Phaeophyceae
Las algas pardas o feofíceas o marrones (clase Phaeophyceae) son un grupo de algas incluido en el reino Protista. No son verdaderas plantas pues se clasifican en el grupo Heterokonta. Comprende unos 265 géneros con unas 1500-2000 especies,[2] principalmente marinos pues solo seis géneros son de agua dulce. Las algas pardas son los principales productores primarios de varias comunidades de animales y protistas.
| Algas pardas | ||
|---|---|---|
 Algas pardas | ||
| Taxonomía | ||
| Dominio: | Eukarya | |
| Reino: | Protista | |
| (sin rango) | SAR o Harosa | |
| Superfilo: | Heterokonta | |
| Filo: | Ochrophyta | |
| Subfilo: | Chrysista | |
| Infrafilo: | Marista | |
| Superclase: | Fucistia | |
| Clase: |
Phaeophyceae de Bary 1881, Hauck 1885, Hansgirg 1886 | |
| Órdenes[1] | ||
| ||
La mayoría viven en las costas rocosas de las zonas templadas y subpolares, dominando la zona intermareal como en el caso de las Fucales y Laminariales. Macrocystis, que forma bosques submarinos prominentes, puede alcanzar hasta 60 m de longitud. También se presentan en formas flotantes libres, como Sargassum, que forma grandes extensiones (del tamaño de una provincia) en el Mar de los Sargazos.
Las algas pardas comprenden formas pluricelulares con tejidos diferenciados. Generalmente presentan rizoide, estipe y láminas. El estipe de algunas formas puede estar provisto de un tamiz de tubos capaces de transportar agua y productos de la fotosíntesis. Como pigmentos presentan clorofilas a, c1 y c2 y fucoxantina. Este último es el responsable del color pardo que presentan estas algas. Los cloroplastos están rodeados de cuatro membranas, por lo que se supone que son el resultado de la endosimbiosis secundaria de un alga roja.
Morfología
Formas y tamaños
Existen algas pardas de una amplia variedad de tamaños y formas. Los miembros más pequeños del grupo constan de solo unas pocas células, por lo que constituyen algas microscópicas. Otras crecen como penachos de células filiformes de no más de unos pocos centímetros de largo.[3] Otros grupos de algas pardas crecen hasta tamaños mucho más grandes, por ejemplo, kelp y sargazos son a menudo las algas más visibles en sus hábitats.[4] El kelp puede variar en tamaño desde los 50 cm de la palma de mar (Postelsia) hasta los 60 m del alga gigante Macrocystis pyrifera, que es la mayor de todas las algas conocidas.[5][6] Con respecto a su forma, las algas pardas comprenden desde pequeñas costras pegadas al sustrato hasta frondosas matas flotantes formadas por las especies de sargazos.[7] Se incluyen desde delicadas hebras de células, como en Ectocarpus, hasta anchas ramas aplanadas en forma de abanico, como en Padina.
Sin importar el tamaño o forma, dos características visibles distinguen a Phaeophyceae de todas las demás algas. En primer lugar, los miembros del grupo presentan un color característico que va desde verde oliva a varios tonos de marrón. El tono particular depende de la cantidad de fucoxantina presente en el alga.[8] En segundo lugar, todas las algas pardas son pluricelulares. No hay especies conocidas que existan como células individuales o como colonias de células,[8] por lo que las algas pardas son el único grupo importante de algas que no incluye tales formas. Sin embargo, esto puede ser el resultado de la clasificación y no una consecuencia de la evolución, ya que todos los grupos que son parientes cercanos de las algas pardas incluyen formas unicelulares o coloniales.

Estructuras visibles

Cualquiera que sea su forma, el cuerpo de las algas pardas se denomina talo, lo que significa que carecen de los complejos tejidos xilema y floema de las plantas vasculares. Esto no significa que las algas pardas carezcan por completo de estructuras especializadas. Pero, debido a que los botánicos definen (verdaderos) tallos, hojas y raíces por la presencia de estos tejidos, su ausencia en las algas pardas significa que las estructuras equivalentes a tallos y hojas presentes en algunos grupos de algas pardas deben describirse utilizando terminología diferente.[9] Aunque no todas las algas pardas son estructuralmente complejas, típicamente poseen una o más partes características.

Un rizoide es una estructura equivalente a la raíz, presente en la base del alga, que le sirve para fijarse sustrato y que, a diferencia de la raíz, no es utilizado como el órgano principal para la captación de agua ni de nutrientes. La apariencia del rizoide en las algas pardas es variado: puede ser muy ramificado o tener forma de disco o bulbo.
El estipe, equivalente al tallo, puede crecer como una estructura corta en la base del alga, como en Laminaria, o crecer hasta convertirse en una gran estructura, como en Sargassum o Macrocystis. En las algas pardas más estructuralmente diferenciadas, tales como Fucus, los tejidos dentro del estipe se dividen en tres capas o regiones distintas. Estas regiones incluyen una médula central, una corteza circundante y una epidermis externa, cada una de los cuales tiene su análogo en el tallo de una planta vascular. En algunas algas pardas, la región de la médula incluye un núcleo de células alargadas que se asemejan al floema de las plantas vasculares, tanto en estructura como función. En otras (como en Nereocystis), el centro del estipe es hueco y al estar lleno de gas sirve como sostén del alga. El estipe puede ser relativamente flexible y elástico en especies como Macrocystis pyrifera que crecen en corrientes fuertes, o pueden ser más rígidos en especies como Postelsia, que están expuestos a la atmósfera durante la marea baja.
Las láminas son las partes planas de las algas equivalentes a las hojas. Algunas algas pardas presentan una única lámina, mientras que en otras son numerosas. Estas láminas se pueden unir directamente al estipe, directamente al rizoide si el estipe esta ausente, o puede haber una cámara de aire entre el estipe y la lámina. La superficie de la lámina puede ser lisa o arrugada; sus tejidos pueden ser delgados y flexibles o gruesos y correosos. En especies como Egregia menziesii, esta característica puede variar dependiendo de la turbulencia de las aguas en las que crece.[3] En otras especies, la superficie de la lámina se recubre con limo para desalentar la fijación de epifitas o para disuadir a los herbívoros. Las láminas a menudo son también las partes del alga que soportan las estructuras reproductivas.
_-_geograph.org.uk_-_224125.jpg.webp)
Vesículas llenas de gas denominadas neumatocistos proporcionan flotabilidad a muchas laminarias y miembros de las Fucales. Estas estructuras en forma de vejiga se suelen localizar en las láminas, o próximas a ellas, para mantenerlas más cerca de la superficie del agua y recibir más luz para la fotosíntesis. Los neumatocistos son generalmente esféricos o elipsoidales, pero pueden variar en forma entre las diferentes especies. Especies como Nereocystis luetkeana y Pelagophycus porra presentan un único gran neumatocisto entre la parte superior del estipe y la base de las láminas. Por el contrario, el alga gigante Macrocystis pyrifera presenta muchas láminas a lo largo de su estipe, con un neumatocisto en la base de cada lámina, donde se une al estipe central. Las especies de Sargassum también tienen muchas láminas y neumatocistos, pero ambos tipos de estructuras se unen por separado al estipe por tallos cortos. En las especies de Fucus, los neumatocistos se desarrollan dentro de la propia lámina, ya sea como vejigas esféricas individuales o regiones alargadas llenas de gas que siguen el contorno de la lámina en la que se desarrollan.
Crecimiento

Las algas pardas son las algas marinas más grandes y de más rápido crecimiento.[3] Las láminas de Macrocystis puede crecer hasta 50 cm por día, mientras que los estipes puede crecer hasta 6 cm por día.[10]
El crecimiento de algas pardas se produce en el extremo de las estructuras como resultado de las divisiones de una única célula apical o en una fila de tales células.[4] Cuando esta célula apical se divide, las nuevas células que produce dan lugar a todos los tejidos del alga. Las ramificaciones y otras estructuras laterales aparecen cuando la célula apical se divide para producir dos nuevas células apicales. Sin embargo, algunos grupos como Ectocarpus crecen de forma difusa, pues la producción de nuevas células se pueden producir en cualquier parte del talo.[8]
Organización de los tejidos
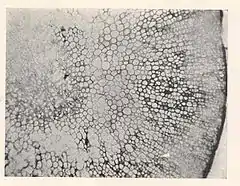
Las algas pardas más simples son filamentosas, es decir, sus células son alargadas con septos de separación entre ellas. Estas se ramifican mediante la formación de extremos más anchos y dividiendo la ampliación.[11]
Aparte de las formas filamentosas, hay dos tipos principales de organización del tejido en las algas pardas: seudoparenquimatoso (haplóstico) y parenquimatoso (polístico).[12] Las láminas pueden ser de tipo multiaxial o monoaxial.
La pared celular se compone de dos capas: la capa interior se compone de celulosa y es resistente, por lo que sirve de sostén, mientras que la capa exterior está compuesta de algina principalmente y es pegajosa cuando está mojada, pero se vuelve dura y quebradiza cuando se seca.[12]
Ciclo vital

Las algas pardas son todas multicelulares y presentan siempre reproducción sexual, aunque existe una gran variabilidad de un grupo a otro. La reproducción sexual puede ser por isogamia, anisogamia u oogamia. En este último caso los gametofitos pueden ser femeninos o masculinos, dando lugar a óvulos y espermatozoos, respectivamente. Estos últimos son biflagelados, con un flagelo mastigonemado dirigido hacia delante y otro liso dirigido hacia atrás. La unión de los gametos puede tener lugar en el agua o, en el caso de la oogamia, en el oogonio. El cigoto germina en respuesta a unas determinadas condiciones de luz. Sin embargo, el ciclo vital muestra una gran variabilidad de un grupo a otro. Así, se distinguen dos tipos de ciclos básicos:
- Monogénetico. Comprende un solo tipo de generación, pues el cigoto da lugar directamente a nuevos gametófitos, cerrando el ciclo. Se presenta en Fucales.
- Digenético. En este caso el organismo diploide que se desarrolla a partir del cigoto recibe el nombre de esporófito y produce esporas. Las esporas se desarrollan y dan lugar a nuevos gametófitos, cerrando el ciclo. Existen tanto ciclos isomórficos (o monomórficos), en los que esporófitos y gametófitos presentan una forma similar, como ciclos heteromórficos (o dimórficos) en los que esporófitos y gametófitos son diferentes. En este último caso puede dominar el esporófito o hacerlo el gametófito. Por ejemplo, el ciclo vital de Laminaria consiste en una generación diploide, que es la gran planta que conoce la mayoría de la gente. Esta produce esporangios de estructura microscópica especializada que se dividen por meiosis antes de que las esporas sean liberadas. Se producen igual número de esporas haploides masculinas y femeninas.[2][13]
Clasificación
Las algas pardas pertenecen al filo Ochrophyta (heterocontos),[14] un extenso grupo de organismos eucariotas que se disntinguen por poseer cloroplastos rodeados por cuatro membranas, lo que sugiere un origen de relación simbiótica entre un eucarionte basal y un eucarionte fotosintético. La mayoría de las algas pardas contienen el pigmento fucoxantina, que proporciona el distintivo color verde-marrón que da nombre al grupo. Las algas pardas son únicas entre los heterocontos en que desarrollan organismos multicelulares con tejidos diferenciados, pero se reproducen por medio de esporas flageladas, que se asemejan a otras células heterocontas. Los estudios genéticos muestran que sus parientes más cercanos son las algas verde-amarillas (Xanthophyceae).
Las algas pardas aparecen en el registro fósil en el Mesozoico, posiblemente ya en el Jurásico. Su presencia como fósil es rara, debido a un cuerpo generalmente blando. Otros grupos de algas como las algas rojas (Rhodophyta) y las algas verdes (Chlorophyta) incluyen algunos miembros calcáreos, que son más propensos a dejar huellas en el registro fósil que los órganos blandos de las algas pardas. Se han encontrado los fósiles del Mioceno de una gran alga parda de cuerpo blando, Julescrania, en buen estado de conservación en formación diatomita de Monterey. Se conocen también algunos otros pocos ejemplares de mayor antigüedad en el registro fósil, aunque su identificación es difícil.[15]
Filogenia
Se han establecido las siguientes relaciones (2014): [16]
| Phaeophyceae |
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Galería
 Saccorhiza polyschides (alga rizada)
Saccorhiza polyschides (alga rizada)






 Laminaria saccharina (fucus azucarado)
Laminaria saccharina (fucus azucarado) Macrocystis pyrifera (hasta 60 m de longitud)
Macrocystis pyrifera (hasta 60 m de longitud)
Referencias
- Silberfeld, T., Rousseau, F., & de Reviers, B. (2014). An updated classification of brown algae (Ochrophyta, Phaeophyceae). Cryptogamie, Algologie, 35(2), 117-156.
- Thomas,D.N. 2002 Seaweeds. The Natural History Museum, London. ISBN 0 56509175 1
- Connor, J.; Baxter, C. (1989). Kelp Forests. Monterey Bay Aquarium. ISBN 1-878244-01-9.
- Dittmer, H. J. (1964). Phylogeny and Form in the Plant Kingdom. Princeton, NJ: D. Van Nostrand Company. pp. 115–137. ISBN 0-88275-167-0.
- Abbott, I. A.; Hollenberg, G. J. (1976). Marine Algae of California. California: Stanford University Press. ISBN 0-8047-0867-3.
- Cribb, A. B. (1953). «Macrocystis pyrifera (L.) Ag. in Tasmanian waters». Australian Journal of Marine and Freshwater Research 5 (1): 1-34. doi:10.1071/MF9540001.
- Jones, W. E. (1962). «A key to the genera of the British seaweeds». Field Studies] 1 (4): 1-32.
- Bold, H. C.; Alexopoulos, C. J.; Delevoryas, T. (1987). Morphology of Plants and Fungi (5th edición). New York: Harper & Row Publishers. pp. 112–131, 174–186. ISBN 0-06-040839-1.
- Raven, P. H.; Evert, R. F.; Eichhorn, S. E. (2005). Biology of Plants (7th edición). New York: W. H. Freeman and Company. pp. 316–321, 347. ISBN 0-7167-1007-2.
- Round, F. E. (1981). The Ecology of Algae. Cambridge: Cambridge University Press. p. 103. ISBN 0-521-26906-7.
- Wynne, M. J. (1981). «The Biology of seaweeds». En Lobban, C. S.; Wynne, M. J., eds. Phaeophyta: Morphology and Classification. Botanical Monographs 17. University of California Press. p. 52. ISBN 978-0-520-04585-9.
- Sharma, O. P (1986). Textbook of Algae. Tata McGraw-Hill. p. 298. ISBN 978-0-07-451928-8.
- Fletcher,R.L. 1987. Seaweeds of the British Isles. Volume 3. Part 1. British Museum (Natural History), London. ISBN 0-565-00992-3
- Guiry, M. D. & G. M. Guiry (2006). «AlgaeBase version 4.2». National University of Ireland, Galway. Consultado el 7 de diciembre de 2006.
- Coyer, J.A.; G.J. Smith, R.A. Anderson (2001). «Evolution of Macrocystis spp. (Phaeophyta) as determined by ITS1 and ITS2 sequences». Journal of Phycology (Blackwell Publishing) 37: 574-585.
- Silberfeld, Thomas; Rousseau, Florence; de Reviers, Bruno (2014). "An Updated Classification of Brown Algae (Ochrophyta, Phaeophyceae)". Cryptogamie, Algologie. BioOne. pp. 117–156. doi:10.7872/crya.v35.iss2.2014.117
Enlaces externos
 Wikimedia Commons alberga una categoría multimedia sobre Phaeophyceae.
Wikimedia Commons alberga una categoría multimedia sobre Phaeophyceae.- Monterey Bay Flora
- Phaeophyceae, National University of Ireland, Galway
- http://www.ucmp.berkeley.edu/chromista/browns/phaeofr.html
| Control de autoridades |
|
|---|
 Datos: Q184573
Datos: Q184573- Multimedia: Phaeophyceae / Q184573
 Especies: Phaeophyceae
Especies: Phaeophyceae