Gen blanco dominante del caballo
El blanco dominante es un grupo de condiciones genéticas en el color del pelo del caballo, conocidas principalmente por producir una capa completamente blanca, pero también por algunos tipos de manchas y marcas de ese mismo color. Los caballos blancos dominantes nacen con la piel de color rosado, sin pigmentación, y pelo blanco con ojos oscuros, sin embargo esto puede variar dependiendo de la mutación genética que esté involucrada. El blanco dominante es una mutación poco frecuente y, en condiciones normales, al menos uno de los padres debe ser de color blanco dominante para producir descendencia de este tipo. Sin embargo, la mayoría de los Alelos actualmente conocidos del blanco dominante pueden estar vinculados a una mutación espontánea documentada en un único ancestro.[¿quién?]
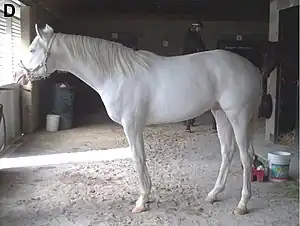
La mutación blanco dominante puede suceder en cualquier raza, y ha sido estudiado en muchas razas diferentes, de hecho, dos razas de color, el caballo blanco americano y el caballo blanco camarillo se caracterizan por sus capas blancas.
Todos los colores conocidos de la capa blanco dominante están asociados al gen KIT. Hay varios alelos del gen KIT que dan como resultado el blanco dominante; en genética, desde 2013, están etiquetados del W1 hasta el W20. Como el nombre lo sugiere, los alelos que dan lugar a estas capas blancas conocidas son de herencia dominante, por lo que un caballo necesita solo una copia del alelo W para tener una capa blanca o una capa con manchas blancas.
El blanco dominante es genéticamente distinto al sabino y ambos, genética y visualmente, distintos del tordo y cremello. El blanco dominante no es el mismo que el factor letal blanco dominante ni el de los caballos blancos "albinos". El albinismo nunca se ha documentado en los caballos[cita requerida]. Se cree que algunas formas de blanco dominante para dar lugar a embriones no viables es cuando un cigoto es homocigoto, es decir los dos alelos son W, sin embargo, esto no ha sido verificado para todas las variaciones genéticas de blancos dominantes.
Identificación

Los caballos blancos dominantes nacen con la piel rosada y una capa blanca que conservan toda su vida.[1] A pesar de que el término "blanco dominante" se asocia típicamente con la capa de color blanco puro estos caballos pueden ser todos blancos, casi blancos, parcialmente blancos o exhibir un patrón de manchado irregular similar a la de caballos sabino.[2] La cantidad de pelo blanco depende de qué alelo mutado de W esté involucrado.[3] Las áreas no blancas de la piel y el cabello son más frecuentes a lo largo de la línea media dorsal del caballo, conocida como la línea escapulo espinal, y son especialmente comunes en la crin y en las orejas.[2] También pueden tener motas intercaladas o manchas en la piel y pelo no blancas. Además, los cascos son, más a menudo, blancos, pero pueden tener rayas si hay piel pigmentada en la banda coronaria, justo por encima de la pezuña.[4][5] En algunos casos, los potros nacidos con pelo no blanco residual puede perder parte o todo este pigmento con la edad y sin ayuda del factor tordo.[6] El moteado blanco dominante no afecta el color de ojos, los caballos blancos dominantes tienen ojos marrones.[7] La piel de color rosado está desprovista de melanocitos, y aparece de color rosado de la red subyacente de los capilares. El pelo blanco tiene sus raíces en la piel de color rosado sin pigmentación. Hay muchos otros factores genéticos que producen blanco, casi blanco, y los colores del pelaje de color blanco apagado en caballos, algunos de los cuales son visualmente muy similares al blanco dominante.[8]
Prevalencia
El blanco dominante es una de las diversas causas genéticas potenciales para los caballos con capas casi o completamente blancas, y puede ocurrir a través de mutación espontánea, por lo que puede ser encontrado inesperadamente en cualquier raza, incluso aquellos que desalientan marcas blancas excesivas. Hasta la fecha, el blanco dominante ha sido identificado en varias familias de purasangre,[6] Caballos americanos Cuarto de Milla,[2] caballos frederiksborg,[2] caballos islandeses,[2] Poni de las Shetland,[6] caballos franches montagnes,[6] caballos de tiro del sur de Alemania,[2] y en una familia de caballos árabes.[6] El caballo blanco americano, que desciende principalmente de un caballo blanco dominante cruzado con yeguas que no son blancas, es conocido por su pelaje blanco dominante, como es el caballo blanco de camarillo.[9][10]
Herencia
El locus W fue mapeado en el gen KIT en el año 2007.[6] Los términos "oncogen kit" y gen "manchado dominante", simbolizados por KIT y W, respectivamente, se pueden utilizar indistintamente.[11] La investigación actual ha demostrado que existen múltiples formas, o alelos, del gen W.[6] Todos los caballos poseen el gen KIT ya que es necesario para la supervivencia desde las primeras etapas del desarrollo. La presencia o ausencia del blanco dominante se basa en la presencia de ciertas formas alteradas del gen KIT. Cada forma única se llama alelo, y para cada rasgo, todos los animales heredan un alelo de cada progenitor. La forma original o "normal" del gen KIT que se espera en los caballos sin manchas de blanco dominante, es llamado alelo "tipo salvaje", por lo tanto, un caballo blanco dominante tiene un alelo de KIT con una mutación asociada con manchas blancas dominantes, y un alelo de KIT de tipo silvestre.
Historia de la investigación del blanco dominante
Los caballos blancos dominantes fueron descritos por primera vez en la literatura científica en 1912. El criador de caballos William P. Newell describió su familia de caballos blancos y casi blancos al investigador A. P. Sturtevant de la Universidad de Columbia:


" El color de la piel es de color blanco o llamado rosa, por lo general con un par de pequeñas manchas oscuras en la piel. Algunos tienen un gran número de manchas oscuras en la piel. Estos últimos suelen tener unas pocas rayas oscuras en los cascos; de lo contrario, los cascos son casi invariablemente blancos. Aquellos que no tienen manchas oscuras en la piel, por lo general tienen vidrio u ojos que ven; de lo contrario, ojos oscuros [...] Tengo un potro de un año de edad que es de color blanco puro, sin una mota ni cabello de color en él, y con ojos [azules] de vidrio"[5]
Sturtevant y sus contemporáneos coincidieron en que los ojos azules de este potro fueron heredados por separado de la capa blanca.[12] En 1912, Sturtevant asignó el carácter "blanco" al locus blanco o W.[5] En ese momento no había medios para asignar W a una posición en el cromosoma o a un gen.
Esta familia de caballos blancos produjo a Old King en 1908, un semental blanco de ojos oscuros que fue comprado por Caleb R. y Hudson B. Thompson. Old King fue cruzado con yeguas morgan para producir una raza de caballo conocida hoy como caballo blanco americano.[9] Un nieto de Old King, Snow King, estaba en el centro del primer estudio importante de la capa de color blanco dominante en caballos, realizado en 1969 por el Dr. William L. Pulos de la Universidad de Alfred y el Dr. Frederick B. Hutt de Cornell. Concluyeron, basándose en los apareamientos de prueba y en las relaciones de fenotipo de progenie que la capa blanca fue heredada predominantemente, y que poduce embriones letales en homocigosis.[13] Otros factores, tales como las variaciones en la expresividad y la influencia de múltiples genes, pudo haber influido en las proporciones de la progenie que Pulos y Hutt observaron.[14] La capa blanca del caballo blanco americano aun no ha sido mapeada.
En un estudio de 1924 realizado por C. Wriedt se identificó un color blanco hereditario en el caballo de frederiksborg.[15] Wriedt describió una gama de lo que consideró fenotipos homocigotos: todo blanco, blanco con manchas pigmentadas, o weißgraue, que se traduce como "blanco-gris"[16] El término alemán para el caballo tordo es schimmel, no weißgraue.[17] Los heterocigotos, según Wriedt, oscilaron entre ruano o diluido con caballos blancos más o menos sólidos. Los revisores, reinterpretaron los datos de Wriedt en años sucesivos, mientras que Pulos y Hutt consideraban que su trabajo había sido "erróneo" porque Wriedt nunca llegó a la conclusión de que el factor blanco era letal cuando estaba en homocigosis.[18]
Otros investigadores, antes del análisis de ADN moderno, desarrollaron teorías notablemente premonitorias. El gen, en sí mismo, fue propuesto por primera vez y nombrado como W en 1948.[3] En un trabajo de 1969 sobre los colores del pelaje del caballo, A los colores del caballo, Miguel Odriozola sugirió que las diversas formas de manchas blancas de herencia dominante podrían estar dispuestas secuencialmente a lo largo de un cromosoma, por lo tanto permite la expresión variable del blanco dominante. También propuso que otros genes distantes podrían influir en la cantidad de blanco prente.[19]

Durante el estudio Pulos y de Hutt en 1969 y el inicio de la investigación a nivel molecular en el blanco dominante en el siglo XXI un modelo, conocido como "sabino", llegó a ser considerado por algunos como una de las causas más probables de los fenotipos de color blanco.[20] El sabino es un tipo de caballo de manchas blancas y el único alelo por ahora mapeado; aunque es distinto, la herencia dominante Sabino-1 (SB-1) está relacionada genéticamente. Cuando son homocigotos, pueden producir SB-1 casi todos los caballos blancos que se asemejen al blanco dominante. Otros genes responsables de todos los patrones posibles etiquetados como "sabino" aun no han sido identificados, aunque algunas formas del gen blanco salpicado pueden ser responsables de ciertos patrones.
En el año 2007 investigadores de Suiza y Estados Unidos publicaron un documento que identificaba la causa genética del moteado blanco dominante en caballos franches montagnes, caballo blanco camarillo, caballo árabe y razas de purasangre.[6] Cada una de estas mutaciones de blancos dominantes se habían producido por separado y de forma espontánea en los últimos 75 años y cada una representa un alelo diferente (variación o forma) de un mismo gen. Estos mismos investigadores identificaron otras siete causas únicas de blanco dominante en el año 2009: tres en familias distintas de purasangre, en un caballo islandés, en un holstein, una gran familia de caballos Cuarto de Milla y una familia de caballos de tiro de Alemania del Sur.[2]
Series alélicas
El gen KIT contiene más de 2000 pares de bases, un cambio en cualquiera de ellos, resulta en un alelo mutado.[2] Se han identificado, mediante la secuenciación de los genes KIT de varios caballos, más de cuarenta alelos de ese tipo.[2] El fenotipo resultante de la mayoría de estos alelos no se conoce todavía, pero veinte se han relacionado con blanco dominante.[3] Hasta la fecha las pruebas de ADN pueden identificar si un caballo lleva los diversos alelos de W identificados, algunas disponibles comercialmente.[21]
- W1 se encontró en caballos freiberger descendientes de una yegua blanca llamada Cigale nacida en 1957. Las capas de los padres de Cigale no estaban ampliamente marcados.[6] Un polimorfismo de un solo nucleótido (SNP), se cree que se produjo con Cigale un tipo de mutación en la cual un solo nucleótido es, accidentalmente, intercambiado por otro. Se cree que esta mutación (c.2151C> G) afecta gravemente la función del gen KIT.[6] Se trata de una mutación sin sentido encontrada en el Exon15 de KIT.[3] Algunos caballos con la mutación W1 nacen de color blanco puro, pero muchos tienen pigmento residual a lo largo de la línea escapulo espinal, que luego pueden perder con el tiempo. Basándose en estudios de mutaciones de KIT en ratones, la gravedad de esta mutación sugiere que puede no ser viable en homocigosis.[22] Sin embargo, en los caballos con la mutación W1 se ha encontrado que tienen parámetros normales de sangre y no sufren de anemia.[23]
- W2 se encontró en caballos de purasangre descendientes de KY Coronel, un semental nacido en 1946. Este fue descrito como un castaño con extensas manchas blancas, y conocido por engendrar una familia de caballos blancos puros a través de su hija blanca, "White Beauty", nacida en 1963.[24] El alelo W2 también está relacionado con un polimorfismo de un solo nucleótido (c.1960G>A),[6] pero es una mutación con cambio de sentido situado en el Exón17.[3]
- W3 se encontró en los caballos árabes descendientes de R. Khasper, un semental casi blanco nacido en 1996. Ninguno de sus padres eran blancos y la mutación causante (c.706A>T) es la que se cree que se originó con este caballo.[6] Es una mutación con cambio de sentido en el Exón4.[3] Los caballos con el alelo W3 a menudo conservan manchas intercaladas o regiones de la piel pigmentadas que pueden desaparecer con el tiempo.[24] Algunos miembros de esta familia tienen los ojos azules, pero se piensa que éstos se heredan por separado de la capa blanca.[24] Basado en estudios similares en ratones, los investigadores han nombrado a W3 como potencialmente homocigótico inviable.[22]

- W4 se encontró en caballos blancos Camarillo, una raza que se caracteriza por su capa blanca, que comienza con un semental blanco espontáneo nacido en 1912 llamado Sultán.[6] Al igual que W1 y W3, los caballos con este tipo del blanco dominante puede ser de color blanco puro o casi blanco, con zonas pigmentadas a lo largo de la línea superior que se desvanece con el tiempo.[24] Esta mutación es también un SNP (c.1805C> T),[6] una mutación con cambio de sentido en el Exón12.[3]
- W5 se encontró en los purasangre que descienden de Puchilingui,[21] un semental de 1984 con manchas blancos, sabino y ruano.[2] Los caballos con el alelo W5 exhiben una enorme variedad en el fenotipo blanco: unos pocos han sido de color blanco puro o casi blanco, mientras que otros tienen manchas sabino limitado a medias altas, irregulares y llamas que cubrían la cara. Se estudiaron veintidós miembros de esta familia, y doce con cierto grado de moteado blanco dominante, se encontró que tenían una deleción en el exón 15 (p.T732QfsX9)[2] en la forma de una mutación con desplazamiento del marco de lectura.[3] Un estudio posterior encontró que los miembros de esta familia con mayor despigmentación eran heterocigotos compuestos que también portaban el alelo W20.[25]
- W6 se encontró en un purasangre casi de color blanco nacido nacido de padres no blancos en 2004. La potencial gama de expresividad no se conoce todavía. Se cree la mutación (c.856G> A) se ha producido de forma espontánea, en este caballo,[2] una mutación con cambio de sentido en el Exón5.[3]
- W7 se encontró en una yegua de purasangre casi blanca nacida en 2005, tuvo nueve descendientes, todos no-blanco. No tenía el alelo W7, que resulta de una mutación del sitio de empalme (c.338-1G> C),[2] localizada en el Intrón2 de KIT.[3]
- W8 se encontró en un caballo islandés con manchas sabino blancas, moteado, y runeado. Los padres y los cuatro medios hermanos maternos eran todos no-blancos, se encontraron sin el alelo W8. El alelo W8 es también una mutación del sitio de empalme (c.2222-1G> A),[2] localizada en el Intrón15.[3]
- W9 se encontró en un caballo holsteiner totalmente blanco con un polimorfismo de un solo nucleótido (c.1789G> A). No se estudió a ningún pariente, pero ambos padres eran no blancos.[2] Es una mutación con cambio de sentido en el Exón12.[3]
- W10 se encontró en un estudio en veintisiete caballos en una familia de Caballos Cuarto de Milla, diez de los cuales son de color blanco o manchado y diecisiete eran sólidos y no blancos. Los diez miembros de la familia con W10 tenían una deleción en el exón 7 (c.1126_1129delGAAC). Como en el W5, se observó una amplia gama de fenotipos. El más marcado tenía grandes cantidades de color blanco en la cara y las patas, y algunos puntos de tamaño medio en el vientre, mientras que lo demás era casi totalmente blanco.[2] Se trata de una mutación de marco en el Exón7.[3] El fundador de esta línea era GQ Santana que nació en el año 2000.[21]
- W11 se encontró en una familia de caballos de tiro en Alemania del Sur que descienden de un solo caballo blanco, en el que se cree que se originó la mutación casusal. La mutación responsable del fenotipo W11 es una mutación del sitio de empalme del intrón 20 (c.2684+1G>A).[2]
- W12 se encontró en los caballos purasangre, y es una mutación de deleción que se encuentra en el Exón3.[3]
- W13 se encontró en los caballos Cuarto de Milla y es un una mutación del sitio de empalme en el Intrón17.[3]
- W14 es una mutación de deleción en el Exón17, se encontró en purasangres.[3]
- W15 se encontró en los caballos árabes y es una mutación de cambio de sentido sentido en el Exón10.[3]
- W16 se encontró en el Oldenburger y es una mutación sin sentido en el Exón18.[3]
- W17 se encontró en un caballo de tiro japonés y es una mutación de cambio de sentido sentido en el Exón14.[3]
- W18 es una mutación del sitio de empalme en el Intrón8 (c.1346 +1G>A) se encontró en la bahía suiza "Warmblood" con un amplio moteado. Ambos padres eran de color sólido y no tenía marcas extendidas en la cabeza o las piernas.[25]
- W19 se encontró en tres caballos árabes con marcas calvas en la cara, otras blancas desde las patas hasta las rodillas y los corvejones, y manchas irregulares en el vientre. Los tres caballos dieron negativo para sabino-1, marco overo y blanco salpicado. W19 es una mutación sin sentido en el exón 8 (c.1322A.G; p.Tur41Cys). Este gen se prevé que sea "probablemente perjudicial" y uno de los tres caballos tiene la descendencia de un solo color que no portan el alelo.[25]
- W20 es una mutación sin sentido en el Exón14 (c.2045G> A; p.Arg682His) descubierto originalmente en 2007, pero no reconocido por tener un papel sutil en el aumento de las marcas blancas y la pigmentación blanca. Aparece en muchas razas, pero sus efectos fueron reconocidos por primera vez en la familia W5 de purasangre y se determinó que era el factor causante de los caballos ("blancos") más ampliamente-despigmentados.[25]
Estos alelos no tienen en cuenta todas las manchas blancas de herencia dominante en los caballos. Se espera que más alelos KIT se encuentren con papeles en el manchado blanco.[2] La mayoría de los alelos de W surgen dentro de una raza o una familia específica y son mutaciones espontáneas. El propio gen KIT parece propenso a la mutación, y así podrían surgir virtualmente nuevos alelos de W podrían ocurrir en cualquier raza.[3]
Genética molecular
El gen KIT codifica una proteína llamada "receptor del factor de acero", que es crítica para la diferenciación de las células madre en células sanguíneas, espermatozoides, y células pigmentantes. Un proceso llamado empalme alternativo que utiliza la información codificada en el gen KIT para producir proteínas (isoformas) ligeramente diferentes para su uso en distintas circunstancias, puede afectar si una mutación en KIT afecta a las células sangíneas, células, los espematzoides o las células de pigmentarias. El receptor del factor acero interactúa químicamente con el factor acero o factor de células madre para retransmitir mensajes químicos. Estos mensajes se utilizan durante el desarrollo embrionario para señalar la migración de los melanocitos (células tempranas de pigmento) desde el tejido de la cresta neural a sus destinos finales en la capa dérmica. La cresta neural es un tejido transitorio en el embrión que se encuentra a lo largo de la línea dorsal. Los melanocitos migran a lo largo de la línea dorsal a una serie de sitios específicos: cerca del ojo, cerca de la oreja, y la parte superior de la cabeza; seis sitios a lo largo de cada lado del cuerpo, y unos pocos a lo largo de la cola. En estos sitios, las células se someten a unas cuantas rondas de replicación y diferenciación, y luego migran hacia abajo y alrededor del cuerpo desde la cara dorsal hacia la cara ventral y las yemas de las extremidades.[26]
El momento de esta migración es crítica. Todas las marcas blancas, desde de una pequeña estrella hasta una capa de color blanco puro, son causados por la migración fallida de los melanocitos.[27]
Un cierto grado de la cantidad eventual de blanco, y su diseño, es completamente al azar. El desarrollo de un organismo de una sola célula a completamente formado es un proceso con muchos pasos. Incluso a partir de genomas idénticos como clones o gemelos idénticos, el proceso es poco probable que suceda de la misma forma dos veces. Este proceso con el elemento de aleatoriedad se llama proceso estocástico y la diferenciación celular es, en parte, un proceso estocástico.[28] El elemento estocástico de desarrollo es, por un lado, responsable de la aparición eventual de blanco sobre un caballo, lo que potencialmente representa casi una cuarta parte del fenotipo.[29] El equipo de investigación que estudió el blanco dominante citó que "sutiles variaciones en la cantidad de proteína KIT residual" son una posible causa de la variabilidad en el fenotipo de los caballos con el mismo tipo de blanco dominante.[2]
Letalidad
La letalidad embrionaria temprana, también conocida como muerte embrionaria temprana o embrión no viable, se puede producir cuando el embrión posee dos alelos blanco dominante (W/+), o es homocigótico (W/W).[30] La razón de esto es que muchas mutaciones para W son causadas por mutaciones sin sentido, mutaciones de cambio o deleciones de ADN, las cuales, si están en homocigocis, harían imposible la producción de una proteína KIT funcional. Sin embargo, es posible que los embriones homocigóticos de los alelos de sentido erróneo y mutaciones de un sitio de empalme podrían ser viables debido a que tienen menos efecto sobre la función de genes.[3] Un estudio del 2013 también descubrió caballos que estaban compuestos de heterocigotos W5/W20, casi completamente blancos, esencialmente con mayor despigmentación de lo que podría explicarse por cualquier alelo.[25]
La hipótesis de la letalidad embrionaria fue apoyada inicialmente por el estudio de Pulos y de Hutt de 1969 de los coeficientes de la progenie mendeliana.[1] Las conclusiones acerca de los rasgos mendelianos que son controlados por un solo gen, pueden extraerse de crías de prueba con muestreos de gran tamaño. Sin embargo, los rasgos que son controlados por una serie alélica o múltiples loci, no son caracteres mendelianos, y no están sujetos a proporciones mendelianas.[31]
Pulos y Hutt sabían que si el alelo que creó una capa blanca era recesiva, a continuación, los caballos blancos tendrían que ser homocigotos para la condición y, por lo tanto, la cría de los caballos blancos juntos siempre resultaría en un potro blanco. Sin embargo, esto no pasó en su estudio y se llegó a la conclusión de que no era blanco recesivo. Por el contrario, si una capa blanca era autosómica dominante simple, los caballos w/w serían no blancos, mientras que los caballos W/w y W/W serían blancos, y este siempre produciría descendencia blanca. Pero Pulos y Hutt no observaron ningún caballo blanco que siempre produjera descendencia blanca, lo que sugiere que los caballos blancos dominantes homocigóticos (W/W) no existían. Como resultado, Pulos y Hutt concluyeron que el blanco era semidominante y letal en homocigosis: los caballos w/w eran no-blancos, W/w eran blancos y W/W murieron.[32]
Pulos y Hutt informaron que las tasas de mortalidad neonatal en los potros blancos eran similares a las de los potros no blancos, y llegaron a la conclusión de que los fetos blancos homocigotos murieron durante la gestación.[33] No se encontraron fetos abortados, lo que sugiere que la muerte se produjo al principio del embrión o el desarrollo del feto y que el feto fue reabsorbido.[34]
Antes del trabajo de Pulos y de Hutt, los investigadores estaban divididos en el modo de herencia del blanco y si era deletéreo (nocivo).[35] La investigación reciente ha descubierto varias vías genéticas posibles a una capa blanca, así que las disparidades en estos hallazgos históricos pueden reflejar la acción de diversos genes. También es posible que los orígenes variados de los caballos blancos de Pulos y Hutt pudieran ser responsables de la falta de homocigotos. Por lo tanto, queda por probarse si todas las mutaciones blancas dominantes equinas causan letalidad embrionaria en el estado homocigótico.[14]
El locus (W) blanco se reconoció por primera vez en ratones en 1908.[36] La mutación del mismo nombre produce una mancha en el vientre, pelos blancos intercalados en la cara dorsal de la capa en el heterocigoto (W/+), y negro de ojos blancos en el homocigoto (W/W). Mientras que los heterocigóticos son saludables, los ratones homocigóticos W tienen anemia macrocítica severa y mueren en cuestión de días.[37] Una mutación que afecta a múltiples sistemas son pleiotrópicas. Después de ser mapeado el gen KIT al locus W en 1988, los investigadores comenzaron a identificar otras mutaciones como parte de una serie alélica de W.[38] Hay más de 90 alelos W conocidos, cada uno representando una única mutación en el gen KIT que produce manchas blancas desde puntos pequeños de la cabeza a capas plenamente blancas, anemia macrocítica de leve a mortal, y esterilidad.[37] Algunos alelos, como una faja, producen manchas blancas solas, mientras que otros afectan a la salud del animal, incluso en el estado heterocigoto. Los alelos que codifican pequeñas cantidades de blanco no tienen más probabilidades de estar vinculados con anemia y la esterilidad que los que codifican un blanco visible. Actualmente, no hay evidencia anecdótica o investigación que sugiera que las mutaciones KIT de equinos afectan a la salud o la fertilidad.[39] Un estudio reciente mostró que los parámetros sanguíneos en los caballos con la mutación W1 fueron normales.[23]
Caballos "blancos" que no son blanco dominante

Los caballos blancos son símbolos potentes en muchas culturas.[40] Una serie de colores de la capa del caballo pueden ser identificados erróneamente como blanco, a pesar de que muchos son genéticamente distintos del "blanco dominante".
Los caballos albinos nunca han sido documentados a pesar de las referencias que se tienen sobre los caballos "albinos".[41][42] El blanco dominante es causado por la ausencia de células de pigmento (melanocitos), mientras que los animales albinos tienen una distribución normal de los melanocitos.[43] Además, un diagnóstico de albinismo en los seres humanos se basa en la discapacidad visual, que no ha sido descrita en caballos con colores blancos dominantes o similares de la capa.[44] En otros mamíferos, el diagnóstico de albinismo se basa en la alteración de la producción de tirosinasa a través de defectos en el gen del color.[45] No hay mutaciones de la tirosinasa o gen C conocidas en caballos.[46]
Colores no blancos
- Caballos Cremello o crema ojiazulado tienen la piel rosada,[47] ojos azul claro y la capa de color crema, lo que indica que las células de pigmento y el pigmento están presentes en la piel, ojos y pelo, pero a niveles más bajos.[48] Los caballos blancos dominantes no tienen células de pigmento, y por lo tanto no hay pigmento en el piel o capa. Además, los caballos blancos dominantes rara vez tienen los ojos azules.[7] Otros factores genéticos, o combinaciones de factores genéticos, tales como el gen perla o gen champagne, también pueden producir capas cremello parecidas. Estos colores de la capa pueden ser distinguibles del blanco dominante por parte de sus ojos inusualmente coloreados.[49]
- Los caballos grises nacen de cualquier color y sustituyen progresivamente su capa de color con pelos grises y blancos. La mayoría de los caballos grises tienen la piel oscura, a menos que lleven también genes para la piel rosada o sin pigmentar. A diferencia de los caballos blancos dominantes, los caballos grises no nacen blancos, ni su color de piel se afecta por su cambio de color del pelaje.[50]
- Los caballos de complejo leopardo, tales como las razas Appaloosa y Knabstrupper, son genéticamente muy distintos de todos los otros patrones de manchas blancas. El patrón de leopardo "fewspot", sin embargo, puede asemejarse a blanco. Hay dos factores que influyen en la aparición eventual de una capa compleja de leopardo: ya sea una copia o dos copias de los alelos leopardo presentes, y el grado de densidad del patrón blanco asociado a un leopardo que está presente al nacer.[51] Si un potro es homocigoto para el alelo LP y tiene un amplio patrón blanco denso, aparecerán casi blanco al nacer, y pueden continuar aclarándose con la edad. En otras partes del mundo, estos caballos se llaman "blancos nacidos."[52][53] Los potros "blancos nacidos" son menos comunes entre los caballos Appaloosa que tienden a tener mantas y barnices ruanos, tienden a ser leopardos completos.[54]

- Tovero, sombrero de Medicina o capo de guerra son términos que a veces se aplican a los caballos pintos con áreas residuales que no son blancos solamente alrededor de la cabeza, sobre todo las orejas, mientras que la mayoría del resto de la capa es de color blanco.[55] Si bien los caballos blancos dominantes pueden tener áreas de pigmento residuales solamente alrededor de las orejas, el término "sombrero de la medicina", por lo general se refiere a los caballos conocidos con genes blancos manchados:tobiano, combinados con overo de marco, sabino o blanco salpicado.[56]
Blanco letal
Los potros con síndrome blanco letal (LWS) tienen dos copias del gen overo y nacen con capa blanca o casi blanca y piel rosada. Sin embargo, a diferencia de los caballos blancos dominantes, los potros con LWS nacen con un colon subdesarrollado que es intratable, y si no es eutanasia, invariablemente mueren de cólicos a los pocos días de nacer.[57]
Los caballos que llevan un solo alelo del gen LWS son saludables y normalmente exhiben el patrón de "marco overo" manchado. En los casos de los caballos "sólidos" con ascendencia overo marco, fenotipo "overo" incierto (no tobiano), o caballos con múltiples patrones, el alelo LWS se puede detectar mediante la prueba de ADN.[58]
Sabino
Tanto el blanco dominante y los caballos "Sabino-blancos" se identifican por ser todos blancos o casi blancos y tener capas con piel de color rosa ojos obscuros, a menudo con pigmento residual a lo largo de la línea media dorsal dominante. Sin embargo, hay diferencias genéticas. El término "blanco dominante" está reservado para los alelos W conocidos. Los caballos blancos dominantes son heterocigotos para cualquiera de los 11 alelos conocidos del gen KIT (por ejemplo W8/+) La homocigosidad para algunos de los 11 alelos conocidos no pueden crear un embrión viable. En contraste, los caballos Sabino-blancos son homocigotos para el Sabino 1 alelo del gen KIT (SB1/SB1).[59][60]

Otro tipo de patrón Sabino, llamado simplemente "Sabino"; "Sabino mínimo" si es muy leve;o si es particularmente dramático, "Sabino máximo", se refiere a los caballos que dan negativo para cualquiera de los alelos blanco dominante, negativo para Sabino 1, y también negativo para Tobiano y marco overo.[61] En un principio, el blanco dominante se separó de Sabino a razón de que los primeros tenían que ser totalmente blancos, mientras que el último podría poseer algo de pigmento. Sin embargo, los estudios de blanco dominante de 2007 y 2009, mostraron que muchos alelos blancos dominantes producen una gama de fenotipos blancos que incluyen caballos con manchas de pigmentación en el cabello y la piel.[2] Cada una de las familias más grandes estudiadas de blanco dominante incluyeron caballos blancos puros, caballos descritos como que tenían marcas blancas "sabino", así como los caballos blancos que se describen como "Sabino máximo."[2][6]
Recientemente, el blanco dominante y sabino, se distinguieron entre sí por los alelos de blancos dominantes que producen embriones no viables en el estado homocigótico, mientras que sabino 1 era viable cuando eran homocigotos.[62] Sin embargo, no todos los alelos KIT actualmente identificados como "blanco dominante " se han demostrado letales.[22]
Las similitudes entre el blanco dominante, Sabino 1, y otras formas de Sabino pueden reflejar su origen molecular común: La serie W1-W11 y SB1 se han mapeado a KIT. Los investigadores que asignan Sabino 1, sugirieron que otros patrones parecidos a sabino, también podrían mapearse a KIT.[63] Del mismo modo, los principales alelos de las piernas blancas y las marcas faciales también se han mapeado en o cerca del gen KIT.[64]
Mosaicismo
El mosaicismo en los caballos se piensa que da cuenta de algunas ocurrencias espontáneas de los caballos blancos, casi blancos, manchados, y ruanos.[65] El mosaicismo se refiere a las mutaciones que se producen después de la etapa de una única célula, y por lo tanto afecta solo una parte de las células adultas.[66] El mosaicismo puede ser una posible causa de la rara aparición de la coloración atigrada en los caballos.[67] Los caballos blancos con mosaicismo serían visualmente indistinguibles de los blancos dominantes. El mosaicismo podría producir potros blancos o parcialmente blancos si una célula madre en el desarrollo del potro sufrió una mutación, o un cambio al ADN, que resultó en piel y cabello sin pigmentar. Las células que descienden de la célula madre afectada exhiben la mutación, mientras que el resto de las células no se ven afectadas.
Una mutación mosaico puede o no ser heredable, dependiendo de las poblaciones de células afectadas.[68] Aunque esto no es siempre el caso, las mutaciones genéticas pueden ocurrir de forma espontánea en una célula sexual de un padre durante la gametogénesis.[69] En estos casos, las llamadas mutaciones en la línea germinal, la mutación estará presente en el cigoto unicelular concebido desde el espermatozoide o el óvulo afectado, y la condición puede ser heredada por la siguiente generación.[65]
Condiciones homólogas

En los seres humanos, una condición de la piel llamada piebaldismo es causada por más de una docena de mutaciones diferentes en el gen KIT. El piebaldismo en el ser humano se caracteriza por un copete blanco, y parches sin pigmento en la piel de la frente, cara, tronco y extremidades ventrales. Fuera de la pigmentación, el piebaldismo es una condición de algún modo benigna.[70] En los cerdos, el "parche", "con cinturón," y los "colores blancos" comerciales son causados por mutaciones en el gen KIT.[71] El modelo más conocido de la función del gen KIT es el ratón en el que se han descrito más de 90 alelos. Los diferentes alelos producen desde dedos blancos, ratones blancos de ojos negros hasta pandas blanquecinos salpicados. Muchos de estos alelos son letales en el estado homocigótico, letal cuando se combinan, o subletal debido a la anemia. Los ratones machos con mutaciones de KIT son a menudo estériles.[72] Hasta la fecha, no hay tales efectos pleiotrópicos descritos en los caballos con mutaciones de KIT.
Referencias
- Pulos WL, Hutt FB (1969). «Lethal dominant white in horses». The Journal of Heredity 60 (2): 59-63. PMID 5816567.
- Haase B, Brooks SA, Tozaki T (octubre de 2009). «Seven novel KIT mutations in horses with white coat colour phenotypes». Animal Genetics 40 (5): 623-9. PMID 19456317. doi:10.1111/j.1365-2052.2009.01893.x.
- Bailey, Ernest; Brooks, Samantha A. (2013). Horse genetics (2. edición). Wallingford: CABI. pp. 56-59. ISBN 9781780643298.
- Pulos, WL; FB Hutt. «Lethal Dominant White in Horses». Journal of Heredity.
- Sturtevant, AH (1912). «A critical examination of recent studies on coat colour inheritance in horses». Journal of Genetics 2 (1): 41-51. doi:10.1007/BF02981546. «"The colour of skin is white or so-called pink, usually with a few small dark specks in skin. Some have a great many dark spots in skin. These latter usually have a few dark stripes in hoofs; otherwise the hoofs are almost invariably white. Those that do not have dark specks in skin usually have glass or watch eyes, otherwise dark eyes ... I have one colt coming one year old that is pure white, not a coloured speck on him, not a coloured hair on him, and with glass eyes." [WP Newell] The term "glass eye" means a white eye. Therefore the colt described above is almost an albino in appearance. However, his sire is one of the dark-eyed somewhat spotted whites, his dam being a brown Trotter. Since "glass" eyes occur not infrequently in pigmented horses it seems probable that this white-eyed albino (?) is really an extreme case of spotting, plus an entirely independent "glass" eye. Mr Newell writes that white mated to white gives about 50% white to 50% pigmented. He reports only three matings of white to white. The results of these were, one white, one roan, and one gray. »
- Haase B, Brooks SA, Schlumbaum A (noviembre de 2007). «Allelic heterogeneity at the equine KIT locus in dominant white (W) horses». PLoS Genetics 3 (11): e195. PMC 2065884. PMID 17997609. doi:10.1371/journal.pgen.0030195.
- Haase B, Brooks SA, Schlumbaum A (noviembre de 2007). «Allelic heterogeneity at the equine KIT locus in dominant white (W) horses». PLoS Genetics 3 (11): e195. PMC 2065884. PMID 17997609. doi:10.1371/journal.pgen.0030195. «Eyes are normally pigmented in dominant white horses, probably due to the different origin of the retinal melanocytes, which develop from local neuroectoderm and not from the neural crest, as do the skin melanocytes. »
- Haase B, Brooks SA, Schlumbaum A (noviembre de 2007). «Allelic heterogeneity at the equine KIT locus in dominant white (W) horses». PLoS Genetics 3 (11): e195. PMC 2065884. PMID 17997609. doi:10.1371/journal.pgen.0030195. «The dominant white coat colour is phenotypically very similar to the coat colour in homozygous sabino horses ».
- «American Creme and White». Breeds of Livestock. Oklahoma State University. 3 de mayo de 1999. Archivado desde el original el 9 de diciembre de 2009. Consultado el 20 de junio de 2009.
- «Camarillo White Horse Association (CWHA)». Camarillo White Horse Association. 17 de marzo de 2009. Consultado el 3 de julio de 2009.
- «Nomenclature History, Kit». Mouse Genome Informatics. The Jackson Laboratory. Consultado el 30 de junio de 2009.
- Sturtevant, AH (1912). "Since "glass" eyes occur not infrequently in pigmented horses it seems probable that this white-eyed albino [sic] is really an extreme case of spotting, plus an entirely independent "glass" eye."
- Pulos, WL; FB Hutt (1969). «Lethal Dominant White in Horses». Journal of Heredity.
- Haase, B. et al (2007). "However, this report on the embryonic lethality was derived from the analysis of offspring phenotype ratios in a single herd segregating one or more unknown mutations."
- Wriedt, C (1924). «Vererbungsfaktoren bei weissen Pferden im Gestut Fredriksborg». Zeitschrift für Tierzuchtung und Zuchtungsbiologie 1: 231-242. doi:10.1111/j.1439-0388.1924.tb00195.x.
- WL Pulos & FB Hutt (1969). "Although Wriedt referred to Sturtevant's report in his genetic analysis of records of the Frederiksborg white horses, he considered the latter to be recessive whites, with homozygotes white, white with gray spots, or gray white ("weissgraue"). Heterozygotes were believed to vary all the way from dilute gray to full color."
- Beth Mead (9 de septiembre de 2005). «International Horse & Pony Colour Term Dictionary Online (Part 2)». Consultado el 8 de julio de 2009.
- WL Pulos & FB Hutt (1969). "In the light of more recent evidence, these conclusions now seem to have been erroneous..."
- WL Pulos & FB Hutt (1969). "Odriozola added no new data on dominant white, but... suggested that different forms of W arranged linearly in the chromosome might be responsible for the differing degrees of white... and that the expression of white is also influenced by modifying genes."
- «Sabino». Equine Color. Archivado desde el original el 25 de diciembre de 2008. Consultado el 9 de diciembre de 2011. «The problem with this theory was that many horses who were "Dominant White" didn't have the necessary "Dominant White" parent, the rule of dominant genes being that at least one parent must have it for the foal to have it. Research has shown that these "Dominant White" horses who are usually born from non-white parents are really Sabinos with the maximum expression of the pattern. The theory of the "Dominant White" gene has been left in the past, especially since there has never been any scientific evidence to support that it exists ».
- Dominant White - Horse Coat Pattern Genetic Testing
- Haase, B. et al (2007) "While [homozygous lethality] is certainly likely for the two nonsense mutations found in Franches-Montagnes Horses and Arabians, it should not necessarily be assumed for the two reported missense mutations or for any of the other as-yet unknown W mutations."
- Haase, B; Obexer-Ruff G; Dolf G; Rieder S; Burger D; Poncet PA; Gerber V; Howard J et al. (9 de abril de 2009). «Haematological parameters are normal in dominant white Franches-Montagnes horses carrying a KIT mutation.». Veterinary Journal 184 (3): 315-7. PMID 19362501. doi:10.1016/j.tvjl.2009.02.017.
- Castle, Nancy (19 de mayo de 2009). «Equine KIT Gene Mutations». Archivado desde el original el 30 de mayo de 2009. Consultado el 18 de junio de 2009.
- Hauswirth, Regula; Jude, Rony; Haase, Bianca; Bellone, Rebecca R.; Archer, Sheila; Holl, Heather; Brooks, Samantha A.; Tozaki, Teruaki; Penedo, Maria Cecilia T.; Rieder, Stefan; Leeb, Tosso (diciembre de 2013). «Novel variants in the KIT and PAX3 genes in horses with white-spotted coat colour phenotypes». Animal Genetics 44 (6): 763-765. doi:10.1111/age.12057. Consultado el 30 de junio de 2015.
- Thiruvenkadan, AK; N Kandasamy; S Panneerselvam (2008). «Review: Coat colour inheritance in horses». Livestock Science (Elsevier) 117 (2-3): 109-129. doi:10.1016/j.livsci.2008.05.008. «During embryogenesis the pigment cells (melanocytes) migrate to specific sites on either side of the body as well as the backline. There are three such sites on the head (near the eye, ear, and top of the head), and six sites along each side of the body, and several along the tail. A few pigment cellsmigrate to each of these sites, there they proliferate and migrate outwards, joining up to form larger patches, spreading down the legs and down the head until they meet up under the chin, and down the body until they meet up on the belly (Cattanach, 1999). »
- Rieder S, Hagger C, Obexer-Ruff G, Leeb T, Poncet PA (2008). «Genetic analysis of white facial and leg markings in the Swiss Franches-Montagnes Horse Breed». The Journal of Heredity 99 (2): 130-6. PMID 18296388. doi:10.1093/jhered/esm115. «Phenotypes may vary from tiny depigmentated body spots to white head and leg markings, further on to large white spotting and finally nearly complete depigmentation in white-born horses... White markings result from the lack of melanocytes in the hair follicles and the skin... A completely pigmented head or leg depends on the complete migration and clonal proliferation of the melanoblasts in the mesoderm of the developing fetus, thus ensuring that limbs and the head acquire a full complement of melanocytes ».
- Kurakin A (enero de 2005). «Self-organization vs Watchmaker: stochastic gene expression and cell differentiation». Development Genes and Evolution 215 (1): 46-52. PMID 15645318. doi:10.1007/s00427-004-0448-7.
- Woolf CM (1995). «Influence of stochastic events on the phenotypic variation of common white leg markings in the Arabian horse: implications for various genetic disorders in humans». The Journal of Heredity 86 (2): 129-35. PMID 7751597.
- Haase, B. et al. (2007) "In one study, white horses were shown to be obligate heterozygous (W/+), as the W/W genotype was hypothesized to cause early embryonal lethality [4]."
- Strachan, Tom & Read, Andrew (1999) [1996]. «Genes in pedigrees: 3.4 Nonmendelian characters». En Kingston, Fran, ed. Human Molecular Genetics. BIOS Scientific Publishers (2 edición). New York: John Wiley & Sons. p. 333. ISBN 1-85996-202-5. Consultado el 10 de julio de 2009.
- Pulos & Hutt (1969). "Each of the five white stallions used in the stud sired one or more colored foals. Similarly, all of the eight white mares that were adequately tested produced at least one colored foal. The fact that these 13 white horses were all proven to be heterozygotes agrees with previous reports that white horses with colored eyes did not breed true to type, but always produced some colored progeny. This, in turn, suggests that the genoytpe W/W is not viable."
- Pulos & Hutt (1969). "Among six white foals (from parents both white) that died soon after birth, one had been unable to stand and nurse; death of another was attributed to exposure, one was strangled and another killed by the mare. The possibility that any of these might have been homozygotes is refuted by the fact that similar conditions caused death of several foals from the colored pony mares. Some of those foals were white, and some colored, but none could have been W/W."
- Pulos & Hutt (1969). "As aborted foetuses were not found although a constant watch was maintained for them, it is possible that the homozygotes die early in gestation and are resorbed."
- Pulos & Hutt (1969). "... in his genetic analysis of records of the Frederiksborg white horses, [Wriedt] considered [them] to be recessive whites, with homozygotes white, white with gray spots, or gray white ("weissgraue") ... He considered that the gene for white could not itself be lethal because four fertile white mares produced from 46 matings a total of 37 foals, none of which was dead or weak, and that good record (80 percent fertility) was better than could have been expected if the gene for white color were lethal. Subsequently von Lehmann-Mathildenhoh reported evidence of a dominant white in the Bellschwitz and Ruschof studs ... He did not consider the possibility that it might be associated with any lethal action ... [Salisbury] made no reference to effects of the gene in homozygotes ... Berge lists dominant white horses as heterozygotes, and follows Castle in suggesting that homozygosity for W is lethal."
- Durham, F.M. A preliminary account of the inheritance of coat colour in mice. Reports to the Evolution Committee IV: 41-53, 1908.
- Silvers, Willys K. (1979). «10: Dominant Spotting, Patch, and Rump-White». The Coat Colors of Mice: A Model for Mammalian Gene Action and Interaction. Springer Verlag. ISBN 0-387-90367-4. Consultado el 3 de julio de 2009.
- Chabot, Benoit; Dennis A. Stephenson; Verne M. Chapman; Peter Besmer; Alan Bernstein (1 de septiembre de 1988). «The proto-oncogene c-kit encoding a transmembrane tyrosine kinase receptor maps to the mouse W locus». Nature 335 (6185): 88-9. PMID 2457811. doi:10.1038/335088a0.
- Haase, B. et al (2009). "Currently, there is little known about possible pleiotropic effects of KIT mutations in horses."
- Cooper, JC (1978). «Horse». An Illustrated Encyclopedia of Traditional Symbols. London: Thames & Hudson. pp. 85–6. ISBN 0-500-27125-9. «... [T]he white horse ... represents pure intellect; the unblemished; innocence; life and light, and is ridden by heroes. »
- Castle, William E (1948). «The ABC of Color Inheritance in Horses». Genetics 33 (1): 22-35. PMC 1209395. PMID 17247268. «No true albino mutation of the color gene is known among horses, though several varieties of white horse are popularly known as albinos. »
- O'Hara, Mary (1941). My Friend Flicka. Lippincott. ISBN 0-06-080902-7.
- Silvers, Willys K. (1979). «3: The b-Locus and c (Albino) Series of Alleles». The Coat Colors of Mice: A Model for Mammalian Gene Action and Interaction. Springer Verlag. p. 59. Consultado el 7 de julio de 2009. «... the inability of albino animals to produce pigment stems not from an absence of melanocytes ».
- «What is Albinism?». The National Organization for Albinism and Hypopigmentation. Archivado desde el original el 14 de mayo de 2012. Consultado el 7 de julio de 2009.
- Cheville, Norman F (agosto de 2006). Introduction to veterinary pathology (3 edición). Wiley-Blackwell. ISBN 978-0-8138-2495-6. «Albinism results from a structural gene mutation at the locus that codes for tyrosinase; that is, albino animals have a genetically determined failure of tyrosine synthesis. »
- Castle, William E. (1948). «The ABC of Color Inheritance in Horses». Genetics 33 (1): 22. «No true albino mutation of the color gene is known among horses, though several varieties of white horse are popularly known as albinos. »
- «Facts and Myths». Cream Gene Information. Cremello and Perlino Education Association. Archivado desde el original el 7 de febrero de 2012. Consultado el 8 de julio de 2009.
- Mariat, Denis; Sead Taourit; Gérard Guérin (2003). «A mutation in the MATP gene causes the cream coat colour in the horse.». Genet. Sel. Evol. (INRA, EDP Sciences) 35 (1): 119-133. PMC 2732686. PMID 12605854. doi:10.1051/gse:2002039.
- «Champagne-Cream Combinations». International Champagne Horse Registry. Consultado el 4 de junio de 2009.
- Rosengren Pielberg G, Golovko A, Sundström E (agosto de 2008). «A cis-acting regulatory mutation causes premature hair graying and susceptibility to melanoma in the horse». Nature Genetics 40 (8): 1004-9. PMID 18641652. doi:10.1038/ng.185.
- Bellone, Rebecca R; Samantha A Brookers; Lynne Sandmeyer; Barbara A Murphy; George Forsyth; Sheila Archer; Ernest Bailey; Bruce Grahn (agosto de 2008). «Differential Gene Expression of TRPM1, the Potential Cause of Congenital Stationary Night Blindness and Coat Spotting Patterns (LP) in the Appaloosa Horse (Equus caballus)». Genetics (Genetics Society of America) 179 (4): 1861-1870. PMC 2516064. PMID 18660533. doi:10.1534/genetics.108.088807. «A single autosomal dominant gene, leopard complex (LP), is thought to be responsible for the inheritance of these patterns and associated traits, while modifier genes are thought to play a role in determining the amount of white patterning that is inherited (Miller 1965; Sponenberg et al. 1990; S. Archer and R. R. Bellone, unpublished data) ».
- «Rules & Knabstrupper Breed Standard of the German ZfDP Registry». UK Knabstrupper Association. Archivado desde el original el 29 de mayo de 2009. Consultado el 20 de junio de 2009.
- «Die Farbmerkmale» (en alemán). Knabstrupper.de. Archivado desde el original el 18 de junio de 2009. Consultado el 20 de junio de 2009.
- Sponenberg, Dan Phillip (2003). «5. Patterns Characterized by Patches of White». Equine color genetics (2 edición). Wiley-Blackwell. p. 94. ISBN 0-8138-0759-X. «... most Appaloosas have a blanket or varnish roan phenotype ... In the Noriker breed most horses with LpLp are leopard, and the few varnish roans or blanketed horses in the breed tend to produce leopards more than their own blanket or varnish roan pattern ».
- Vrotsos, Paul D.; Elizabeth M. Santschi; James R. Mickelson (2001). «The Impact of the Mutation Causing Overo Lethal White Syndrome on White Patterning in Horses». Proceedings of the Annual Convention of the AAEP (American Association of Equine Practitioners) 47: 385-391. «This is a rare color pattern in which the coat is almost entirely white (Fig. 6). Pigmented areas are found primarily on the ears and poll, but may also appear on the thorax, flank, dorsal midline, and tail head. Medicine hat horses can arise from overo or tovero bloodlines; when of overo bloodlines, medicine hat horses may have pigment that is quite faint on the dorsal midline. »
- Janet Piercy (2001). «Breed Close Up Part II». The Colorful World of Paints & Pintos. International Registry of Colored Horses. Archivado desde el original el 24 de febrero de 2009. Consultado el 3 de julio de 2009. «The perfectly marked medicine hat is usually a tovero, but these horses can be overos and tobianos too ».
- Metallinos, DL; Bowling AT; Rine J (junio de 1998). «A missense mutation in the endothelin-B receptor gene is associated with Lethal White Foal Syndrome: an equine version of Hirschsprung Disease». Mammalian Genome (New York: Springer New York) 9 (6): 426-31. PMID 9585428. doi:10.1007/s003359900790. Consultado el 4 de septiembre de 2008.
- «Equine Coat Color Tests». Veterinary Genetics Laboratory. UC Davis. Consultado el 8 de julio de 2009.
- Brooks, Samantha; Ernest Bailey (2005). «Exon skipping in the KIT gene causes a Sabino spotting pattern in horses». Mammalian Genome (PDF) 16 (11): 893-902. PMID 16284805. doi:10.1007/s00335-005-2472-y. «Chapter 3 ».
- UC Davis. «Horse Coat Color Tests». Veterinary Genetics Laboratory. University of California - Davis. Consultado el 8 de julio de 2009. «Horses with 2 copies of the Sabino1 gene, are at least 90% white and are referred to as Sabino-white. »
- Castle, Nancy (2009). "It has been the belief of horse enthusiasts that true “white” horses were always completely white with no retained pigment, and that if a horse retained some pigment of the skin and/or hair, it was genetically some form of sabino if it were not the result of other known white spotting patterns (tobiano, frame overo, splash white, etc.)"
- Castle, Nancy (2009). "KIT mutations that cause depigmentation generally ranging from approximately 50% depigmented to all white phenotypes, and are also predicted to be embryonic lethal when homozygous, are classified as Dominant White. Mutations that are viable in the homozygous state are categorized as Sabino."
- Brooks, Samantha (2005). "Presumably variation at other genetic sites within KIT, or another gene, is responsible for those sabino phenotypes."
- Rieder, Stefan et al (2008). "Our association analysis indicated that the putative major gene for white markings is located at or near the KIT locus."
- Haase, B. et al (2009). "Whenever a white foal is born out of solid-coloured parents, the most likely explanation is a KIT mutation in the germline of one of its parents or alternatively a mutation in the early developing embryo itself, which might lead to mosaic foals."
- Strachan, Tom & Read, Andrew (1999) [1996]. «Genes in pedigrees: 3.2 Complications to the basic pedigree patterns». En Kingston, Fran, ed. Human Molecular Genetics. BIOS Scientific Publishers (2 edición). New York: John Wiley & Sons. p. 297. ISBN 1-85996-202-5. Consultado el 8 de julio de 2009. «Post-zygotic mutations produce mosaics with two (or more) genetically distinct cell lines. [...] Mutations occurring in a parent's germ line can cause de novo inherited disease in a child. When an early germ-line mutation has produced a person who harbors a large clone of mutant germ-line cells (germinal, or gonadal, mosaicism), a normal couple with no previous family history may produce more than one child with the same serious dominant disease ».
- Kay L. Isaac. «Brindle Information». American Brindle Equine Association. Consultado el 8 de julio de 2009. «One only outwardly appearing brindle that is likely the result of a mosaic or chimeric equine ... » (enlace roto disponible en Internet Archive; véase el historial, la primera versión y la última).
- Haase, B. et al (2009) "our study included several founder animals where mosaicism cannot be excluded. One example for such a scenario is the W8 allele observed in a single "mottled" Icelandic horse, which represents the founder animal for this mutation (Fig. 1g). This horse might be a mosaic, and it remains to be determined whether it will consistently produce offspring with the mottled phenotype."
- Strachan, Tom & Andrew Read (1999) "A common assumption is that an entirely normal person produces a single mutant gamete. However, this is not necessarily what happens. Unless there is something special about the mutational process, such that it can happen only during gametogenesis, mutations may arise at any time during post-zygotic life."
- Michael D. Fox; Camila K. Janniger (30 de enero de 2009). «Piebaldism». eMedicine. WebMD. Consultado el 20 de junio de 2009.
- Pielberg G, Olsson C, Syvänen AC, Andersson L (enero de 2002). «Unexpectedly high allelic diversity at the KIT locus causing dominant white color in the domestic pig». Genetics 160 (1): 305-11. PMC 1461930. PMID 11805065.
- Silvers, Willys K. (1979). The Coat Colors of Mice. Springer Verlag. ISBN 0-387-90367-4.
| Control de autoridades |
|
|---|
 Datos: Q2729941
Datos: Q2729941 Multimedia: Dominant white horse / Q2729941
Multimedia: Dominant white horse / Q2729941