Telomerase
Telomerase, also called terminal transferase,[1] is a ribonucleoprotein that adds a species-dependent telomere repeat sequence to the 3' end of telomeres. A telomere is a region of repetitive sequences at each end of the chromosomes of most eukaryotes. Telomeres protect the end of the chromosome from DNA damage or from fusion with neighbouring chromosomes. The fruit fly Drosophila melanogaster lacks telomerase, but instead uses retrotransposons to maintain telomeres.[2]


Telomerase is a reverse transcriptase enzyme that carries its own RNA molecule (e.g., with the sequence 3′-CCCAAUCCC-5′ in Trypanosoma brucei)[3] which is used as a template when it elongates telomeres. Telomerase is active in gametes and most cancer cells, but is normally absent from, or at very low levels in, most somatic cells.
History
The existence of a compensatory mechanism for telomere shortening was first found by Soviet biologist Alexey Olovnikov in 1973,[4] who also suggested the telomere hypothesis of aging and the telomere's connections to cancer.
Telomerase in the ciliate Tetrahymena was discovered by Carol W. Greider and Elizabeth Blackburn in 1984.[5] Together with Jack W. Szostak, Greider and Blackburn were awarded the 2009 Nobel Prize in Physiology or Medicine for their discovery.[6]
The role of telomeres and telomerase in cell aging and cancer was established by scientists at biotechnology company Geron with the cloning of the RNA and catalytic components of human telomerase[7] and the development of a polymerase chain reaction (PCR) based assay for telomerase activity called the TRAP assay, which surveys telomerase activity in multiple types of cancer.[8]
The negative stain electron microscopy (EM) structures of human and Tetrahymena telomerases were characterized in 2013.[9][10] Two years later, the first cryo-electron microscopy (cryo-EM) structure of telomerase holoenzyme (Tetrahymena) was determined.[11] In 2018, the structure of human telomerase was determined through cryo-EM by UC Berkeley scientists.[12]
Human telomerase structure
The molecular composition of the human telomerase complex was determined by Scott Cohen and his team at the Children's Medical Research Institute (Sydney Australia) and consists of two molecules each of human telomerase reverse transcriptase (TERT), telomerase RNA (TR or TERC), and dyskerin (DKC1).[13] The genes of telomerase subunits, which include TERT,[14] TERC,[15] DKC1[16] and TEP1,[17] are located on different chromosomes. The human TERT gene (hTERT) is translated into a protein of 1132 amino acids.[18] TERT polypeptide folds with (and carries) TERC, a non-coding RNA (451 nucleotides long). TERT has a 'mitten' structure that allows it to wrap around the chromosome to add single-stranded telomere repeats.
TERT is a reverse transcriptase, which is a class of enzymes that creates single-stranded DNA using single-stranded RNA as a template.
The protein consists of four conserved domains (RNA-Binding Domain (TRBD), fingers, palm and thumb), organized into a "right hand" ring configuration that shares common features with retroviral reverse transcriptases, viral RNA replicases and bacteriophage B-family DNA polymerases.[19][20]
TERT proteins from many eukaryotes have been sequenced.[21]
Mechanism
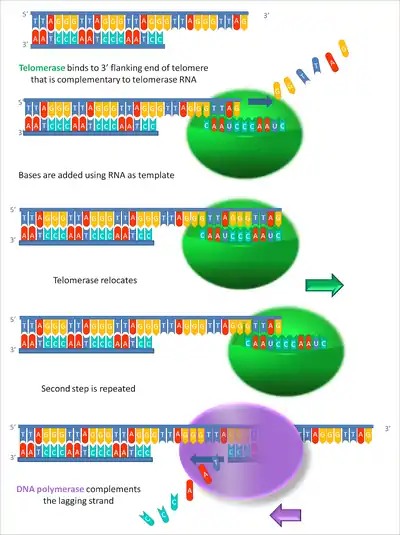
By using TERC, TERT can add a six-nucleotide repeating sequence, 5'-TTAGGG (in vertebrates, the sequence differs in other organisms) to the 3' strand of chromosomes. These TTAGGG repeats (with their various protein binding partners) are called telomeres. The template region of TERC is 3'-CAAUCCCAAUC-5'.[22]
Telomerase can bind the first few nucleotides of the template to the last telomere sequence on the chromosome, add a new telomere repeat (5'-GGTTAG-3') sequence, let go, realign the new 3'-end of telomere to the template, and repeat the process. Telomerase reverses telomere shortening.
Clinical implications
Aging
Telomerase restores short bits of DNA known as telomeres, which are otherwise shortened after repeated division of a cell via mitosis.
In normal circumstances, where telomerase is absent, if a cell divides recursively, at some point the progeny reach their Hayflick limit,[23] which is believed to be between 50 and 70 cell divisions. At the limit the cells become senescent and cell division stops.[24] Telomerase allows each offspring to replace the lost bit of DNA, allowing the cell line to divide without ever reaching the limit. This same unbounded growth is a feature of cancerous growth.[25]
Embryonic stem cells express telomerase, which allows them to divide repeatedly and form the individual. In adults, telomerase is highly expressed only in cells that need to divide regularly, especially in male sperm cells,[26] but also in epidermal cells,[27] in activated T cell[28] and B cell[29] lymphocytes, as well as in certain adult stem cells, but in the great majority of cases somatic cells do not express telomerase.[30]
A comparative biology study of mammalian telomeres indicated that telomere length of some mammalian species correlates inversely, rather than directly, with lifespan, and concluded that the contribution of telomere length to lifespan is unresolved.[31] Telomere shortening does not occur with age in some postmitotic tissues, such as in the rat brain.[32] In humans, skeletal muscle telomere lengths remain stable from ages 23 –74.[33] In baboon skeletal muscle, which consists of fully differentiated postmitotic cells, less than 3% of myonuclei contain damaged telomeres and this percentage does not increase with age.[34] Thus, telomere shortening does not appear to be a major factor in the aging of the differentiated cells of brain or skeletal muscle. In human liver, cholangiocytes and hepatocytes show no age-related telomere shortening.[35] Another study found little evidence that, in humans, telomere length is a significant biomarker of normal aging with respect to important cognitive and physical abilities.[36]
Some experiments have raised questions on whether telomerase can be used as an anti-aging therapy, namely, the fact that mice with elevated levels of telomerase have higher cancer incidence and hence do not live longer.[37] On the other hand, one study showed that activating telomerase in cancer-resistant mice by overexpressing its catalytic subunit extended lifespan.[38] A study found that long-lived subjects inherited a hyperactive version of telomerase.[39]
Premature aging
Premature aging syndromes including Werner syndrome, Progeria, Ataxia telangiectasia, Ataxia-telangiectasia like disorder, Bloom syndrome, Fanconi anemia and Nijmegen breakage syndrome are associated with short telomeres.[40] However, the genes that have mutated in these diseases all have roles in the repair of DNA damage and the increased DNA damage may, itself, be a factor in the premature aging (see DNA damage theory of aging). An additional role in maintaining telomere length is an active area of investigation.
Cancer
In vitro, when cells approach the Hayflick limit, the time to senescence can be extended by inactivating the tumor suppressor proteins - p53 and Retinoblastoma protein (pRb).[41] Cells that have been so-altered eventually undergo an event termed a "crisis" when the majority of the cells in the culture die. Sometimes, a cell does not stop dividing once it reaches a crisis. In a typical situation, the telomeres are shortened[42] and chromosomal integrity declines with every subsequent cell division. Exposed chromosome ends are interpreted as double-stranded breaks (DSB) in DNA; such damage is usually repaired by reattaching the broken ends together. When the cell does this due to telomere-shortening, the ends of different chromosomes can be attached to each other. This solves the problem of lacking telomeres, but during cell division anaphase, the fused chromosomes are randomly ripped apart, causing many mutations and chromosomal abnormalities. As this process continues, the cell's genome becomes unstable. Eventually, either fatal damage is done to the cell's chromosomes (killing it via apoptosis), or an additional mutation that activates telomerase occurs.[41]
With telomerase activation some types of cells and their offspring become immortal (bypass the Hayflick limit), thus avoiding cell death as long as the conditions for their duplication are met. Many cancer cells are considered 'immortal' because telomerase activity allows them to live much longer than any other somatic cell, which, combined with uncontrollable cell proliferation[43] is why they can form tumors. A good example of immortal cancer cells is HeLa cells, which have been used in laboratories as a model cell line since 1951.
While this method of modelling human cancer in cell culture is effective and has been used for many years by scientists, it is also very imprecise. The exact changes that allow for the formation of the tumorigenic clones in the above-described experiment are not clear. Scientists addressed this question by the serial introduction of multiple mutations present in a variety of human cancers. This has led to the identification of mutation combinations that form tumorigenic cells in a variety of cell types. While the combination varies by cell type, the following alterations are required in all cases: TERT activation, loss of p53 pathway function, loss of pRb pathway function, activation of the Ras or myc proto-oncogenes, and aberration of the PP2A protein phosphatase.[44] That is to say, the cell has an activated telomerase, eliminating the process of death by chromosome instability or loss, absence of apoptosis-induction pathways, and continued mitosis activation.
This model of cancer in cell culture accurately describes the role of telomerase in actual human tumors. Telomerase activation has been observed in ~90% of all human tumors,[45] suggesting that the immortality conferred by telomerase plays a key role in cancer development. Of the tumors without TERT activation,[46] most employ a separate pathway to maintain telomere length termed Alternative Lengthening of Telomeres (ALT).[47] The exact mechanism behind telomere maintenance in the ALT pathway is unclear, but likely involves multiple recombination events at the telomere.
Elizabeth Blackburn et al., identified the upregulation of 70 genes known or suspected in cancer growth and spread through the body, and the activation of glycolysis, which enables cancer cells to rapidly use sugar to facilitate their programmed growth rate (roughly the growth rate of a fetus).[48]
Approaches to controlling telomerase and telomeres for cancer therapy include gene therapy, immunotherapy, small-molecule and signal pathway inhibitors.[49]
Drugs
The ability to maintain functional telomeres may be one mechanism that allows cancer cells to grow in vitro for decades.[50] Telomerase activity is necessary to preserve many cancer types and is inactive in somatic cells, creating the possibility that telomerase inhibition could selectively repress cancer cell growth with minimal side effects.[51] If a drug can inhibit telomerase in cancer cells, the telomeres of successive generations will progressively shorten, limiting tumor growth.[52]
Telomerase is a good biomarker for cancer detection because most human cancers cells express high levels of it. Telomerase activity can be identified by its catalytic protein domain (hTERT). This is the rate-limiting step in telomerase activity. It is associated with many cancer types. Various cancer cells and fibroblasts transformed with hTERT cDNA have high telomerase activity, while somatic cells do not. Cells testing positive for hTERT have positive nuclear signals. Epithelial stem cell tissue and its early daughter cells are the only noncancerous cells in which hTERT can be detected. Since hTERT expression is dependent only on the number of tumor cells within a sample, the amount of hTERT indicates the severity of cancer.[53]
The expression of hTERT can also be used to distinguish benign tumors from malignant tumors. Malignant tumors have higher hTERT expression than benign tumors. Real-time reverse transcription polymerase chain reaction (RT-PCR) quantifying hTERT expression in various tumor samples verified this varying expression.[54]

The lack of telomerase does not affect cell growth until the telomeres are short enough to cause cells to “die or undergo growth arrest”. However, inhibiting telomerase alone is not enough to destroy large tumors. It must be combined with surgery, radiation, chemotherapy or immunotherapy.[53]
Cells may reduce their telomere length by only 50-252 base pairs per cell division, which can lead to a long lag phase.[55][56]
A telomerase activator TA-65 is commercially available and is claimed to delay aging and to provide relief from certain disease conditions.[57][58][59][60][61] This formulation contains a molecule called cycloastragenol derived from a legume Astragalus membranaceus, which has been used in Chinese traditional medicine for extending healthy life.
Immunotherapy
Immunotherapy successfully treats some kinds of cancer, such as melanoma. This treatment involves manipulating a human's immune system to destroy cancerous cells. Humans have two major antigen identifying lymphocytes: CD8+ cytotoxic T-lymphocytes (CTL) and CD4+ helper T-lymphocytes that can destroy cells. Antigen receptors on CTL can bind to a 9-10 amino acid chain that is presented by the major histocompatibility complex (MHC) as in Figure 4. HTERT is a potential target antigen. Immunotargeting should result in relatively few side effects since hTERT expression is associated only with telomerase and is not essential in almost all somatic cells.[62] GV1001 uses this pathway.[49] Experimental drug and vaccine therapies targeting active telomerase have been tested in mouse models, and clinical trials have begun. One drug, imetelstat, is being clinically researched as a means of interfering with telomerase in cancer cells.[63] Most of the harmful cancer-related effects of telomerase are dependent on an intact RNA template. Cancer stem cells that use an alternative method of telomere maintenance are still killed when telomerase's RNA template is blocked or damaged.
Telomerase Vaccines
Two telomerase vaccines have been developed: GRNVAC1 and GV1001. GRNVAC1 isolates dendritic cells and the RNA that codes for the telomerase protein and puts them back into the patient to make cytotoxic T cells that kill the telomerase-active cells. GV1001 is a peptide from the active site of hTERT and is recognized by the immune system that reacts by killing the telomerase-active cells.[49]
Targeted apoptosis

Another independent approach is to use oligoadenylated anti-telomerase antisense oligonucleotides and ribozymes to target telomerase RNA, reducing dissociation and apoptosis (Figure 5). The fast induction of apoptosis through antisense binding may be a good alternative to the slower telomere shortening.[55]
Small interfering RNA (siRNA)
siRNAs are small RNA molecules that induce the sequence-specific degradation of other RNAs. siRNA treatment can function similar to traditional gene therapy by destroying the mRNA products of particular genes, and therefore preventing the expression of those genes. A 2012 study found that targeting TERC with an siRNA reduced telomerase activity by more than 50% and resulted in decreased viability of immortal cancer cells.[64] Treatment with both the siRNA and radiation caused a greater reduction in tumor size in mice than treatment with radiation alone, suggesting that targeting telomerase could be a way to increase the efficacy of radiation in treating radiation-resistant tumors.
Heart disease, diabetes and quality of life
Blackburn also discovered that mothers caring for very sick children have shorter telomeres when they report that their emotional stress is at a maximum and that telomerase was active at the site of blockages in coronary artery tissue, possibly accelerating heart attacks.
In 2009, it was shown that the amount of telomerase activity significantly increased following psychological stress. Across the sample of patients telomerase activity in peripheral blood mononuclear cells increased by 18% one hour after the end of the stress.[65]
A study in 2010 found that there was "significantly greater" telomerase activity in participants than controls after a three-month meditation retreat.[66]
Telomerase deficiency has been linked to diabetes mellitus and impaired insulin secretion in mice, due to loss of pancreatic insulin-producing cells.[67]
Rare human diseases
Mutations in TERT have been implicated in predisposing patients to aplastic anemia, a disorder in which the bone marrow fails to produce blood cells, in 2005.[68]
Cri du chat syndrome (CdCS) is a complex disorder involving the loss of the distal portion of the short arm of chromosome 5. TERT is located in the deleted region, and loss of one copy of TERT has been suggested as a cause or contributing factor of this disease.[69]
Dyskeratosis congenita (DC) is a disease of the bone marrow that can be caused by some mutations in the telomerase subunits.[70] In the DC cases, about 35% cases are X-linked-recessive on the DKC1 locus[71] and 5% cases are autosomal dominant on the TERT[72] and TERC[73] loci.
Patients with DC have severe bone marrow failure manifesting as abnormal skin pigmentation, leucoplakia (a white thickening of the oral mucosa) and nail dystrophy, as well as a variety of other symptoms. Individuals with either TERC or DKC1 mutations have shorter telomeres and defective telomerase activity in vitro versus other individuals of the same age.[74]
In one family autosomal dominant DC was linked to a heterozygous TERT mutation.[75] These patients also exhibited an increased rate of telomere-shortening, and genetic anticipation (i.e., the DC phenotype worsened with each generation).
See also
- DNA repair
- Imetelstat
- TA-65
- Telomere
- Epitalon
Reference
- What are telomeres and telomerase?
- Pardue ML, DeBaryshe PG (2011). "Retrotransposons that maintain chromosome ends". PNAS. 108 (51): 20317–24. doi:10.1073/pnas.1100278108. PMC 3251079. PMID 21821789.
- Cano MI, Dungan JM, Agabian N, Blackburn EH (March 1999). "Telomerase in kinetoplastid parasitic protozoa". Proceedings of the National Academy of Sciences of the United States of America. 96 (7): 3616–21. Bibcode:1999PNAS...96.3616C. doi:10.1073/pnas.96.7.3616. PMC 22343. PMID 10097086.
- Olovnikov AM (September 1973). "A theory of marginotomy. The incomplete copying of template margin in enzymic synthesis of polynucleotides and biological significance of the phenomenon". Journal of Theoretical Biology. 41 (1): 181–90. Bibcode:1973JThBi..41..181O. doi:10.1016/0022-5193(73)90198-7. PMID 4754905.
- Greider CW, Blackburn EH (December 1985). "Identification of a specific telomere terminal transferase activity in Tetrahymena extracts". Cell. 43 (2 Pt 1): 405–13. doi:10.1016/0092-8674(85)90170-9. PMID 3907856.
- "The Nobel Prize in Physiology or Medicine 2009". The Nobel Foundation. 2009-10-05. Retrieved 2010-10-23.
- Feng J, Funk WD, Wang SS, Weinrich SL, Avilion AA, Chiu CP, Adams RR, Chang E, Allsopp RC, Yu J (September 1995). "The RNA component of human telomerase". Science. 269 (5228): 1236–41. Bibcode:1995Sci...269.1236F. doi:10.1126/science.7544491. PMID 7544491. S2CID 9440710.
- Kim NW, Piatyszek MA, Prowse KR, Harley CB, West MD, Ho PL, Coviello GM, Wright WE, Weinrich SL, Shay JW (December 1994). "Specific association of human telomerase activity with immortal cells and cancer". Science. 266 (5193): 2011–5. Bibcode:1994Sci...266.2011K. doi:10.1126/science.7605428. PMID 7605428. S2CID 11965342.
- Sauerwald A, Sandin S, Cristofari G, Scheres SH, Lingner J, Rhodes D (April 2013). "Structure of active dimeric human telomerase". Nature Structural & Molecular Biology. 20 (4): 454–60. doi:10.1038/nsmb.2530. PMC 3785136. PMID 23474713.
- Jiang J, Miracco EJ, Hong K, Eckert B, Chan H, Cash DD, Min B, Zhou ZH, Collins K, Feigon J (April 2013). "The architecture of Tetrahymena telomerase holoenzyme". Nature. 496 (7444): 187–92. Bibcode:2013Natur.496..187J. doi:10.1038/nature12062. PMC 3817743. PMID 23552895.
- Jiang J, Chan H, Cash DD, Miracco EJ, Ogorzalek Loo RR, Upton HE, Cascio D, O'Brien Johnson R, Collins K, Loo JA, Zhou ZH, Feigon J (October 2015). "Structure of Tetrahymena telomerase reveals previously unknown subunits, functions, and interactions". Science. 350 (6260): aab4070. doi:10.1126/science.aab4070. PMC 4687456. PMID 26472759.
- Nguyen TH, Tam J, Wu RA, Greber BJ, Toso D, Nogales E, Collins K (May 2018). "Cryo-EM structure of substrate-bound human telomerase holoenzyme". Nature. 557 (7704): 190–195. Bibcode:2018Natur.557..190N. doi:10.1038/s41586-018-0062-x. PMC 6223129. PMID 29695869.
- Cohen S, Graham M, Lovrecz G, Bache N, Robinson P, Reddel R (2007). "Protein composition of catalytically active human telomerase from immortal cells". Science. 315 (5820): 1850–3. Bibcode:2007Sci...315.1850C. doi:10.1126/science.1138596. PMID 17395830. S2CID 36658925.
- "HGNC database of human gene names - HUGO Gene Nomenclature Committee". genenames.org.
- HGNC - TERC
- HGNC - DKC1
- HGNC - TEP1
- NCBI - telomerase reverse transcriptase isoform 1
- Gillis AJ, Schuller AP, Skordalakes E. Structure of the Tribolium castaneum telomerase catalytic subunit TERT. Nature. 2008 Oct 2;455(7213):633-7
- Mitchell M, Gillis A, Futahashi M, Fujiwara H, Skordalakes E. Structural basis for telomerase catalytic subunit TERT binding to RNA template and telomeric DNA. Nat Struct Mol Biol. 2010 Apr;17(4):513-8
- NCBI - telomerase reverse transcriptase
- Gavory G, Farrow M, Balasubramanian S (October 2002). "Minimum length requirement of the alignment domain of human telomerase RNA to sustain catalytic activity in vitro". Nucleic Acids Res. 30 (20): 4470–80. doi:10.1093/nar/gkf575. PMC 137139. PMID 12384594.
- Hayflick L, Moorhead PS (1961). "The serial cultivation of human diploid cell strains". Exp Cell Res. 25 (3): 585–621. doi:10.1016/0014-4827(61)90192-6. PMID 13905658.
- Siegel, L (2013). Are Telomeres the Key to Aging and Cancer? Archived 2013-12-01 at the Wayback Machine The University of Utah. Retrieved 30 September 2013
- Hanahan D, Weinberg RA (March 2011). "Hallmarks of cancer: the next generation". Cell. 144 (5): 646–74. doi:10.1016/j.cell.2011.02.013. PMID 21376230.
- Fice HE, Robaire B (July 2019). "Telomere Dynamics Throughout Spermatogenesis". Genes. 10 (7): 525. doi:10.3390/genes10070525. PMC 6678359. PMID 31336906.
- Härle-Bachor C, Boukamp P (June 1996). "Telomerase activity in the regenerative basal layer of the epidermis inhuman skin and in immortal and carcinoma-derived skin keratinocytes". Proceedings of the National Academy of Sciences of the United States of America. 93 (13): 6476–6481. Bibcode:1996PNAS...93.6476H. doi:10.1073/pnas.93.13.6476. PMC 39048. PMID 8692840.
- Barsov EV (March 2011). "Telomerase and primary T cells: biology and immortalization for adoptive immunotherapy". Immunotherapy. 3 (3): 407–421. doi:10.2217/imt.10.107. PMC 3120014. PMID 21395382.
- Bougel S, Renaud S, Braunschweig R, Loukinov D, Morse HC, Bosman FT, et al. (January 2010). "PAX5 activates the transcription of the human telomerase reverse transcriptase gene in B cells". The Journal of Pathology. 220 (1): 87–96. doi:10.1002/path.2620. PMC 3422366. PMID 19806612.
- Cong YS, Wright WE, Shay JW (September 2002). "Human telomerase and its regulation". Microbiology and Molecular Biology Reviews. 66 (3): 407–25, table of contents. doi:10.1128/MMBR.66.3.407-425.2002. PMC 120798. PMID 12208997.
- Gomes NM, Ryder OA, Houck ML, Charter SJ, Walker W, Forsyth NR, Austad SN, Venditti C, Pagel M, Shay JW, Wright WE (October 2011). "Comparative biology of mammalian telomeres: hypotheses on ancestral states and the roles of telomeres in longevity determination". Aging Cell. 10 (5): 761–8. doi:10.1111/j.1474-9726.2011.00718.x. PMC 3387546. PMID 21518243.
- Cherif H, Tarry JL, Ozanne SE, Hales CN (March 2003). "Ageing and telomeres: a study into organ- and gender-specific telomere shortening". Nucleic Acids Research. 31 (5): 1576–83. doi:10.1093/nar/gkg208. PMC 149817. PMID 12595567.
- Renault V, Thornell LE, Eriksson PO, Butler-Browne G, Mouly V, Thorne LE (December 2002). "Regenerative potential of human skeletal muscle during aging". Aging Cell. 1 (2): 132–9. doi:10.1046/j.1474-9728.2002.00017.x. PMID 12882343.
- Jeyapalan JC, Ferreira M, Sedivy JM, Herbig U (January 2007). "Accumulation of senescent cells in mitotic tissue of aging primates". Mechanisms of Ageing and Development. 128 (1): 36–44. doi:10.1016/j.mad.2006.11.008. PMC 3654105. PMID 17116315.
- Verma S, Tachtatzis P, Penrhyn-Lowe S, Scarpini C, Jurk D, Von Zglinicki T, Coleman N, Alexander GJ (October 2012). "Sustained telomere length in hepatocytes and cholangiocytes with increasing age in normal liver". Hepatology. 56 (4): 1510–20. doi:10.1002/hep.25787. PMID 22504828. S2CID 25965027.
- Harris SE, Martin-Ruiz C, von Zglinicki T, Starr JM, Deary IJ (July 2012). "Telomere length and aging biomarkers in 70-year-olds: the Lothian Birth Cohort 1936". Neurobiology of Aging. 33 (7): 1486.e3–8. doi:10.1016/j.neurobiolaging.2010.11.013. PMID 21194798. S2CID 10309423.
- de Magalhães JP, Toussaint O (2004). "Telomeres and telomerase: a modern fountain of youth?". Rejuvenation Res. 7 (2): 126–33. CiteSeerX 10.1.1.318.8027. doi:10.1089/1549168041553044. PMID 15312299.
- Tomás-Loba A, Flores I, Fernández-Marcos PJ, Cayuela ML, Maraver A, Tejera A, Borrás C, Matheu A, Klatt P, Flores JM, Viña J, Serrano M, Blasco MA (November 2008). "Telomerase reverse transcriptase delays aging in cancer-resistant mice". Cell. 135 (4): 609–22. doi:10.1016/j.cell.2008.09.034. PMID 19013273. S2CID 14753825.
- Atzmon G, Cho M, Cawthon RM, Budagov T, Katz M, Yang X, Siegel G, Bergman A, Huffman DM, Schechter CB, Wright WE, Shay JW, Barzilai N, Govindaraju DR, Suh Y (January 2010). "Genetic variation in human telomerase is associated with telomere length in Ashkenazi centenarians". Proc. Natl. Acad. Sci. U.S.A. 107 (Suppl 1): 1710–7. doi:10.1073/pnas.0906191106. PMC 2868292. PMID 19915151.
- Lay summary in: "One Key Found for Living to 100". LiveScience. November 12, 2009.
- Blasco MA (August 2005). "Telomeres and human disease: ageing, cancer and beyond". Nature Reviews. Genetics. 6 (8): 611–22. doi:10.1038/nrg1656. PMID 16136653. S2CID 14828121.
- Akincilar SC, Unal B, Tergaonkar V (April 2016). "Reactivation of telomerase in cancer". Cellular and Molecular Life Sciences. 73 (8): 1659–1670. doi:10.1007/s00018-016-2146-9. PMC 4805692. PMID 26846696.
- Skloot R (2010). The Immortal Life of Henrietta Lacks. New York: Broadway Paperbacks. pp. 216, 217. ISBN 978-1-4000-5218-9.
- Dr. Todd Hennessey, 2016 University at Buffalo
- Grech G, Baldacchino S, Saliba C, Grixti MP, Gauci R, Petroni V, et al. (September 2016). "Deregulation of the protein phosphatase 2A, PP2A in cancer: complexity and therapeutic options". Tumour Biology. 37 (9): 11691–11700. doi:10.1007/s13277-016-5145-4. PMID 27444275. S2CID 24784814.
- Shay JW, Bacchetti S (April 1997). "A survey of telomerase activity in human cancer". European Journal of Cancer. 33 (5): 787–91. doi:10.1016/S0959-8049(97)00062-2. PMID 9282118.
- Bryan TM, Englezou A, Gupta J, Bacchetti S, Reddel RR (September 1995). "Telomere elongation in immortal human cells without detectable telomerase activity". The EMBO Journal. 14 (17): 4240–8. doi:10.1002/j.1460-2075.1995.tb00098.x. PMC 394507. PMID 7556065.
- Henson JD, Neumann AA, Yeager TR, Reddel RR (January 2002). "Alternative lengthening of telomeres in mammalian cells". Oncogene. 21 (4): 598–610. doi:10.1038/sj.onc.1205058. PMID 11850785.
- Blackburn EH (February 2005). "Telomeres and telomerase: their mechanisms of action and the effects of altering their functions". FEBS Letters. 579 (4): 859–862. doi:10.1016/j.febslet.2004.11.036. PMID 15680963.
- Tian X, Chen B, Liu X (March 2010). "Telomere and telomerase as targets for cancer therapy". Applied Biochemistry and Biotechnology. 160 (5): 1460–72. doi:10.1007/s12010-009-8633-9. PMID 19412578. S2CID 20560225.
- Griffiths AJ, Wesslet SR, Carroll SB, Doebley J (2008). Introduction to Genetic Analysis. W. H. Freeman. ISBN 978-0-7167-6887-6.
- Williams SC (January 2013). "No end in sight for telomerase-targeted cancer drugs". Nature Medicine. 19 (1): 6. doi:10.1038/nm0113-6. PMID 23295993. S2CID 12232531.
- Blasco MA (2001). "Telomeres in Cancer Therapy". Journal of Biomedicine & Biotechnology. 1 (1): 3–4. doi:10.1155/S1110724301000109. PMC 79678. PMID 12488618.
- Shay JW, Zou Y, Hiyama E, Wright WE (April 2001). "Telomerase and cancer". Human Molecular Genetics. 10 (7): 677–85. doi:10.1093/hmg/10.7.677. PMID 11257099.
- Gül I, Dündar O, Bodur S, Tunca Y, Tütüncü L (September 2013). "The status of telomerase enzyme activity in benign and malignant gynaecologic pathologies". Balkan Medical Journal. 30 (3): 287–92. doi:10.5152/balkanmedj.2013.7328. PMC 4115914. PMID 25207121.
- Saretzki G (May 2003). "Telomerase inhibition as cancer therapy". Cancer Letters. 194 (2): 209–19. doi:10.1016/s0304-3835(02)00708-5. PMID 12757979.
- Stoyanov V (2009). "T-loop deletion factor showing speeding aging of Homo telomere diversity and evolution". Rejuvenation Research. 12 (1): 52.
- Fernandez ML, Thomas MS, Lemos BS, DiMarco DM, Missimer A, Melough M, et al. (2018). "TA-65, A Telomerase Activator improves Cardiovascular Markers in Patients with Metabolic Syndrome". Current Pharmaceutical Design. 24 (17): 1905–1911. doi:10.2174/1381612824666180316114832. PMID 29546832. S2CID 3892746.
- Harley CB, Liu W, Blasco M, Vera E, Andrews WH, Briggs LA, Raffaele JM (February 2011). "A natural product telomerase activator as part of a health maintenance program". Rejuvenation Research. 14 (1): 45–56. doi:10.1089/rej.2010.1085. PMC 3045570. PMID 20822369.
- Salvador L, Singaravelu G, Harley CB, Flom P, Suram A, Raffaele JM (December 2016). "A Natural Product Telomerase Activator Lengthens Telomeres in Humans: A Randomized, Double Blind, and Placebo Controlled Study". Rejuvenation Research. 19 (6): 478–484. doi:10.1089/rej.2015.1793. PMC 5178008. PMID 26950204.
- Harley CB, Liu W, Flom PL, Raffaele JM (October 2013). "A natural product telomerase activator as part of a health maintenance program: metabolic and cardiovascular response". Rejuvenation Research. 16 (5): 386–395. doi:10.1089/rej.2013.1430. PMID 23808324.
- Hoffmann J, Richardson G, Haendeler J, Altschmied J, Andrés V, Spyridopoulos I (March 2021). "Telomerase as a Therapeutic Target in Cardiovascular Disease". Arteriosclerosis, Thrombosis, and Vascular Biology. 41 (3): 1047–1061. doi:10.1161/ATVBAHA.120.315695. PMID 33504179. S2CID 231753311.
- Patel KP, Vonderheide RH (June 2004). "Telomerase as a tumor-associated antigen for cancer immunotherapy". Cytotechnology. 45 (1–2): 91–9. doi:10.1007/s10616-004-5132-2. PMC 3449959. PMID 19003246.
- Johnson SR (2 September 2015). "Experimental blood disorder therapy shows promise in new studies". Modern Healthcare.
- Chen M, Xing LN (2012). "siRNA-mediated inhibition of hTERC enhances radiosensitivity of cervical cancer". Asian Pacific Journal of Cancer Prevention. 13 (12): 5975–9. doi:10.7314/apjcp.2012.13.12.5975. PMID 23464388.
- Epel ES, Lin J, Dhabhar FS, Wolkowitz OM, Puterman E, Karan L, Blackburn EH (May 2010). "Dynamics of telomerase activity in response to acute psychological stress". Brain, Behavior, and Immunity. 24 (4): 531–9. doi:10.1016/j.bbi.2009.11.018. PMC 2856774. PMID 20018236.
- Jacobs TL, Epel ES, Lin J, Blackburn EH, Wolkowitz OM, Bridwell DA, Zanesco AP, Aichele SR, Sahdra BK, MacLean KA, King BG, Shaver PR, Rosenberg EL, Ferrer E, Wallace BA, Saron CD (June 2011). "Intensive meditation training, immune cell telomerase activity, and psychological mediators". Psychoneuroendocrinology. 36 (5): 664–81. doi:10.1016/j.psyneuen.2010.09.010. PMID 21035949. S2CID 4890811.
- Kuhlow D, Florian S, von Figura G, Weimer S, Schulz N, Petzke KJ, Zarse K, Pfeiffer AF, Rudolph KL, Ristow M (October 2010). "Telomerase deficiency impairs glucose metabolism and insulin secretion". Aging. 2 (10): 650–8. doi:10.18632/aging.100200. PMC 2993795. PMID 20876939.
- Yamaguchi H, Calado RT, Ly H, Kajigaya S, Baerlocher GM, Chanock SJ, Lansdorp PM, Young NS (April 2005). "Mutations in TERT, the gene for telomerase reverse transcriptase, in aplastic anemia". The New England Journal of Medicine. 352 (14): 1413–24. doi:10.1056/NEJMoa042980. PMID 15814878.
- Zhang A, Zheng C, Hou M, Lindvall C, Li KJ, Erlandsson F, Björkholm M, Gruber A, Blennow E, Xu D (April 2003). "Deletion of the telomerase reverse transcriptase gene and haploinsufficiency of telomere maintenance in Cri du chat syndrome". American Journal of Human Genetics. 72 (4): 940–8. doi:10.1086/374565. PMC 1180356. PMID 12629597.
- Yamaguchi H (June 2007). "Mutations of telomerase complex genes linked to bone marrow failures". Journal of Nippon Medical School. 74 (3): 202–9. doi:10.1272/jnms.74.202. PMID 17625368.
- Heiss NS, Knight SW, Vulliamy TJ, Klauck SM, Wiemann S, Mason PJ, Poustka A, Dokal I (May 1998). "X-linked dyskeratosis congenita is caused by mutations in a highly conserved gene with putative nucleolar functions". Nature Genetics. 19 (1): 32–8. doi:10.1038/ng0598-32. PMID 9590285. S2CID 205342127.
- Vulliamy TJ, Walne A, Baskaradas A, Mason PJ, Marrone A, Dokal I (2005). "Mutations in the reverse transcriptase component of telomerase (TERT) in patients with bone marrow failure". Blood Cells, Molecules & Diseases. 34 (3): 257–63. doi:10.1016/j.bcmd.2004.12.008. PMID 15885610.
- Vulliamy T, Marrone A, Goldman F, Dearlove A, Bessler M, Mason PJ, Dokal I (September 2001). "The RNA component of telomerase is mutated in autosomal dominant dyskeratosis congenita". Nature. 413 (6854): 432–5. Bibcode:2001Natur.413..432V. doi:10.1038/35096585. PMID 11574891. S2CID 4348062.
- Marrone A, Walne A, Dokal I (June 2005). "Dyskeratosis congenita: telomerase, telomeres and anticipation". Current Opinion in Genetics & Development. 15 (3): 249–57. doi:10.1016/j.gde.2005.04.004. PMID 15917199.
- Armanios M, Chen JL, Chang YP, Brodsky RA, Hawkins A, Griffin CA, Eshleman JR, Cohen AR, Chakravarti A, Hamosh A, Greider CW (November 2005). "Haploinsufficiency of telomerase reverse transcriptase leads to anticipation in autosomal dominant dyskeratosis congenita". Proceedings of the National Academy of Sciences of the United States of America. 102 (44): 15960–4. Bibcode:2005PNAS..10215960A. doi:10.1073/pnas.0508124102. PMC 1276104. PMID 16247010.
Further reading
- The Immortal Cell, by Michael D. West, Doubleday (2003) ISBN 978-0-385-50928-2
External links
- Gene Ontology: GO:0003720: telomerase activity
- Human telomerase reverse transcriptase (TERT) gene on genecards.org
- The Telomerase Database - A Web-based tool for telomerase research
- Three-dimensional model of telomerase at MUN
- Elizabeth Blackburn's Seminars: Telomeres and Telomerase
- Telomerase at the US National Library of Medicine Medical Subject Headings (MeSH)
- Overview of all the structural information available in the PDB for UniProt: O14746 (Human Telomerase reverse transcriptase) at the PDBe-KB.
- Overview of all the structural information available in the PDB for UniProt: Q0QHL8 (Tribolium castaneum Telomerase reverse transcriptase) at the PDBe-KB.
{kind=link}