms such as prokaryotes and eukaryotes, or that of DNA or RNA in viruses, such as double-stranded RNA viruses.[3] Synthesis of daughter strands starts at discrete sites, termed replication origins, and proceeds in a bidirectional manner until all genomic DNA is replicated. Despite the fundamental nature of these events, organisms have evolved surprisingly divergent strategies that control replication onset.[2] Although the specific replication origin organization structure and recognition varies from species to species, some common characteristics are shared.

History
In the second half of the 19th century, Gregor Mendel's pioneering work on the inheritance of traits in pea plants suggested that specific “factors” (today established as genes) are responsible for transferring organismal traits between generations.[4] Although proteins were initially assumed to serve as the hereditary material, Avery, MacLeod and McCarty established a century later DNA, which had been discovered by Friedrich Miescher, as the carrier of genetic information.[5] These findings paved the way for research uncovering the chemical nature of DNA and the rules for encoding genetic information, and ultimately led to the proposal of the double-helical structure of DNA by Watson and Crick.[6] This three-dimensional model of DNA illuminated potential mechanisms by which the genetic information could be copied in a semiconservative manner prior to cell division, a hypothesis that was later experimentally supported by Meselson and Stahl using isotope incorporation to distinguish parental from newly synthesized DNA.[7][8] The subsequent isolation of DNA polymerases, the enzymes that catalyze the synthesis of new DNA strands, by Kornberg and colleagues pioneered the identification of many different components of the biological DNA replication machinery, first in the bacterial model organism E. coli, but later also in eukaryotic life forms.[2][9]
Features
A key prerequisite for DNA replication is that it must occur with extremely high fidelity and efficiency exactly once per cell cycle to prevent the accumulation of genetic alterations with potentially deleterious consequences for cell survival and organismal viability.[10] Incomplete, erroneous, or untimely DNA replication events can give rise to mutations, chromosomal polyploidy or aneuploidy, and gene copy number variations, each of which in turn can lead to diseases, including cancer.[11][12] To ensure complete and accurate duplication of the entire genome and the correct flow of genetic information to progeny cells, all DNA replication events are not only tightly regulated with cell cycle cues but are also coordinated with other cellular events such as transcription and DNA repair.[2][13][14][15] Additionally, origin sequences commonly have high AT-content across all kingdoms, since repeats of adenine and thymine are easier to separate because their base stacking interactions are not as strong as those of guanine and cytosine.[16]
DNA replication is divided into different stages. During initiation, the replication machineries – termed replisomes – are assembled on DNA in a bidirectional fashion. These assembly loci constitute the start sites of DNA replication or replication origins. In the elongation phase, replisomes travel in opposite directions with the replication forks, unwinding the DNA helix and synthesizing complementary daughter DNA strands using both parental strands as templates. Once replication is complete, specific termination events lead to the disassembly of replisomes. As long as the entire genome is duplicated before cell division, one might assume that the location of replication start sites does not matter; yet, it has been shown that many organisms use preferred genomic regions as origins.[17][18] The necessity to regulate origin location likely arises from the need to coordinate DNA replication with other processes that act on the shared chromatin template to avoid DNA strand breaks and DNA damage.[2][12][15][19][20][21][22][23]
Replicon model
More than five decades ago, Jacob, Brenner, and Cuzin proposed the replicon hypothesis to explain the regulation of chromosomal DNA synthesis in E. coli.[24] The model postulates that a diffusible, trans-acting factor, a so-called initiator, interacts with a cis-acting RNA element, the replicator, to promote replication onset at a nearby origin. Once bound to replicators, initiators (often with the help of co-loader proteins) deposit replicative helicases onto DNA, which subsequently drive the recruitment of additional replisome components and the assembly of the entire replication machinery. The replicator thereby specifies the location of replication initiation events, and the chromosome region that is replicated from a single origin or initiation event is defined as the replicon.[2]
A fundamental feature of the replicon hypothesis is that it relies on positive regulation to control DNA replication onset, which can explain many experimental observations in bacterial and phage systems.[24] For example, it accounts for the failure of extrachromosomal DNAs without origins to replicate when introduced into host cells. It further rationalizes plasmid incompatibilities in E. coli, where certain plasmids destabilize each other's inheritance due to competition for the same molecular initiation machinery.[25] By contrast, a model of negative regulation (analogous to the replicon-operator model for transcription) fails to explain the above findings.[24] Nonetheless, research subsequent to Jacob's, Brenner's and Cuzin's proposal of the replicon model has discovered many additional layers of replication control in bacteria and eukaryotes that comprise both positive and negative regulatory elements, highlighting both the complexity and the importance of restricting DNA replication temporally and spatially.[2][26][27][28]
The concept of the replicator as a genetic entity has proven very useful in the quest to identify replicator DNA sequences and initiator proteins in prokaryotes, and to some extent also in eukaryotes, although the organization and complexity of replicators differ considerably between the domains of life.[29][30] While bacterial genomes typically contain a single replicator that is specified by consensus DNA sequence elements and that controls replication of the entire chromosome, most eukaryotic replicators – with the exception of budding yeast – are not defined at the level of DNA sequence; instead, they appear to be specified combinatorially by local DNA structural and chromatin cues.[31][32][33][34][35][36][37][38][39][40] Eukaryotic chromosomes are also much larger than their bacterial counterparts, raising the need for initiating DNA synthesis from many origins simultaneously to ensure timely replication of the entire genome. Additionally, many more replicative helicases are loaded than activated to initiate replication in a given cell cycle. The context-driven definition of replicators and selection of origins suggests a relaxed replicon model in eukaryotic systems that allows for flexibility in the DNA replication program.[29] Although replicators and origins can be spaced physically apart on chromosomes, they often co-localize or are located in close proximity; for simplicity, we will thus refer to both elements as ‘origins’ throughout this review. Taken together, the discovery and isolation of origin sequences in various organisms represents a significant milestone towards gaining mechanistic understanding of replication initiation. In addition, these accomplishments had profound biotechnological implications for the development of shuttle vectors that can be propagated in bacterial, yeast and mammalian cells.[2][41][42][43]
Bacterial

Most bacterial chromosomes are circular and contain a single origin of chromosomal replication (oriC). Bacterial oriC regions are surprisingly diverse in size (ranging from 250 bp to 2 kbp), sequence, and organization;[45][46] nonetheless, their ability to drive replication onset typically depends on sequence-specific readout of consensus DNA elements by the bacterial initiator, a protein called DnaA.[47][48][49][50] Origins in bacteria are either continuous or bipartite and contain three functional elements that control origin activity: conserved DNA repeats that are specifically recognized by DnaA (called DnaA-boxes), an AT-rich DNA unwinding element (DUE), and binding sites for proteins that help regulate replication initiation.[17][51][52] Interactions of DnaA both with the double-stranded (ds) DnaA-box regions and with single-stranded (ss) DNA in the DUE are important for origin activation and are mediated by different domains in the initiator protein: a Helix-turn-helix (HTH) DNA binding element and an ATPase associated with various cellular activities (AAA+) domain, respectively.[53][54][55][56][57][58][59] While the sequence, number, and arrangement of origin-associated DnaA-boxes vary throughout the bacterial kingdom, their specific positioning and spacing in a given species are critical for oriC function and for productive initiation complex formation.[2][45][46][60][61][62][63][64]
Among bacteria, E. coli is a particularly powerful model system to study the organization, recognition, and activation mechanism of replication origins. E. coli oriC comprises an approximately ~260 bp region containing four types of initiator binding elements that differ in their affinities for DnaA and their dependencies on the co-factor ATP. DnaA-boxes R1, R2, and R4 constitute high-affinity sites that are bound by the HTH domain of DnaA irrespective of the nucleotide-binding state of the initiator.[47][65][66][67][68][69] By contrast, the I, τ, and C-sites, which are interspersed between the R-sites, are low-affinity DnaA-boxes and associate preferentially with ATP-bound DnaA, although ADP-DnaA can substitute for ATP-DnaA under certain conditions.[70][71][72][63] Binding of the HTH domains to the high- and low-affinity DnaA recognition elements promotes ATP-dependent higher-order oligomerization of DnaA's AAA+ modules into a right-handed filament that wraps duplex DNA around its outer surface, thereby generating superhelical torsion that facilitates melting of the adjacent AT-rich DUE.[53][73][74][75] DNA strand separation is additionally aided by direct interactions of DnaA's AAA+ ATPase domain with triplet repeats, so-called DnaA-trios, in the proximal DUE region.[76] The engagement of single-stranded trinucleotide segments by the initiator filament stretches DNA and stabilizes the initiation bubble by preventing reannealing.[57] The DnaA-trio origin element is conserved in many bacterial species, indicating it is a key element for origin function.[76] After melting, the DUE provides an entry site for the E. coli replicative helicase DnaB, which is deposited onto each of the single DNA strands by its loader protein DnaC.[2]
Although the different DNA binding activities of DnaA have been extensively studied biochemically and various apo, ssDNA-, or dsDNA-bound structures have been determined,[56][57][58][74] the exact architecture of the higher-order DnaA-oriC initiation assembly remains unclear. Two models have been proposed to explain the organization of essential origin elements and DnaA-mediated oriC melting. The two-state model assumes a continuous DnaA filament that switches from a dsDNA binding mode (the organizing complex) to an ssDNA binding mode in the DUE (the melting complex).[74][77] By contrast, in the loop-back model, the DNA is sharply bent in oriC and folds back onto the initiator filament so that DnaA protomers simultaneously engage double- and single-stranded DNA regions.[78] Elucidating how exactly oriC DNA is organized by DnaA remains thus an important task for future studies. Insights into initiation complex architecture will help explain not only how origin DNA is melted, but also how a replicative helicase is loaded directionally onto each of the exposed single DNA strands in the unwound DUE, and how these events are aided by interactions of the helicase with the initiator and specific loader proteins.[2]
Archaeal

Archaeal replication origins share some but not all of the organizational features of bacterial oriC. Unlike bacteria, Archaea often initiate replication from multiple origins per chromosome (one to four have been reported);[79][80][81][82][83][84][85][86][46] yet, archaeal origins also bear specialized sequence regions that control origin function.[87][88][89] These elements include both DNA sequence-specific origin recognition boxes (ORBs or miniORBs) and an AT-rich DUE that is flanked by one or several ORB regions.[85][90] ORB elements display a considerable degree of diversity in terms of their number, arrangement, and sequence, both among different archaeal species and among different origins within in a single species.[80][85][91] An additional degree of complexity is introduced by the initiator, Orc1/Cdc6 in archaea, which binds to ORB regions. Archaeal genomes typically encode multiple paralogs of Orc1/Cdc6 that vary substantially in their affinities for distinct ORB elements and that differentially contribute to origin activities.[85][92][93][94] In Sulfolobus solfataricus, for example, three chromosomal origins have been mapped (oriC1, oriC2, and oriC3), and biochemical studies have revealed complex binding patterns of initiators at these sites.[85][86][95][96] The cognate initiator for oriC1 is Orc1-1, which associates with several ORBs at this origin.[85][93] OriC2 and oriC3 are bound by both Orc1-1 and Orc1-3.[85][93][96] Conversely, a third paralog, Orc1-2, footprints at all three origins but has been postulated to negatively regulate replication initiation.[85][96] Additionally, the WhiP protein, an initiator unrelated to Orc1/Cdc6, has been shown to bind all origins as well and to drive origin activity of oriC3 in the closely related Sulfolobus islandicus.[93][95] Because archaeal origins often contain several adjacent ORB elements, multiple Orc1/Cdc6 paralogs can be simultaneously recruited to an origin and oligomerize in some instances;[94][97] however, in contrast to bacterial DnaA, formation of a higher-order initiator assembly does not appear to be a general prerequisite for origin function in the archaeal domain.[2]
Structural studies have provided insights into how archaeal Orc1/Cdc6 recognizes ORB elements and remodels origin DNA.[97][98] Orc1/Cdc6 paralogs are two-domain proteins and are composed of a AAA+ ATPase module fused to a C-terminal winged-helix fold.[99][100][101] DNA-complexed structures of Orc1/Cdc6 revealed that ORBs are bound by an Orc1/Cdc6 monomer despite the presence of inverted repeat sequences within ORB elements.[97][98] Both the ATPase and winged-helix regions interact with the DNA duplex but contact the palindromic ORB repeat sequence asymmetrically, which orients Orc1/Cdc6 in a specific direction on the repeat.[97][98] Interestingly, the DUE-flanking ORB or miniORB elements often have opposite polarities,[80][85][94][102][103] which predicts that the AAA+ lid subdomains and the winged-helix domains of Orc1/Cdc6 are positioned on either side of the DUE in a manner where they face each other.[97][98] Since both regions of Orc1/Cdc6 associate with a minichromosome maintenance (MCM) replicative helicase,[104][105] this specific arrangement of ORB elements and Orc1/Cdc6 is likely important for loading two MCM complexes symmetrically onto the DUE.[85] Surprisingly, while the ORB DNA sequence determines the directionality of Orc1/Cdc6 binding, the initiator makes relatively few sequence-specific contacts with DNA.[97][98] However, Orc1/Cdc6 severely underwinds and bends DNA, suggesting that it relies on a mix of both DNA sequence and context-dependent DNA structural features to recognize origins.[97][98][106] Notably, base pairing is maintained in the distorted DNA duplex upon Orc1/Cdc6 binding in the crystal structures,[97][98] whereas biochemical studies have yielded contradictory findings as to whether archaeal initiators can melt DNA similarly to bacterial DnaA.[93][94][107] Although the evolutionary kinship of archaeal and eukaryotic initiators and replicative helicases indicates that archaeal MCM is likely loaded onto duplex DNA (see next section), the temporal order of origin melting and helicase loading, as well as the mechanism for origin DNA melting, in archaeal systems remains therefore to be clearly established. Likewise, how exactly the MCM helicase is loaded onto DNA needs to be addressed in future studies.[2]
Eukaryotic
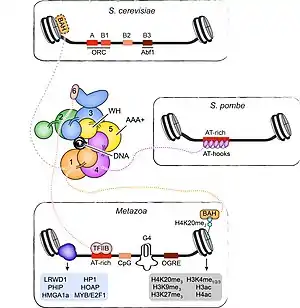
Origin organization, specification, and activation in eukaryotes are more complex than in bacterial or archaeal domains and significantly deviate from the paradigm established for prokaryotic replication initiation. The large genome sizes of eukaryotic cells, which range from 12 Mbp in S. cerevisiae to 3 Gbp in humans, necessitates that DNA replication starts at several hundred (in budding yeast) to tens of thousands (in humans) origins to complete DNA replication of all chromosomes during each cell cycle.[27][36] With the exception of S. cerevisiae and related Saccharomycotina species, eukaryotic origins do not contain consensus DNA sequence elements but their location is influenced by contextual cues such as local DNA topology, DNA structural features, and chromatin environment.[29][35][37] Nonetheless, eukaryotic origin function still relies on a conserved initiator protein complex to load replicative helicases onto DNA during the late M and G1 phases of the cell cycle, a step known as origin licensing.[110] In contrast to their bacterial counterparts, replicative helicases in eukaryotes are loaded onto origin duplex DNA in an inactive, double-hexameric form and only a subset of them (10-20% in mammalian cells) is activated during any given S phase, events that are referred to as origin firing.[111][112][113] The location of active eukaryotic origins is therefore determined on at least two different levels, origin licensing to mark all potential origins, and origin firing to select a subset that permits assembly of the replication machinery and initiation of DNA synthesis. The extra licensed origins serve as backup and are activated only upon slowing or stalling of nearby replication forks, ensuring that DNA replication can be completed when cells encounter replication stress.[114][115] In the absence of stress, firing of extra origins is suppressed by a replication-associated signaling mechanism.[116][117] Together, the excess of licensed origins and the tight cell cycle control of origin licensing and firing embody two important strategies to prevent under- and overreplication and to maintain the integrity of eukaryotic genomes.[2]
Early studies in S. cerevisiae indicated that replication origins in eukaryotes might be recognized in a DNA-sequence-specific manner analogously to those in prokaryotes. In budding yeast, the search for genetic replicators lead to the identification of autonomously replicating sequences (ARS) that support efficient DNA replication initiation of extrachromosomal DNA.[118][119][120] These ARS regions are approximately 100-200 bp long and exhibit a multipartite organization, containing A, B1, B2, and sometimes B3 elements that together are essential for origin function.[121][122] The A element encompasses the conserved 11 bp ARS consensus sequence (ACS),[123][124] which, in conjunction with the B1 element, constitutes the primary binding site for the heterohexameric origin recognition complex (ORC), the eukaryotic replication initiator.[125][126][127][128] Within ORC, five subunits are predicated on conserved AAA+ ATPase and winged-helix folds and co-assemble into a pentameric ring that encircles DNA.[128][129][130] In budding yeast ORC, DNA binding elements in the ATPase and winged-helix domains, as well as adjacent basic patch regions in some of the ORC subunits, are positioned in the central pore of the ORC ring such that they aid the DNA-sequence-specific recognition of the ACS in an ATP-dependent manner.[128][131] By contrast, the roles of the B2 and B3 elements are less clear. The B2 region is similar to the ACS in sequence and has been suggested to function as a second ORC binding site under certain conditions, or as a binding site for the replicative helicase core.[132][133][134][135][136] Conversely, the B3 element recruits the transcription factor Abf1, albeit B3 is not found at all budding yeast origins and Abf1 binding does not appear to be strictly essential for origin function.[2][121][137][138]
Origin recognition in eukaryotes other than S. cerevisiae or its close relatives does not conform to the sequence-specific read-out of conserved origin DNA elements. Pursuits to isolate specific chromosomal replicator sequences more generally in eukaryotic species, either genetically or by genome-wide mapping of initiator binding or replication start sites, have failed to identify clear consensus sequences at origins.[139][140][141][142][143][144][145][146][147][148][149][150] Thus, sequence-specific DNA-initiator interactions in budding yeast signify a specialized mode for origin recognition in this system rather than an archetypal mode for origin specification across the eukaryotic domain. Nonetheless, DNA replication does initiate at discrete sites that are not randomly distributed across eukaryotic genomes, arguing that alternative means determine the chromosomal location of origins in these systems. These mechanisms involve a complex interplay between DNA accessibility, nucleotide sequence skew (both AT-richness and CpG islands have been linked to origins), Nucleosome positioning, epigenetic features, DNA topology and certain DNA structural features (e.g., G4 motifs), as well as regulatory proteins and transcriptional interference.[17][18][34][35][37][151][152][144][153] Importantly, origin properties vary not only between different origins in an organism and among species, but some can also change during development and cell differentiation. The chorion locus in Drosophila follicle cells constitutes a well-established example for spatial and developmental control of initiation events. This region undergoes DNA-replication-dependent gene amplification at a defined stage during oogenesis and relies on the timely and specific activation of chorion origins, which in turn is regulated by origin-specific cis-elements and several protein factors, including the Myb complex, E2F1, and E2F2.[154][155][156][157][158] This combinatorial specification and multifactorial regulation of metazoan origins has complicated the identification of unifying features that determine the location of replication start sites across eukaryotes more generally.[2]
To facilitate replication initiation and origin recognition, ORC assemblies from various species have evolved specialized auxiliary domains that are thought to aid initiator targeting to chromosomal origins or chromosomes in general. For example, the Orc4 subunit in S. pombe ORC contains several AT-hooks that preferentially bind AT-rich DNA,[159] while in metazoan ORC the TFIIB-like domain of Orc6 is thought to perform a similar function.[160] Metazoan Orc1 proteins also harbor a bromo-adjacent homology (BAH) domain that interacts with H4K20me2-nucleosomes.[109] Particularly in mammalian cells, H4K20 methylation has been reported to be required for efficient replication initiation, and the Orc1-BAH domain facilitates ORC association with chromosomes and Epstein-Barr virus origin-dependent replication.[161][162][163][164][165] Therefore, it is intriguing to speculate that both observations are mechanistically linked at least in a subset of metazoa, but this possibility needs to be further explored in future studies. In addition to the recognition of certain DNA or epigenetic features, ORC also associates directly or indirectly with several partner proteins that could aid initiator recruitment, including LRWD1, PHIP (or DCAF14), HMGA1a, among others.[33][166][167][168][169][170][171][172] Interestingly, Drosophila ORC, like its budding yeast counterpart, bends DNA and negative supercoiling has been reported to enhance DNA binding of this complex, suggesting that DNA shape and malleability might influence the location of ORC binding sites across metazoan genomes.[31][128][173][174][175] A molecular understanding for how ORC's DNA binding regions might support the read out of structural properties of the DNA duplex in metazoans rather than of specific DNA sequences as in S. cerevisiae awaits high-resolution structural information of DNA-bound metazoan initiator assemblies. Likewise, whether and how different epigenetic factors contribute to initiator recruitment in metazoan systems is poorly defined and is an important question that needs to be addressed in more detail.[2]
Once recruited to origins, ORC and its co-factors Cdc6 and Cdt1 drive the deposition of the minichromosome maintenance 2-7 (Mcm2-7) complex onto DNA.[110][176] Like the archaeal replicative helicase core, Mcm2-7 is loaded as a head-to-head double hexamer onto DNA to license origins.[111][112][113] In S-phase, Dbf4-dependent kinase (DDK) and Cyclin-dependent kinase (CDK) phosphorylate several Mcm2-7 subunits and additional initiation factors to promote the recruitment of the helicase co-activators Cdc45 and GINS, DNA melting, and ultimately bidirectional replisome assembly at a subset of the licensed origins.[28][177] In both yeast and metazoans, origins are free or depleted of nucleosomes, a property that is crucial for Mcm2-7 loading, indicating that chromatin state at origins regulates not only initiator recruitment but also helicase loading.[145][178][179][180][181][182] A permissive chromatin environment is further important for origin activation and has been implicated in regulating both origin efficiency and the timing of origin firing. Euchromatic origins typically contain active chromatin marks, replicate early, and are more efficient than late-replicating, heterochromatic origins, which conversely are characterized by repressive marks.[27][180][183] Not surprisingly, several chromatin remodelers and chromatin-modifying enzymes have been found to associate with origins and certain initiation factors,[184][185] but how their activities impact different replication initiation events remains largely obscure. Remarkably, cis-acting “early replication control elements” (ECREs) have recently also been identified to help regulate replication timing and to influence 3D genome architecture in mammalian cells.[186] Understanding the molecular and biochemical mechanisms that orchestrate this complex interplay between 3D genome organization, local and higher-order chromatin structure, and replication initiation is an exciting topic for further studies.[2]
Why have metazoan replication origins diverged from the DNA sequence-specific recognition paradigm that determines replication start sites in prokaryotes and budding yeast? Observations that metazoan origins often co-localize with promoter regions in Drosophila and mammalian cells and that replication-transcription conflicts due to collisions of the underlying molecular machineries can lead to DNA damage suggest that proper coordination of transcription and replication is important for maintaining genome stability.[140][142][144][147][187][20][21][188] Recent findings also point to a more direct role of transcription in influencing the location of origins, either by inhibiting Mcm2-7 loading or by repositioning of loaded Mcm2-7 on chromosomes.[189][153] Sequence-independent (but not necessarily random) initiator binding to DNA additionally allows for flexibility in specifying helicase loading sites and, together with transcriptional interference and the variability in activation efficiencies of licensed origins, likely determines origin location and contributes to the co-regulation of DNA replication and transcriptional programs during development and cell fate transitions. Computational modeling of initiation events in S. pombe, as well as the identification of cell-type specific and developmentally-regulated origins in metazoans, are in agreement with this notion.[141][149][190][191][192][193][194][153] However, a large degree of flexibility in origin choice also exists among different cells within a single population,[144][150][191] albeit the molecular mechanisms that lead to the heterogeneity in origin usage remain ill-defined. Mapping origins in single cells in metazoan systems and correlating these initiation events with single-cell gene expression and chromatin status will be important to elucidate whether origin choice is purely stochastic or controlled in a defined manner.[2]
Viral

Viruses often possess a single origin of replication.
A variety of proteins have been described as being involved in viral replication. For instance, Polyoma viruses utilize host cell DNA polymerases, which attach to a viral origin of replication if the T antigen is present.
Variations
Although DNA replication is essential for genetic inheritance, defined, site-specific replication origins are technically not a requirement for genome duplication as long as all chromosomes are copied in their entirety to maintain gene copy numbers. Certain bacteriophages and viruses, for example, can initiate DNA replication by homologous recombination independent of dedicated origins.[195] Likewise, the archaeon Haloferax volcanii uses recombination-dependent initiation to duplicate its genome when its endogenous origins are deleted.[81] Similar non-canonical initiation events through break-induced or transcription-initiated replication have been reported in E. coli and S. cerevisiae.[196][197][198][199][200] Nonetheless, despite the ability of cells to sustain viability under these exceptional circumstances, origin-dependent initiation is a common strategy universally adopted across different domains of life.[2]
In addition, detailed studies of replication initiation have focused on a limited number of model systems. The extensively studied fungi and metazoa are both members of the opisthokont supergroup and exemplify only a small fraction of the evolutionary landscape in the eukaryotic domain.[201] Comparably few efforts have been directed at other eukaryotic model systems, such as kinetoplastids or tetrahymena.[202][203][204][205][206][207][208] Surprisingly, these studies have revealed interesting differences both in origin properties and in initiator composition compared to yeast and metazoans.[2]
See also
- OriDB the DNA Replication Origin Database
- Origin of transfer
References
![]() This article was adapted from the following source under a CC BY 4.0, 🅭🅯 4.0 license (2019) (reviewer reports):
Babatunde Ekundayo; Franziska Bleichert (12 September 2019). "Origins of DNA replication". PLOS Genetics. 15 (9): e1008320. doi:10.1371/JOURNAL.PGEN.1008320. ISSN 1553-7390. PMC 6742236. PMID 31513569. Wikidata Q86320168.
This article was adapted from the following source under a CC BY 4.0, 🅭🅯 4.0 license (2019) (reviewer reports):
Babatunde Ekundayo; Franziska Bleichert (12 September 2019). "Origins of DNA replication". PLOS Genetics. 15 (9): e1008320. doi:10.1371/JOURNAL.PGEN.1008320. ISSN 1553-7390. PMC 6742236. PMID 31513569. Wikidata Q86320168.
- Wagner EK, Hewlett M, Bloom D, Camerini D, eds. (2008). "Technical Glossary" (PDF). Basic Virology (3rd ed.). Malden, MA: Blackwell Publishing. ISBN 978-1-4051-4715-6.
- Ekundayo B, Bleichert F (September 2019). "Origins of DNA replication". PLOS Genetics. 15 (9): e1008320. doi:10.1371/journal.pgen.1008320. PMC 6742236. PMID 31513569.
 Material was copied from this source, which is available under a Creative Commons Attribution 4.0 International License.
Material was copied from this source, which is available under a Creative Commons Attribution 4.0 International License. - Hulo C, de Castro E, Masson P, Bougueleret L, Bairoch A, Xenarios I, Le Mercier P (January 2011). "ViralZone: a knowledge resource to understand virus diversity". Nucleic Acids Research. 39 (Database issue): D576-82. doi:10.1093/nar/gkq901. PMC 3013774. PMID 20947564.
- Mendel JG (1866). "Versuche über Pflanzenhybriden". Verhandlungen des naturforschenden Vereines in Brünn. Im Verlage des Vereines. pp. 3–47. For the English translation, see: Druery CT, Bateson W (1901). "Experiments in plant hybridization" (PDF). Journal of the Royal Horticultural Society. 26: 1–32. Retrieved 9 October 2009.
- Avery OT, Macleod CM, McCarty M (February 1944). "Studies on the Chemical Nature of the Substance Inducing Transformation of Pneumococcal Types : Induction of Transformation by a Desoxyribonucleic Acid Fraction Isolated From Pneumococcus Type III". The Journal of Experimental Medicine. 79 (2): 137–58. doi:10.1084/jem.79.2.137. PMC 2135445. PMID 19871359.
- Watson JD, Crick FH (1953). "The structure of DNA". Cold Spring Harbor Symposia on Quantitative Biology. 18: 123–31. doi:10.1101/sqb.1953.018.01.020. PMID 13168976.
- Meselson M, Stahl FW (July 1958). "The replication of DNA in Escherichia coli". Proceedings of the National Academy of Sciences of the United States of America. 44 (7): 671–82. Bibcode:1958PNAS...44..671M. doi:10.1073/pnas.44.7.671. PMC 528642. PMID 16590258.
- Meselson M, Stahl FW (1958). "The replication of DNA". Cold Spring Harbor Symposia on Quantitative Biology. 23: 9–12. doi:10.1101/sqb.1958.023.01.004. PMID 13635537.
- Lehman IR, Bessman MJ, Simms ES, Kornberg A (July 1958). "Enzymatic synthesis of deoxyribonucleic acid. I. Preparation of substrates and partial purification of an enzyme from Escherichia coli". The Journal of Biological Chemistry. 233 (1): 163–70. doi:10.1016/S0021-9258(19)68048-8. PMID 13563462.
- O'Donnell M, Langston L, Stillman B (July 2013). "Principles and concepts of DNA replication in bacteria, archaea, and eukarya". Cold Spring Harbor Perspectives in Biology. 5 (7): a010108. doi:10.1101/cshperspect.a010108. PMC 3685895. PMID 23818497.
- Abbas T, Keaton MA, Dutta A (March 2013). "Genomic instability in cancer". Cold Spring Harbor Perspectives in Biology. 5 (3): a012914. doi:10.1101/cshperspect.a012914. PMC 3578360. PMID 23335075.
- Barlow JH, Nussenzweig A (December 2014). "Replication initiation and genome instability: a crossroads for DNA and RNA synthesis". Cellular and Molecular Life Sciences. 71 (23): 4545–59. doi:10.1007/s00018-014-1721-1. PMC 6289259. PMID 25238783.
- Siddiqui K, On KF, Diffley JF (September 2013). "Regulating DNA replication in eukarya". Cold Spring Harbor Perspectives in Biology. 5 (9): a012930. doi:10.1101/cshperspect.a012930. PMC 3753713. PMID 23838438.
- Sclafani RA, Holzen TM (2007). "Cell cycle regulation of DNA replication". Annual Review of Genetics. 41: 237–80. doi:10.1146/annurev.genet.41.110306.130308. PMC 2292467. PMID 17630848.
- García-Muse T, Aguilera A (September 2016). "Transcription-replication conflicts: how they occur and how they are resolved". Nature Reviews. Molecular Cell Biology. 17 (9): 553–63. doi:10.1038/nrm.2016.88. hdl:11441/101680. PMID 27435505. S2CID 7617164.
- Yakovchuk P, Protozanova E, Frank-Kamenetskii MD (2006). "Base-stacking and base-pairing contributions into thermal stability of the DNA double helix". Nucleic Acids Research. 34 (2): 564–74. doi:10.1093/nar/gkj454. PMC 1360284. PMID 16449200.
- Leonard AC, Méchali M (October 2013). "DNA replication origins". Cold Spring Harbor Perspectives in Biology. 5 (10): a010116. doi:10.1101/cshperspect.a010116. PMC 3783049. PMID 23838439.
- Creager RL, Li Y, MacAlpine DM (April 2015). "SnapShot: Origins of DNA replication". Cell. 161 (2): 418–418.e1. doi:10.1016/j.cell.2015.03.043. PMID 25860614.
- Knott SR, Viggiani CJ, Aparicio OM (August 2009). "To promote and protect: coordinating DNA replication and transcription for genome stability". Epigenetics. 4 (6): 362–5. doi:10.4161/epi.4.6.9712. PMID 19736523.
- Deshpande AM, Newlon CS (May 1996). "DNA replication fork pause sites dependent on transcription". Science. 272 (5264): 1030–3. Bibcode:1996Sci...272.1030D. doi:10.1126/science.272.5264.1030. PMID 8638128. S2CID 38817771.
- Sankar TS, Wastuwidyaningtyas BD, Dong Y, Lewis SA, Wang JD (July 2016). "The nature of mutations induced by replication–transcription collisions". Nature. 535 (7610): 178–81. Bibcode:2016Natur.535..178S. doi:10.1038/nature18316. PMC 4945378. PMID 27362223.
- Liu B, Alberts BM (February 1995). "Head-on collision between a DNA replication apparatus and RNA polymerase transcription complex". Science. 267 (5201): 1131–7. Bibcode:1995Sci...267.1131L. doi:10.1126/science.7855590. PMID 7855590. S2CID 6835136.
- Azvolinsky A, Giresi PG, Lieb JD, Zakian VA (June 2009). "Highly transcribed RNA polymerase II genes are impediments to replication fork progression in Saccharomyces cerevisiae". Molecular Cell. 34 (6): 722–34. doi:10.1016/j.molcel.2009.05.022. PMC 2728070. PMID 19560424.
- Jacob F, Brenner S, Cuzin F (1963-01-01). "On the Regulation of Dna Replication in Bacteria". Cold Spring Harbor Symposia on Quantitative Biology. 28: 329–348. doi:10.1101/sqb.1963.028.01.048. ISSN 0091-7451.
- Novick RP (December 1987). "Plasmid incompatibility". Microbiological Reviews. 51 (4): 381–95. doi:10.1128/MMBR.51.4.381-395.1987. PMC 373122. PMID 3325793.
- Skarstad K, Katayama T (April 2013). "Regulating DNA replication in bacteria". Cold Spring Harbor Perspectives in Biology. 5 (4): a012922. doi:10.1101/cshperspect.a012922. PMC 3683904. PMID 23471435.
- Marks AB, Fu H, Aladjem MI (2017). "Regulation of Replication Origins". Advances in Experimental Medicine and Biology. 1042: 43–59. doi:10.1007/978-981-10-6955-0_2. ISBN 978-981-10-6954-3. PMC 6622447. PMID 29357052.
- Parker MW, Botchan MR, Berger JM (April 2017). "Mechanisms and regulation of DNA replication initiation in eukaryotes". Critical Reviews in Biochemistry and Molecular Biology. 52 (2): 107–144. doi:10.1080/10409238.2016.1274717. PMC 5545932. PMID 28094588.
- Gilbert DM (October 2004). "In search of the holy replicator". Nature Reviews. Molecular Cell Biology. 5 (10): 848–55. doi:10.1038/nrm1495. PMC 1255919. PMID 15459665.
- Aladjem MI, Fanning E (July 2004). "The replicon revisited: an old model learns new tricks in metazoan chromosomes". EMBO Reports. 5 (7): 686–91. doi:10.1038/sj.embor.7400185. PMC 1299096. PMID 15229645.
- Remus D, Beall EL, Botchan MR (February 2004). "DNA topology, not DNA sequence, is a critical determinant for Drosophila ORC-DNA binding". The EMBO Journal. 23 (4): 897–907. doi:10.1038/sj.emboj.7600077. PMC 380993. PMID 14765124.
- Vashee S, Cvetic C, Lu W, Simancek P, Kelly TJ, Walter JC (August 2003). "Sequence-independent DNA binding and replication initiation by the human origin recognition complex". Genes & Development. 17 (15): 1894–908. doi:10.1101/gad.1084203. PMC 196240. PMID 12897055.
- Shen Z, Sathyan KM, Geng Y, Zheng R, Chakraborty A, Freeman B, et al. (October 2010). "A WD-repeat protein stabilizes ORC binding to chromatin". Molecular Cell. 40 (1): 99–111. doi:10.1016/j.molcel.2010.09.021. PMC 5201136. PMID 20932478.
- Dorn ES, Cook JG (May 2011). "Nucleosomes in the neighborhood: new roles for chromatin modifications in replication origin control". Epigenetics. 6 (5): 552–9. doi:10.4161/epi.6.5.15082. PMC 3230546. PMID 21364325.
- Aladjem MI, Redon CE (February 2017). "Order from clutter: selective interactions at mammalian replication origins". Nature Reviews. Genetics. 18 (2): 101–116. doi:10.1038/nrg.2016.141. PMC 6596300. PMID 27867195.
- Fragkos M, Ganier O, Coulombe P, Méchali M (June 2015). "DNA replication origin activation in space and time". Nature Reviews. Molecular Cell Biology. 16 (6): 360–74. doi:10.1038/nrm4002. PMID 25999062. S2CID 37108355.
- Prioleau MN, MacAlpine DM (August 2016). "DNA replication origins-where do we begin?". Genes & Development. 30 (15): 1683–97. doi:10.1101/gad.285114.116. PMC 5002974. PMID 27542827.
- Cayrou C, Coulombe P, Puy A, Rialle S, Kaplan N, Segal E, Méchali M (February 2012). "New insights into replication origin characteristics in metazoans". Cell Cycle. 11 (4): 658–67. doi:10.4161/cc.11.4.19097. PMC 3318102. PMID 22373526.
- Lombraña R, Almeida R, Álvarez A, Gómez M (2015). "R-loops and initiation of DNA replication in human cells: a missing link?". Frontiers in Genetics. 6: 158. doi:10.3389/fgene.2015.00158. PMC 4412123. PMID 25972891.
- Jang SM, Zhang Y, Utani K, Fu H, Redon CE, Marks AB, et al. (July 2018). "The replication initiation determinant protein (RepID) modulates replication by recruiting CUL4 to chromatin". Nature Communications. 9 (1): 2782. Bibcode:2018NatCo...9.2782J. doi:10.1038/s41467-018-05177-6. PMC 6050238. PMID 30018425.
- Zakian VA, Scott JF (March 1982). "Construction, replication, and chromatin structure of TRP1 RI circle, a multiple-copy synthetic plasmid derived from Saccharomyces cerevisiae chromosomal DNA". Molecular and Cellular Biology. 2 (3): 221–32. doi:10.1128/mcb.2.3.221. PMC 369780. PMID 6287231.
- Rhodes N, Company M, Errede B (March 1990). "A yeast-Escherichia coli shuttle vector containing the M13 origin of replication". Plasmid. 23 (2): 159–62. doi:10.1016/0147-619x(90)90036-c. PMID 2194231.
- Paululat A, Heinisch JJ (December 2012). "New yeast/E. coli/Drosophila triple shuttle vectors for efficient generation of Drosophila P element transformation constructs". Gene. 511 (2): 300–5. doi:10.1016/j.gene.2012.09.058. PMID 23026211.
- Ryan VT, Grimwade JE, Camara JE, Crooke E, Leonard AC (March 2004). "Escherichia coli prereplication complex assembly is regulated by dynamic interplay among Fis, IHF and DnaA". Molecular Microbiology. 51 (5): 1347–59. doi:10.1046/j.1365-2958.2003.03906.x. PMID 14982629. S2CID 22598422.
- Mackiewicz P, Zakrzewska-Czerwinska J, Zawilak A, Dudek MR, Cebrat S (2004). "Where does bacterial replication start? Rules for predicting the oriC region". Nucleic Acids Research. 32 (13): 3781–91. doi:10.1093/nar/gkh699. PMC 506792. PMID 15258248.
- Luo H, Gao F (January 2019). "DoriC 10.0: an updated database of replication origins in prokaryotic genomes including chromosomes and plasmids". Nucleic Acids Research. 47 (D1): D74–D77. doi:10.1093/nar/gky1014. PMC 6323995. PMID 30364951.
- Fuller RS, Funnell BE, Kornberg A (October 1984). "The dnaA protein complex with the E. coli chromosomal replication origin (oriC) and other DNA sites". Cell. 38 (3): 889–900. doi:10.1016/0092-8674(84)90284-8. PMID 6091903. S2CID 23316215.
- Fuller RS, Kornberg A (October 1983). "Purified dnaA protein in initiation of replication at the Escherichia coli chromosomal origin of replication". Proceedings of the National Academy of Sciences of the United States of America. 80 (19): 5817–21. Bibcode:1983PNAS...80.5817F. doi:10.1073/pnas.80.19.5817. PMC 390166. PMID 6310593.
- Jakimowicz D, Majka J, Messer W, Speck C, Fernandez M, Martin MC, et al. (May 1998). "Structural elements of the Streptomyces oriC region and their interactions with the DnaA protein". Microbiology. 144 ( Pt 5) (5): 1281–90. doi:10.1099/00221287-144-5-1281. PMID 9611803.
- Tsodikov OV, Biswas T (July 2011). "Structural and thermodynamic signatures of DNA recognition by Mycobacterium tuberculosis DnaA". Journal of Molecular Biology. 410 (3): 461–76. doi:10.1016/j.jmb.2011.05.007. PMID 21620858.
- Costa A, Hood IV, Berger JM (2013). "Mechanisms for initiating cellular DNA replication". Annual Review of Biochemistry. 82: 25–54. doi:10.1146/annurev-biochem-052610-094414. PMC 4696014. PMID 23746253.
- Wolański M, Donczew R, Zawilak-Pawlik A, Zakrzewska-Czerwińska J (2014). "oriC-encoded instructions for the initiation of bacterial chromosome replication". Frontiers in Microbiology. 5: 735. doi:10.3389/fmicb.2014.00735. PMC 4285127. PMID 25610430.
- Messer W, Blaesing F, Majka J, Nardmann J, Schaper S, Schmidt A, et al. (1999). "Functional domains of DnaA proteins". Biochimie. 81 (8–9): 819–25. doi:10.1016/s0300-9084(99)00215-1. PMID 10572294.
- Sutton MD, Kaguni JM (December 1997). "The Escherichia coli dnaA gene: four functional domains". Journal of Molecular Biology. 274 (4): 546–61. doi:10.1006/jmbi.1997.1425. PMID 9417934.
- Speck C, Messer W (March 2001). "Mechanism of origin unwinding: sequential binding of DnaA to double- and single-stranded DNA". The EMBO Journal. 20 (6): 1469–76. doi:10.1093/emboj/20.6.1469. PMC 145534. PMID 11250912.
- Fujikawa N, Kurumizaka H, Nureki O, Terada T, Shirouzu M, Katayama T, Yokoyama S (April 2003). "Structural basis of replication origin recognition by the DnaA protein". Nucleic Acids Research. 31 (8): 2077–86. doi:10.1093/nar/gkg309. PMC 153737. PMID 12682358.
- Duderstadt KE, Chuang K, Berger JM (October 2011). "DNA stretching by bacterial initiators promotes replication origin opening". Nature. 478 (7368): 209–13. Bibcode:2011Natur.478..209D. doi:10.1038/nature10455. PMC 3192921. PMID 21964332.
- Erzberger JP, Pirruccello MM, Berger JM (September 2002). "The structure of bacterial DnaA: implications for general mechanisms underlying DNA replication initiation". The EMBO Journal. 21 (18): 4763–73. doi:10.1093/emboj/cdf496. PMC 126292. PMID 12234917.
- Sutton MD, Kaguni JM (September 1997). "Threonine 435 of Escherichia coli DnaA protein confers sequence-specific DNA binding activity". The Journal of Biological Chemistry. 272 (37): 23017–24. doi:10.1074/jbc.272.37.23017. PMID 9287298.
- Bramhill D, Kornberg A (September 1988). "A model for initiation at origins of DNA replication". Cell. 54 (7): 915–8. doi:10.1016/0092-8674(88)90102-x. PMID 2843291. S2CID 1705480.
- Rozgaja TA, Grimwade JE, Iqbal M, Czerwonka C, Vora M, Leonard AC (October 2011). "Two oppositely oriented arrays of low-affinity recognition sites in oriC guide progressive binding of DnaA during Escherichia coli pre-RC assembly". Molecular Microbiology. 82 (2): 475–88. doi:10.1111/j.1365-2958.2011.07827.x. PMC 3192301. PMID 21895796.
- Zawilak-Pawlik A, Kois A, Majka J, Jakimowicz D, Smulczyk-Krawczyszyn A, Messer W, Zakrzewska-Czerwińska J (July 2005). "Architecture of bacterial replication initiation complexes: orisomes from four unrelated bacteria". The Biochemical Journal. 389 (Pt 2): 471–81. doi:10.1042/BJ20050143. PMC 1175125. PMID 15790315.
- Grimwade JE, Rozgaja TA, Gupta R, Dyson K, Rao P, Leonard AC (July 2018). "Origin recognition is the predominant role for DnaA-ATP in initiation of chromosome replication". Nucleic Acids Research. 46 (12): 6140–6151. doi:10.1093/nar/gky457. PMC 6158602. PMID 29800247.
- Sakiyama Y, Kasho K, Noguchi Y, Kawakami H, Katayama T (December 2017). "Regulatory dynamics in the ternary DnaA complex for initiation of chromosomal replication in Escherichia coli". Nucleic Acids Research. 45 (21): 12354–12373. doi:10.1093/nar/gkx914. PMC 5716108. PMID 29040689.
- Matsui M, Oka A, Takanami M, Yasuda S, Hirota Y (August 1985). "Sites of dnaA protein-binding in the replication origin of the Escherichia coli K-12 chromosome". Journal of Molecular Biology. 184 (3): 529–33. doi:10.1016/0022-2836(85)90299-2. PMID 2995681.
- Margulies C, Kaguni JM (July 1996). "Ordered and sequential binding of DnaA protein to oriC, the chromosomal origin of Escherichia coli". The Journal of Biological Chemistry. 271 (29): 17035–40. doi:10.1074/jbc.271.29.17035. PMID 8663334.
- Schaper S, Messer W (July 1995). "Interaction of the initiator protein DnaA of Escherichia coli with its DNA target". The Journal of Biological Chemistry. 270 (29): 17622–6. doi:10.1074/jbc.270.29.17622. PMID 7615570.
- Weigel C, Schmidt A, Rückert B, Lurz R, Messer W (November 1997). "DnaA protein binding to individual DnaA boxes in the Escherichia coli replication origin, oriC". The EMBO Journal. 16 (21): 6574–83. doi:10.1093/emboj/16.21.6574. PMC 1170261. PMID 9351837.
- Samitt CE, Hansen FG, Miller JF, Schaechter M (March 1989). "In vivo studies of DnaA binding to the origin of replication of Escherichia coli". The EMBO Journal. 8 (3): 989–93. doi:10.1002/j.1460-2075.1989.tb03462.x. PMC 400901. PMID 2542031.
- McGarry KC, Ryan VT, Grimwade JE, Leonard AC (March 2004). "Two discriminatory binding sites in the Escherichia coli replication origin are required for DNA strand opening by initiator DnaA-ATP". Proceedings of the National Academy of Sciences of the United States of America. 101 (9): 2811–6. Bibcode:2004PNAS..101.2811M. doi:10.1073/pnas.0400340101. PMC 365702. PMID 14978287.
- Kawakami H, Keyamura K, Katayama T (July 2005). "Formation of an ATP-DnaA-specific initiation complex requires DnaA Arginine 285, a conserved motif in the AAA+ protein family". The Journal of Biological Chemistry. 280 (29): 27420–30. doi:10.1074/jbc.M502764200. PMID 15901724.
- Speck C, Weigel C, Messer W (November 1999). "ATP- and ADP-dnaA protein, a molecular switch in gene regulation". The EMBO Journal. 18 (21): 6169–76. doi:10.1093/emboj/18.21.6169. PMC 1171680. PMID 10545126.
- Miller DT, Grimwade JE, Betteridge T, Rozgaja T, Torgue JJ, Leonard AC (November 2009). "Bacterial origin recognition complexes direct assembly of higher-order DnaA oligomeric structures". Proceedings of the National Academy of Sciences of the United States of America. 106 (44): 18479–84. Bibcode:2009PNAS..10618479M. doi:10.1073/pnas.0909472106. PMC 2773971. PMID 19833870.
- Erzberger JP, Mott ML, Berger JM (August 2006). "Structural basis for ATP-dependent DnaA assembly and replication-origin remodeling". Nature Structural & Molecular Biology. 13 (8): 676–83. doi:10.1038/nsmb1115. PMID 16829961. S2CID 23586302.
- Zorman S, Seitz H, Sclavi B, Strick TR (August 2012). "Topological characterization of the DnaA-oriC complex using single-molecule nanomanipuation". Nucleic Acids Research. 40 (15): 7375–83. doi:10.1093/nar/gks371. PMC 3424547. PMID 22581769.
- Richardson TT, Harran O, Murray H (June 2016). "The bacterial DnaA-trio replication origin element specifies single-stranded DNA initiator binding". Nature. 534 (7607): 412–6. Bibcode:2016Natur.534..412R. doi:10.1038/nature17962. PMC 4913881. PMID 27281207.
- Duderstadt KE, Mott ML, Crisona NJ, Chuang K, Yang H, Berger JM (September 2010). "Origin remodeling and opening in bacteria rely on distinct assembly states of the DnaA initiator". The Journal of Biological Chemistry. 285 (36): 28229–39. doi:10.1074/jbc.M110.147975. PMC 2934688. PMID 20595381.
- Ozaki S, Katayama T (February 2012). "Highly organized DnaA-oriC complexes recruit the single-stranded DNA for replication initiation". Nucleic Acids Research. 40 (4): 1648–65. doi:10.1093/nar/gkr832. PMC 3287180. PMID 22053082.
- Myllykallio H, Lopez P, López-García P, Heilig R, Saurin W, Zivanovic Y, et al. (June 2000). "Bacterial mode of replication with eukaryotic-like machinery in a hyperthermophilic archaeon". Science. 288 (5474): 2212–5. Bibcode:2000Sci...288.2212M. doi:10.1126/science.288.5474.2212. PMID 10864870.
- Norais C, Hawkins M, Hartman AL, Eisen JA, Myllykallio H, Allers T (May 2007). "Genetic and physical mapping of DNA replication origins in Haloferax volcanii". PLOS Genetics. 3 (5): e77. doi:10.1371/journal.pgen.0030077. PMC 1868953. PMID 17511521.
- Hawkins M, Malla S, Blythe MJ, Nieduszynski CA, Allers T (November 2013). "Accelerated growth in the absence of DNA replication origins". Nature. 503 (7477): 544–547. Bibcode:2013Natur.503..544H. doi:10.1038/nature12650. PMC 3843117. PMID 24185008.
- Wu Z, Liu J, Yang H, Liu H, Xiang H (February 2014). "Multiple replication origins with diverse control mechanisms in Haloarcula hispanica". Nucleic Acids Research. 42 (4): 2282–94. doi:10.1093/nar/gkt1214. PMC 3936714. PMID 24271389.
- Pelve EA, Martens-Habbena W, Stahl DA, Bernander R (November 2013). "Mapping of active replication origins in vivo in thaum- and euryarchaeal replicons". Molecular Microbiology. 90 (3): 538–50. doi:10.1111/mmi.12382. PMID 23991938.
- Pelve EA, Lindås AC, Knöppel A, Mira A, Bernander R (September 2012). "Four chromosome replication origins in the archaeon Pyrobaculum calidifontis". Molecular Microbiology. 85 (5): 986–95. doi:10.1111/j.1365-2958.2012.08155.x. PMID 22812406.
- Robinson NP, Dionne I, Lundgren M, Marsh VL, Bernander R, Bell SD (January 2004). "Identification of two origins of replication in the single chromosome of the archaeon Sulfolobus solfataricus". Cell. 116 (1): 25–38. doi:10.1016/s0092-8674(03)01034-1. PMID 14718164. S2CID 12777774.
- Lundgren M, Andersson A, Chen L, Nilsson P, Bernander R (May 2004). "Three replication origins in Sulfolobus species: synchronous initiation of chromosome replication and asynchronous termination". Proceedings of the National Academy of Sciences of the United States of America. 101 (18): 7046–51. Bibcode:2004PNAS..101.7046L. doi:10.1073/pnas.0400656101. PMC 406463. PMID 15107501.
- Bell SD (2017). "Initiation of DNA Replication in the Archaea". Advances in Experimental Medicine and Biology. 1042: 99–115. doi:10.1007/978-981-10-6955-0_5. ISBN 978-981-10-6954-3. PMID 29357055.
- Ausiannikava D, Allers T (January 2017). "Diversity of DNA Replication in the Archaea". Genes. 8 (2): 56. doi:10.3390/genes8020056. PMC 5333045. PMID 28146124.
- Wu Z, Liu J, Yang H, Xiang H (2014). "DNA replication origins in archaea". Frontiers in Microbiology. 5: 179. doi:10.3389/fmicb.2014.00179. PMC 4010727. PMID 24808892.
- Matsunaga F, Forterre P, Ishino Y, Myllykallio H (September 2001). "In vivo interactions of archaeal Cdc6/Orc1 and minichromosome maintenance proteins with the replication origin". Proceedings of the National Academy of Sciences of the United States of America. 98 (20): 11152–7. Bibcode:2001PNAS...9811152M. doi:10.1073/pnas.191387498. PMC 58699. PMID 11562464.
- Wu Z, Liu H, Liu J, Liu X, Xiang H (September 2012). "Diversity and evolution of multiple orc/cdc6-adjacent replication origins in haloarchaea". BMC Genomics. 13: 478. doi:10.1186/1471-2164-13-478. PMC 3528665. PMID 22978470.
- Bell SD (2012). "Archaeal orc1/cdc6 proteins". The Eukaryotic Replisome: A Guide to Protein Structure and Function. Subcellular Biochemistry. Vol. 62. pp. 59–69. doi:10.1007/978-94-007-4572-8_4. ISBN 978-94-007-4571-1. PMID 22918580.
- Samson RY, Xu Y, Gadelha C, Stone TA, Faqiri JN, Li D, et al. (February 2013). "Specificity and function of archaeal DNA replication initiator proteins". Cell Reports. 3 (2): 485–96. doi:10.1016/j.celrep.2013.01.002. PMC 3607249. PMID 23375370.
- Grainge I, Gaudier M, Schuwirth BS, Westcott SL, Sandall J, Atanassova N, Wigley DB (October 2006). "Biochemical analysis of a DNA replication origin in the archaeon Aeropyrum pernix". Journal of Molecular Biology. 363 (2): 355–69. doi:10.1016/j.jmb.2006.07.076. PMID 16978641.
- Robinson NP, Bell SD (April 2007). "Extrachromosomal element capture and the evolution of multiple replication origins in archaeal chromosomes". Proceedings of the National Academy of Sciences of the United States of America. 104 (14): 5806–11. Bibcode:2007PNAS..104.5806R. doi:10.1073/pnas.0700206104. PMC 1851573. PMID 17392430.
- Robinson NP, Blood KA, McCallum SA, Edwards PA, Bell SD (February 2007). "Sister chromatid junctions in the hyperthermophilic archaeon Sulfolobus solfataricus". The EMBO Journal. 26 (3): 816–24. doi:10.1038/sj.emboj.7601529. PMC 1794387. PMID 17255945.
- Dueber EL, Corn JE, Bell SD, Berger JM (August 2007). "Replication origin recognition and deformation by a heterodimeric archaeal Orc1 complex". Science. 317 (5842): 1210–3. Bibcode:2007Sci...317.1210D. doi:10.1126/science.1143690. PMID 17761879. S2CID 45665434.
- Gaudier M, Schuwirth BS, Westcott SL, Wigley DB (August 2007). "Structural basis of DNA replication origin recognition by an ORC protein". Science. 317 (5842): 1213–6. Bibcode:2007Sci...317.1213G. doi:10.1126/science.1143664. PMID 17761880.
- Capaldi SA, Berger JM (2004). "Biochemical characterization of Cdc6/Orc1 binding to the replication origin of the euryarchaeon Methanothermobacter thermoautotrophicus". Nucleic Acids Research. 32 (16): 4821–32. doi:10.1093/nar/gkh819. PMC 519113. PMID 15358831.
- Liu J, Smith CL, DeRyckere D, DeAngelis K, Martin GS, Berger JM (September 2000). "Structure and function of Cdc6/Cdc18: implications for origin recognition and checkpoint control". Molecular Cell. 6 (3): 637–48. doi:10.1016/s1097-2765(00)00062-9. PMID 11030343.
- Singleton MR, Morales R, Grainge I, Cook N, Isupov MN, Wigley DB (October 2004). "Conformational changes induced by nucleotide binding in Cdc6/ORC from Aeropyrum pernix". Journal of Molecular Biology. 343 (3): 547–57. doi:10.1016/j.jmb.2004.08.044. PMID 15465044.
- Matsunaga F, Norais C, Forterre P, Myllykallio H (February 2003). "Identification of short 'eukaryotic' Okazaki fragments synthesized from a prokaryotic replication origin". EMBO Reports. 4 (2): 154–8. doi:10.1038/sj.embor.embor732. PMC 1315830. PMID 12612604.
- Berquist BR, DasSarma S (October 2003). "An archaeal chromosomal autonomously replicating sequence element from an extreme halophile, Halobacterium sp. strain NRC-1". Journal of Bacteriology. 185 (20): 5959–66. doi:10.1128/jb.185.20.5959-5966.2003. PMC 225043. PMID 14526006.
- Kasiviswanathan R, Shin JH, Kelman Z (2005). "Interactions between the archaeal Cdc6 and MCM proteins modulate their biochemical properties". Nucleic Acids Research. 33 (15): 4940–50. doi:10.1093/nar/gki807. PMC 1201339. PMID 16150924.
- Samson RY, Abeyrathne PD, Bell SD (January 2016). "Mechanism of Archaeal MCM Helicase Recruitment to DNA Replication Origins". Molecular Cell. 61 (2): 287–96. doi:10.1016/j.molcel.2015.12.005. PMC 4724246. PMID 26725007.
- Dueber EC, Costa A, Corn JE, Bell SD, Berger JM (May 2011). "Molecular determinants of origin discrimination by Orc1 initiators in archaea". Nucleic Acids Research. 39 (9): 3621–31. doi:10.1093/nar/gkq1308. PMC 3089459. PMID 21227921.
- Matsunaga F, Takemura K, Akita M, Adachi A, Yamagami T, Ishino Y (January 2010). "Localized melting of duplex DNA by Cdc6/Orc1 at the DNA replication origin in the hyperthermophilic archaeon Pyrococcus furiosus". Extremophiles. 14 (1): 21–31. doi:10.1007/s00792-009-0284-9. PMID 19787415. S2CID 21336802.
- Onishi M, Liou GG, Buchberger JR, Walz T, Moazed D (December 2007). "Role of the conserved Sir3-BAH domain in nucleosome binding and silent chromatin assembly". Molecular Cell. 28 (6): 1015–28. doi:10.1016/j.molcel.2007.12.004. PMID 18158899.
- Kuo AJ, Song J, Cheung P, Ishibe-Murakami S, Yamazoe S, Chen JK, et al. (March 2012). "The BAH domain of ORC1 links H4K20me2 to DNA replication licensing and Meier-Gorlin syndrome". Nature. 484 (7392): 115–9. Bibcode:2012Natur.484..115K. doi:10.1038/nature10956. PMC 3321094. PMID 22398447.
- Bleichert F, Botchan MR, Berger JM (February 2017). "Mechanisms for initiating cellular DNA replication". Science. 355 (6327): eaah6317. doi:10.1126/science.aah6317. PMID 28209641.
- Gambus A, Khoudoli GA, Jones RC, Blow JJ (April 2011). "MCM2-7 form double hexamers at licensed origins in Xenopus egg extract". The Journal of Biological Chemistry. 286 (13): 11855–64. doi:10.1074/jbc.M110.199521. PMC 3064236. PMID 21282109.
- Remus D, Beuron F, Tolun G, Griffith JD, Morris EP, Diffley JF (November 2009). "Concerted loading of Mcm2-7 double hexamers around DNA during DNA replication origin licensing". Cell. 139 (4): 719–30. doi:10.1016/j.cell.2009.10.015. PMC 2804858. PMID 19896182.
- Evrin C, Clarke P, Zech J, Lurz R, Sun J, Uhle S, et al. (December 2009). "A double-hexameric MCM2-7 complex is loaded onto origin DNA during licensing of eukaryotic DNA replication". Proceedings of the National Academy of Sciences of the United States of America. 106 (48): 20240–5. Bibcode:2009PNAS..10620240E. doi:10.1073/pnas.0911500106. PMC 2787165. PMID 19910535.
- Ge XQ, Jackson DA, Blow JJ (December 2007). "Dormant origins licensed by excess Mcm2-7 are required for human cells to survive replicative stress". Genes & Development. 21 (24): 3331–41. doi:10.1101/gad.457807. PMC 2113033. PMID 18079179.
- Ibarra A, Schwob E, Méndez J (July 2008). "Excess MCM proteins protect human cells from replicative stress by licensing backup origins of replication". Proceedings of the National Academy of Sciences of the United States of America. 105 (26): 8956–61. Bibcode:2008PNAS..105.8956I. doi:10.1073/pnas.0803978105. PMC 2449346. PMID 18579778.
- Moiseeva TN, Yin Y, Calderon MJ, Qian C, Schamus-Haynes S, Sugitani N, et al. (July 2019). "An ATR and CHK1 kinase signaling mechanism that limits origin firing during unperturbed DNA replication". Proceedings of the National Academy of Sciences of the United States of America. 116 (27): 13374–13383. doi:10.1073/pnas.1903418116. PMC 6613105. PMID 31209037.
- Moiseeva TN, Bakkenist CJ (September 2019). "Dormant origin signaling during unperturbed replication". DNA Repair. 81: 102655. doi:10.1016/j.dnarep.2019.102655. PMC 6764875. PMID 31311769.
- Stinchcomb DT, Struhl K, Davis RW (November 1979). "Isolation and characterisation of a yeast chromosomal replicator". Nature. 282 (5734): 39–43. Bibcode:1979Natur.282...39S. doi:10.1038/282039a0. PMID 388229. S2CID 4326901.
- Huberman JA, Spotila LD, Nawotka KA, el-Assouli SM, Davis LR (November 1987). "The in vivo replication origin of the yeast 2 microns plasmid". Cell. 51 (3): 473–81. doi:10.1016/0092-8674(87)90643-x. PMID 3311385. S2CID 54385402.
- Brewer BJ, Fangman WL (November 1987). "The localization of replication origins on ARS plasmids in S. cerevisiae". Cell. 51 (3): 463–71. doi:10.1016/0092-8674(87)90642-8. PMID 2822257. S2CID 20152681.
- Marahrens Y, Stillman B (February 1992). "A yeast chromosomal origin of DNA replication defined by multiple functional elements". Science. 255 (5046): 817–23. Bibcode:1992Sci...255..817M. doi:10.1126/science.1536007. PMID 1536007.
- Rao H, Marahrens Y, Stillman B (November 1994). "Functional conservation of multiple elements in yeast chromosomal replicators". Molecular and Cellular Biology. 14 (11): 7643–51. doi:10.1128/mcb.14.11.7643. PMC 359300. PMID 7935478.
- Broach JR, Li YY, Feldman J, Jayaram M, Abraham J, Nasmyth KA, Hicks JB (1983). "Localization and sequence analysis of yeast origins of DNA replication". Cold Spring Harbor Symposia on Quantitative Biology. 47 Pt 2: 1165–73. doi:10.1101/sqb.1983.047.01.132. PMID 6345070.
- Celniker SE, Sweder K, Srienc F, Bailey JE, Campbell JL (November 1984). "Deletion mutations affecting autonomously replicating sequence ARS1 of Saccharomyces cerevisiae". Molecular and Cellular Biology. 4 (11): 2455–66. doi:10.1128/mcb.4.11.2455. PMC 369077. PMID 6392851.
- Rao H, Stillman B (March 1995). "The origin recognition complex interacts with a bipartite DNA binding site within yeast replicators". Proceedings of the National Academy of Sciences of the United States of America. 92 (6): 2224–8. Bibcode:1995PNAS...92.2224R. doi:10.1073/pnas.92.6.2224. PMC 42456. PMID 7892251.
- Rowley A, Cocker JH, Harwood J, Diffley JF (June 1995). "Initiation complex assembly at budding yeast replication origins begins with the recognition of a bipartite sequence by limiting amounts of the initiator, ORC". The EMBO Journal. 14 (11): 2631–41. doi:10.1002/j.1460-2075.1995.tb07261.x. PMC 398377. PMID 7781615.
- Bell SP, Stillman B (May 1992). "ATP-dependent recognition of eukaryotic origins of DNA replication by a multiprotein complex". Nature. 357 (6374): 128–34. Bibcode:1992Natur.357..128B. doi:10.1038/357128a0. PMID 1579162. S2CID 4346767.
- Li N, Lam WH, Zhai Y, Cheng J, Cheng E, Zhao Y, et al. (July 2018). "Structure of the origin recognition complex bound to DNA replication origin". Nature. 559 (7713): 217–222. Bibcode:2018Natur.559..217L. doi:10.1038/s41586-018-0293-x. PMID 29973722. S2CID 49577101.
- Bleichert F, Botchan MR, Berger JM (March 2015). "Crystal structure of the eukaryotic origin recognition complex". Nature. 519 (7543): 321–6. Bibcode:2015Natur.519..321B. doi:10.1038/nature14239. PMC 4368505. PMID 25762138.
- Sun J, Evrin C, Samel SA, Fernández-Cid A, Riera A, Kawakami H, et al. (August 2013). "Cryo-EM structure of a helicase loading intermediate containing ORC-Cdc6-Cdt1-MCM2-7 bound to DNA". Nature Structural & Molecular Biology. 20 (8): 944–51. doi:10.1038/nsmb.2629. PMC 3735830. PMID 23851460.
- Kawakami H, Ohashi E, Kanamoto S, Tsurimoto T, Katayama T (October 2015). "Specific binding of eukaryotic ORC to DNA replication origins depends on highly conserved basic residues". Scientific Reports. 5: 14929. Bibcode:2015NatSR...514929K. doi:10.1038/srep14929. PMC 4601075. PMID 26456755.
- Palzkill TG, Newlon CS (May 1988). "A yeast replication origin consists of multiple copies of a small conserved sequence". Cell. 53 (3): 441–50. doi:10.1016/0092-8674(88)90164-x. PMID 3284655. S2CID 7534654.
- Wilmes GM, Bell SP (January 2002). "The B2 element of the Saccharomyces cerevisiae ARS1 origin of replication requires specific sequences to facilitate pre-RC formation". Proceedings of the National Academy of Sciences of the United States of America. 99 (1): 101–6. Bibcode:2002PNAS...99..101W. doi:10.1073/pnas.012578499. PMC 117521. PMID 11756674.
- Coster G, Diffley JF (July 2017). "Bidirectional eukaryotic DNA replication is established by quasi-symmetrical helicase loading". Science. 357 (6348): 314–318. Bibcode:2017Sci...357..314C. doi:10.1126/science.aan0063. PMC 5608077. PMID 28729513.
- Zou L, Stillman B (May 2000). "Assembly of a complex containing Cdc45p, replication protein A, and Mcm2p at replication origins controlled by S-phase cyclin-dependent kinases and Cdc7p-Dbf4p kinase". Molecular and Cellular Biology. 20 (9): 3086–96. doi:10.1128/mcb.20.9.3086-3096.2000. PMC 85601. PMID 10757793.
- Lipford JR, Bell SP (January 2001). "Nucleosomes positioned by ORC facilitate the initiation of DNA replication". Molecular Cell. 7 (1): 21–30. doi:10.1016/s1097-2765(01)00151-4. PMID 11172708.
- Diffley JF, Cocker JH (May 1992). "Protein-DNA interactions at a yeast replication origin". Nature. 357 (6374): 169–72. Bibcode:1992Natur.357..169D. doi:10.1038/357169a0. PMID 1579168. S2CID 4354585.
- Diffley JF, Stillman B (April 1988). "Purification of a yeast protein that binds to origins of DNA replication and a transcriptional silencer". Proceedings of the National Academy of Sciences of the United States of America. 85 (7): 2120–4. Bibcode:1988PNAS...85.2120D. doi:10.1073/pnas.85.7.2120. PMC 279940. PMID 3281162.
- Miotto B, Ji Z, Struhl K (August 2016). "Selectivity of ORC binding sites and the relation to replication timing, fragile sites, and deletions in cancers". Proceedings of the National Academy of Sciences of the United States of America. 113 (33): E4810-9. doi:10.1073/pnas.1609060113. PMC 4995967. PMID 27436900.
- MacAlpine HK, Gordân R, Powell SK, Hartemink AJ, MacAlpine DM (February 2010). "Drosophila ORC localizes to open chromatin and marks sites of cohesin complex loading". Genome Research. 20 (2): 201–11. doi:10.1101/gr.097873.109. PMC 2813476. PMID 19996087.
- Eaton ML, Prinz JA, MacAlpine HK, Tretyakov G, Kharchenko PV, MacAlpine DM (February 2011). "Chromatin signatures of the Drosophila replication program". Genome Research. 21 (2): 164–74. doi:10.1101/gr.116038.110. PMC 3032920. PMID 21177973.
- Dellino GI, Cittaro D, Piccioni R, Luzi L, Banfi S, Segalla S, et al. (January 2013). "Genome-wide mapping of human DNA-replication origins: levels of transcription at ORC1 sites regulate origin selection and replication timing". Genome Research. 23 (1): 1–11. doi:10.1101/gr.142331.112. PMC 3530669. PMID 23187890.
- Cayrou C, Ballester B, Peiffer I, Fenouil R, Coulombe P, Andrau JC, et al. (December 2015). "The chromatin environment shapes DNA replication origin organization and defines origin classes". Genome Research. 25 (12): 1873–85. doi:10.1101/gr.192799.115. PMC 4665008. PMID 26560631.
- Cayrou C, Coulombe P, Vigneron A, Stanojcic S, Ganier O, Peiffer I, et al. (September 2011). "Genome-scale analysis of metazoan replication origins reveals their organization in specific but flexible sites defined by conserved features". Genome Research. 21 (9): 1438–49. doi:10.1101/gr.121830.111. PMC 3166829. PMID 21750104.
- Lubelsky Y, Sasaki T, Kuipers MA, Lucas I, Le Beau MM, Carignon S, et al. (April 2011). "Pre-replication complex proteins assemble at regions of low nucleosome occupancy within the Chinese hamster dihydrofolate reductase initiation zone". Nucleic Acids Research. 39 (8): 3141–55. doi:10.1093/nar/gkq1276. PMC 3082903. PMID 21148149.
- Hayashi M, Katou Y, Itoh T, Tazumi A, Tazumi M, Yamada Y, et al. (March 2007). "Genome-wide localization of pre-RC sites and identification of replication origins in fission yeast". The EMBO Journal. 26 (5): 1327–39. doi:10.1038/sj.emboj.7601585. PMC 1817633. PMID 17304213.
- Martin MM, Ryan M, Kim R, Zakas AL, Fu H, Lin CM, et al. (November 2011). "Genome-wide depletion of replication initiation events in highly transcribed regions". Genome Research. 21 (11): 1822–32. doi:10.1101/gr.124644.111. PMC 3205567. PMID 21813623.
- Pourkarimi E, Bellush JM, Whitehouse I (December 2016). "C. elegans". eLife. 5. doi:10.7554/eLife.21728. PMC 5222557. PMID 28009254.
- Rodríguez-Martínez M, Pinzón N, Ghommidh C, Beyne E, Seitz H, Cayrou C, Méchali M (March 2017). "The gastrula transition reorganizes replication-origin selection in Caenorhabditis elegans". Nature Structural & Molecular Biology. 24 (3): 290–299. doi:10.1038/nsmb.3363. PMID 28112731. S2CID 7445974.
- Besnard E, Babled A, Lapasset L, Milhavet O, Parrinello H, Dantec C, et al. (August 2012). "Unraveling cell type-specific and reprogrammable human replication origin signatures associated with G-quadruplex consensus motifs". Nature Structural & Molecular Biology. 19 (8): 837–44. doi:10.1038/nsmb.2339. PMID 22751019. S2CID 20710237.
- Delgado S, Gómez M, Bird A, Antequera F (April 1998). "Initiation of DNA replication at CpG islands in mammalian chromosomes". The EMBO Journal. 17 (8): 2426–35. doi:10.1093/emboj/17.8.2426. PMC 1170585. PMID 9545253.
- Sequeira-Mendes J, Díaz-Uriarte R, Apedaile A, Huntley D, Brockdorff N, Gómez M (April 2009). "Transcription initiation activity sets replication origin efficiency in mammalian cells". PLOS Genetics. 5 (4): e1000446. doi:10.1371/journal.pgen.1000446. PMC 2661365. PMID 19360092.
- Kelly T, Callegari AJ (March 2019). "Dynamics of DNA replication in a eukaryotic cell". Proceedings of the National Academy of Sciences of the United States of America. 116 (11): 4973–4982. doi:10.1073/pnas.1818680116. PMC 6421431. PMID 30718387.
- Austin RJ, Orr-Weaver TL, Bell SP (October 1999). "Drosophila ORC specifically binds to ACE3, an origin of DNA replication control element". Genes & Development. 13 (20): 2639–49. doi:10.1101/gad.13.20.2639. PMC 317108. PMID 10541550.
- Beall EL, Manak JR, Zhou S, Bell M, Lipsick JS, Botchan MR (2002). "Role for a Drosophila Myb-containing protein complex in site-specific DNA replication". Nature. 420 (6917): 833–7. Bibcode:2002Natur.420..833B. doi:10.1038/nature01228. PMID 12490953. S2CID 4425307.
- Beall EL, Bell M, Georlette D, Botchan MR (July 2004). "Dm-myb mutant lethality in Drosophila is dependent upon mip130: positive and negative regulation of DNA replication". Genes & Development. 18 (14): 1667–80. doi:10.1101/gad.1206604. PMC 478189. PMID 15256498.
- Lewis PW, Beall EL, Fleischer TC, Georlette D, Link AJ, Botchan MR (December 2004). "Identification of a Drosophila Myb-E2F2/RBF transcriptional repressor complex". Genes & Development. 18 (23): 2929–40. doi:10.1101/gad.1255204. PMC 534653. PMID 15545624.
- Bosco G, Du W, Orr-Weaver TL (March 2001). "DNA replication control through interaction of E2F-RB and the origin recognition complex". Nature Cell Biology. 3 (3): 289–95. doi:10.1038/35060086. PMID 11231579. S2CID 24942902.
- Chuang RY, Kelly TJ (March 1999). "The fission yeast homologue of Orc4p binds to replication origin DNA via multiple AT-hooks". Proceedings of the National Academy of Sciences of the United States of America. 96 (6): 2656–61. Bibcode:1999PNAS...96.2656C. doi:10.1073/pnas.96.6.2656. PMC 15824. PMID 10077566.
- Balasov M, Huijbregts RP, Chesnokov I (April 2007). "Role of the Orc6 protein in origin recognition complex-dependent DNA binding and replication in Drosophila melanogaster". Molecular and Cellular Biology. 27 (8): 3143–53. doi:10.1128/MCB.02382-06. PMC 1899928. PMID 17283052.
- Tardat M, Brustel J, Kirsh O, Lefevbre C, Callanan M, Sardet C, Julien E (November 2010). "The histone H4 Lys 20 methyltransferase PR-Set7 regulates replication origins in mammalian cells". Nature Cell Biology. 12 (11): 1086–93. doi:10.1038/ncb2113. PMID 20953199. S2CID 6710289.
- Beck DB, Burton A, Oda H, Ziegler-Birling C, Torres-Padilla ME, Reinberg D (December 2012). "The role of PR-Set7 in replication licensing depends on Suv4-20h". Genes & Development. 26 (23): 2580–9. doi:10.1101/gad.195636.112. PMC 3521623. PMID 23152447.
- Brustel J, Kirstein N, Izard F, Grimaud C, Prorok P, Cayrou C, et al. (September 2017). "Histone H4K20 tri-methylation at late-firing origins ensures timely heterochromatin replication". The EMBO Journal. 36 (18): 2726–2741. doi:10.15252/embj.201796541. PMC 5599798. PMID 28778956.
- Shoaib M, Walter D, Gillespie PJ, Izard F, Fahrenkrog B, Lleres D, et al. (September 2018). "Histone H4K20 methylation mediated chromatin compaction threshold ensures genome integrity by limiting DNA replication licensing". Nature Communications. 9 (1): 3704. Bibcode:2018NatCo...9.3704S. doi:10.1038/s41467-018-06066-8. PMC 6135857. PMID 30209253.
- Noguchi K, Vassilev A, Ghosh S, Yates JL, DePamphilis ML (November 2006). "The BAH domain facilitates the ability of human Orc1 protein to activate replication origins in vivo". The EMBO Journal. 25 (22): 5372–82. doi:10.1038/sj.emboj.7601396. PMC 1636626. PMID 17066079.
- Shen Z, Chakraborty A, Jain A, Giri S, Ha T, Prasanth KV, Prasanth SG (August 2012). "Dynamic association of ORCA with prereplicative complex components regulates DNA replication initiation". Molecular and Cellular Biology. 32 (15): 3107–20. doi:10.1128/MCB.00362-12. PMC 3434513. PMID 22645314.
- Wang Y, Khan A, Marks AB, Smith OK, Giri S, Lin YC, et al. (March 2017). "Temporal association of ORCA/LRWD1 to late-firing origins during G1 dictates heterochromatin replication and organization". Nucleic Acids Research. 45 (5): 2490–2502. doi:10.1093/nar/gkw1211. PMC 5389698. PMID 27924004.
- Bartke T, Vermeulen M, Xhemalce B, Robson SC, Mann M, Kouzarides T (October 2010). "Nucleosome-interacting proteins regulated by DNA and histone methylation". Cell. 143 (3): 470–84. doi:10.1016/j.cell.2010.10.012. PMC 3640253. PMID 21029866.
- Vermeulen M, Eberl HC, Matarese F, Marks H, Denissov S, Butter F, et al. (September 2010). "Quantitative interaction proteomics and genome-wide profiling of epigenetic histone marks and their readers". Cell. 142 (6): 967–80. doi:10.1016/j.cell.2010.08.020. PMID 20850016. S2CID 7926456.
- Hein MY, Hubner NC, Poser I, Cox J, Nagaraj N, Toyoda Y, et al. (October 2015). "A human interactome in three quantitative dimensions organized by stoichiometries and abundances". Cell. 163 (3): 712–23. doi:10.1016/j.cell.2015.09.053. PMID 26496610.
- Thomae AW, Pich D, Brocher J, Spindler MP, Berens C, Hock R, et al. (February 2008). "Interaction between HMGA1a and the origin recognition complex creates site-specific replication origins". Proceedings of the National Academy of Sciences of the United States of America. 105 (5): 1692–7. Bibcode:2008PNAS..105.1692T. doi:10.1073/pnas.0707260105. PMC 2234206. PMID 18234858.
- Zhang Y, Huang L, Fu H, Smith OK, Lin CM, Utani K, et al. (June 2016). "A replicator-specific binding protein essential for site-specific initiation of DNA replication in mammalian cells". Nature Communications. 7: 11748. Bibcode:2016NatCo...711748Z. doi:10.1038/ncomms11748. PMC 4899857. PMID 27272143.
- Bleichert F, Leitner A, Aebersold R, Botchan MR, Berger JM (June 2018). "Conformational control and DNA-binding mechanism of the metazoan origin recognition complex". Proceedings of the National Academy of Sciences of the United States of America. 115 (26): E5906–E5915. doi:10.1073/pnas.1806315115. PMC 6042147. PMID 29899147.
- Clarey MG, Botchan M, Nogales E (December 2008). "Single particle EM studies of the Drosophila melanogaster origin recognition complex and evidence for DNA wrapping". Journal of Structural Biology. 164 (3): 241–9. doi:10.1016/j.jsb.2008.08.006. PMC 2640233. PMID 18824234.
- Lee DG, Bell SP (December 1997). "Architecture of the yeast origin recognition complex bound to origins of DNA replication". Molecular and Cellular Biology. 17 (12): 7159–68. doi:10.1128/mcb.17.12.7159. PMC 232573. PMID 9372948.
- Riera A, Barbon M, Noguchi Y, Reuter LM, Schneider S, Speck C (June 2017). "From structure to mechanism-understanding initiation of DNA replication". Genes & Development. 31 (11): 1073–1088. doi:10.1101/gad.298232.117. PMC 5538431. PMID 28717046.
- Tognetti S, Riera A, Speck C (March 2015). "Switch on the engine: how the eukaryotic replicative helicase MCM2-7 becomes activated". Chromosoma. 124 (1): 13–26. doi:10.1007/s00412-014-0489-2. hdl:10044/1/27085. PMID 25308420. S2CID 175510.
- Berbenetz NM, Nislow C, Brown GW (September 2010). "Diversity of eukaryotic DNA replication origins revealed by genome-wide analysis of chromatin structure". PLOS Genetics. 6 (9): e1001092. doi:10.1371/journal.pgen.1001092. PMC 2932696. PMID 20824081.
- Eaton ML, Galani K, Kang S, Bell SP, MacAlpine DM (April 2010). "Conserved nucleosome positioning defines replication origins". Genes & Development. 24 (8): 748–53. doi:10.1101/gad.1913210. PMC 2854390. PMID 20351051.
- Azmi IF, Watanabe S, Maloney MF, Kang S, Belsky JA, MacAlpine DM, et al. (March 2017). "Nucleosomes influence multiple steps during replication initiation". eLife. 6. doi:10.7554/eLife.22512. PMC 5400510. PMID 28322723.
- Miotto B, Struhl K (January 2010). "HBO1 histone acetylase activity is essential for DNA replication licensing and inhibited by Geminin". Molecular Cell. 37 (1): 57–66. doi:10.1016/j.molcel.2009.12.012. PMC 2818871. PMID 20129055.
- Liu J, Zimmer K, Rusch DB, Paranjape N, Podicheti R, Tang H, Calvi BR (October 2015). "DNA sequence templates adjacent nucleosome and ORC sites at gene amplification origins in Drosophila". Nucleic Acids Research. 43 (18): 8746–61. doi:10.1093/nar/gkv766. PMC 4605296. PMID 26227968.
- Zhao PA, Rivera-Mulia JC, Gilbert DM (2017). "Replication Domains: Genome Compartmentalization into Functional Replication Units". Advances in Experimental Medicine and Biology. 1042: 229–257. doi:10.1007/978-981-10-6955-0_11. ISBN 978-981-10-6954-3. PMID 29357061.
- Sugimoto N, Fujita M (2017). "Molecular Mechanism for Chromatin Regulation During MCM Loading in Mammalian Cells". Advances in Experimental Medicine and Biology. 1042: 61–78. doi:10.1007/978-981-10-6955-0_3. ISBN 978-981-10-6954-3. PMID 29357053.
- MacAlpine DM, Almouzni G (August 2013). "Chromatin and DNA replication". Cold Spring Harbor Perspectives in Biology. 5 (8): a010207. doi:10.1101/cshperspect.a010207. PMC 3721285. PMID 23751185.
- Sima J, Chakraborty A, Dileep V, Michalski M, Klein KN, Holcomb NP, et al. (February 2019). "Identifying cis Elements for Spatiotemporal Control of Mammalian DNA Replication". Cell. 176 (4): 816–830.e18. doi:10.1016/j.cell.2018.11.036. PMC 6546437. PMID 30595451.
- Cadoret JC, Meisch F, Hassan-Zadeh V, Luyten I, Guillet C, Duret L, et al. (October 2008). "Genome-wide studies highlight indirect links between human replication origins and gene regulation". Proceedings of the National Academy of Sciences of the United States of America. 105 (41): 15837–42. Bibcode:2008PNAS..10515837C. doi:10.1073/pnas.0805208105. PMC 2572913. PMID 18838675.
- Azvolinsky A, Giresi PG, Lieb JD, Zakian VA (June 2009). "Highly transcribed RNA polymerase II genes are impediments to replication fork progression in Saccharomyces cerevisiae". Molecular Cell. 34 (6): 722–34. doi:10.1016/j.molcel.2009.05.022. PMC 2728070. PMID 19560424.
- Gros J, Kumar C, Lynch G, Yadav T, Whitehouse I, Remus D (December 2015). "Post-licensing Specification of Eukaryotic Replication Origins by Facilitated Mcm2-7 Sliding along DNA". Molecular Cell. 60 (5): 797–807. doi:10.1016/j.molcel.2015.10.022. PMC 4680849. PMID 26656162.
- Letessier A, Millot GA, Koundrioukoff S, Lachagès AM, Vogt N, Hansen RS, et al. (February 2011). "Cell-type-specific replication initiation programs set fragility of the FRA3B fragile site". Nature. 470 (7332): 120–3. Bibcode:2011Natur.470..120L. doi:10.1038/nature09745. PMID 21258320. S2CID 4302940.
- Smith OK, Kim R, Fu H, Martin MM, Lin CM, Utani K, et al. (2016). "Distinct epigenetic features of differentiation-regulated replication origins". Epigenetics & Chromatin. 9: 18. doi:10.1186/s13072-016-0067-3. PMC 4862150. PMID 27168766.
- Sher N, Bell GW, Li S, Nordman J, Eng T, Eaton ML, et al. (January 2012). "Developmental control of gene copy number by repression of replication initiation and fork progression". Genome Research. 22 (1): 64–75. doi:10.1101/gr.126003.111. PMC 3246207. PMID 22090375.
- Comoglio F, Schlumpf T, Schmid V, Rohs R, Beisel C, Paro R (May 2015). "High-resolution profiling of Drosophila replication start sites reveals a DNA shape and chromatin signature of metazoan origins". Cell Reports. 11 (5): 821–34. doi:10.1016/j.celrep.2015.03.070. PMC 4562395. PMID 25921534.
- Calvi BR, Lilly MA, Spradling AC (March 1998). "Cell cycle control of chorion gene amplification". Genes & Development. 12 (5): 734–44. doi:10.1101/gad.12.5.734. PMC 316579. PMID 9499407.
- Mosig G (1998). "Recombination and recombination-dependent DNA replication in bacteriophage T4". Annual Review of Genetics. 32: 379–413. doi:10.1146/annurev.genet.32.1.379. PMID 9928485.
- Ravoitytė B, Wellinger RE (January 2017). "Non-Canonical Replication Initiation: You're Fired!". Genes. 8 (2): 54. doi:10.3390/genes8020054. PMC 5333043. PMID 28134821.
- Asai T, Sommer S, Bailone A, Kogoma T (August 1993). "Homologous recombination-dependent initiation of DNA replication from DNA damage-inducible origins in Escherichia coli". The EMBO Journal. 12 (8): 3287–95. doi:10.1002/j.1460-2075.1993.tb05998.x. PMC 413596. PMID 8344265.
- Lydeard JR, Jain S, Yamaguchi M, Haber JE (August 2007). "Break-induced replication and telomerase-independent telomere maintenance require Pol32". Nature. 448 (7155): 820–3. Bibcode:2007Natur.448..820L. doi:10.1038/nature06047. PMID 17671506. S2CID 4373857.
- Dasgupta S, Masukata H, Tomizawa J (December 1987). "Multiple mechanisms for initiation of ColE1 DNA replication: DNA synthesis in the presence and absence of ribonuclease H". Cell. 51 (6): 1113–22. doi:10.1016/0092-8674(87)90597-6. PMID 2446774. S2CID 22858038.
- Stuckey R, García-Rodríguez N, Aguilera A, Wellinger RE (May 2015). "Role for RNA:DNA hybrids in origin-independent replication priming in a eukaryotic system". Proceedings of the National Academy of Sciences of the United States of America. 112 (18): 5779–84. Bibcode:2015PNAS..112.5779S. doi:10.1073/pnas.1501769112. PMC 4426422. PMID 25902524.
- Burki F (May 2014). "The eukaryotic tree of life from a global phylogenomic perspective". Cold Spring Harbor Perspectives in Biology. 6 (5): a016147. doi:10.1101/cshperspect.a016147. PMC 3996474. PMID 24789819.
- Lee PH, Meng X, Kapler GM (January 2015). "Developmental regulation of the Tetrahymena thermophila origin recognition complex". PLOS Genetics. 11 (1): e1004875. doi:10.1371/journal.pgen.1004875. PMC 4287346. PMID 25569357.
- Mohammad MM, Donti TR, Sebastian Yakisich J, Smith AG, Kapler GM (December 2007). "Tetrahymena ORC contains a ribosomal RNA fragment that participates in rDNA origin recognition". The EMBO Journal. 26 (24): 5048–60. doi:10.1038/sj.emboj.7601919. PMC 2140106. PMID 18007594.
- Donti TR, Datta S, Sandoval PY, Kapler GM (February 2009). "Differential targeting of Tetrahymena ORC to ribosomal DNA and non-rDNA replication origins". The EMBO Journal. 28 (3): 223–33. doi:10.1038/emboj.2008.282. PMC 2637336. PMID 19153611.
- Marques CA, McCulloch R (February 2018). "Conservation and Variation in Strategies for DNA Replication of Kinetoplastid Nuclear Genomes". Current Genomics. 19 (2): 98–109. doi:10.2174/1389202918666170815144627. PMC 5814967. PMID 29491738.
- Marques CA, Tiengwe C, Lemgruber L, Damasceno JD, Scott A, Paape D, et al. (June 2016). "Diverged composition and regulation of the Trypanosoma brucei origin recognition complex that mediates DNA replication initiation". Nucleic Acids Research. 44 (10): 4763–84. doi:10.1093/nar/gkw147. PMC 4889932. PMID 26951375.
- Tiengwe C, Marcello L, Farr H, Gadelha C, Burchmore R, Barry JD, et al. (2012). "Identification of ORC1/CDC6-interacting factors in Trypanosoma brucei reveals critical features of origin recognition complex architecture". PLOS ONE. 7 (3): e32674. Bibcode:2012PLoSO...732674T. doi:10.1371/journal.pone.0032674. PMC 3297607. PMID 22412905.
- Marques CA, Dickens NJ, Paape D, Campbell SJ, McCulloch R (October 2015). "Genome-wide mapping reveals single-origin chromosome replication in Leishmania, a eukaryotic microbe". Genome Biology. 16: 230. doi:10.1186/s13059-015-0788-9. PMC 4612428. PMID 26481451.
Further reading
- Lewin B (2004). Genes VIII. Prentice Hall. ISBN 978-0-13-144945-9.
External links
- Ori-Finder, an online software for prediction of bacterial and archaeal oriCs
- Replication+Origin at the US National Library of Medicine Medical Subject Headings (MeSH)