Amaryllidaceae
Las amarilidáceas (Amaryllidaceae) son una familia de plantas herbáceas, perennes, bulbosas o rizomatosas, pertenecientes al orden Asparagales de las monocotiledóneas. Pueden ser fácilmente reconocidas por sus flores trímeras dispuestas en inflorescencias similares a umbelas, las cuales se hallan rodeadas de dos brácteas en la extremidad de un escapo. La familia está compuesta por alrededor de 73 géneros y aproximadamente 1600 especies y se distribuye por todo el mundo. La diversidad de esta familia se halla taxonómicamente organizada en tres subfamilias, a las que hasta hace poco tiempo se les reconocía la categoría de familia: amarilidóideas, alióideas y agapantóideas. Salvo esta última, que presenta un solo género, las dos restantes se dividen en varias tribus.[1]
| Amaryllidaceae | ||
|---|---|---|
 | ||
| Taxonomía | ||
| Reino: | Plantae | |
| División: | Magnoliophyta | |
| Clase: | Monocotyledoneae | |
| Orden: | Asparagales | |
| Familia: |
Amaryllidaceae Burnett | |
| Subfamilias | ||
Las amarilidáceas presentan una considerable importancia económica y cultural. De hecho, muchas de sus especies son utilizadas como condimento, plantas ornamentales, hortícolas y medicinales.[2]
Descripción
_-_8.jpg.webp)



Las amarilidáceas son plantas terrestres, raramente acuáticas o epífitas, herbáceas o suculentas y, excepto cuatro especies, perennes. Presentan bulbos o, más raramente, rizomas —por ejemplo en Clivia, Scadoxus y Agapanthus—. Los bulbos se hallan recubiertos por las bases membranosas de las hojas, formando una estructura que se denomina «túnica». La raíz primaria se halla bien desarrollada.[2]
Las hojas son simples, paralelinervadas, de borde entero, lineares, oblongas, elípticas, lanceoladas o filiformes, sésiles o pecioladas, generalmente arrosetadas, espiraladas o dísticas. Presentan un meristema basal persistente y desarrollo basípeto.



Las flores son hermafroditas, actinomórficas (es decir, de simetría radial) o ligeramente zigomórficas, pediceladas o sésiles, vistosas, cada una asociada a una bráctea filiforme. El perigonio está compuesto de seis tépalos dispuestos en dos verticilos de 3 piezas cada uno, las cuales presentan aproximadamente la misma forma y el mismo tamaño. Los tépalos se hallan libres entre sí o bien pueden estar soldados en la base formando un tubo perigonial o hipanto, el cual se prolonga en algunos casos en una «corona» —también llamada paraperigonio o falsa corola, por ejemplo en Narcissus—, a veces reducido a escamas o dientes poco notables.[3][2]
El androceo presenta seis estambres dispuestos en dos verticilos de tres piezas cada uno —rara vez presentan solo tres estambres, como en Zephyra, o de 9 hasta 18, como en Gethyllis—, opuestos a los tépalos, insertos sobre el tubo del perigonio o la base de los tépalos. Los filamentos son delgados, filiformes o se hallan ensanchados en la base, pueden ser libres (Hippeastrum) o estar unidos entre sí por medio de una membrana que rodea a todo el ovario que se denomina «copa estaminal», como por ejemplo en Hymenocallis. Las anteras son oblongas o lineales.[2] Los estambres a veces tienen apéndices que forman una corona estaminal —por ejemplo en Hymenocallis—.
El gineceo es de ovario ínfero en el caso de las amarilióideas y es de ovario súpero en las agapantóideas y alióideas. Está formado por tres carpelos unidos entre sí y delimitando tres cavidades o lóculos, los cuales contienen desde dos a tres hasta muchos óvulos anátropos de placentación axilar. El ovario se continua con un estilo y con un estigma diminuto, capitado a profundamente trífido. Presentan usualmente nectarios florales y la secreción de néctar proviene del perigonio o del gineceo, usualmente en los septos del ovario.[2][4]
Las flores se hallan dispuestas en inflorescencias determinadas cimosas en la extremidad de un escapo, a veces compactadas y similares a umbelas —por lo que reciben el nombre de «pseudoumbelas»—, raramente reducidas a flores solitarias. Las brácteas involucrales, aquellas que encierran a los pimpollos de las flores, pueden estar presentes o ausentes.[2][4]
El fruto es una cápsula indehiscente o una cápsula loculicida, ocasionalmente puede ser una baya —como por ejemplo en Clivia—. Las semillas son secas a carnosas, aplanadas a globosas, aladas o sin alas y presentan endosperma que no reserva almidón sino aceites. La testa está usualmente incrustada con fitomelaninas (con algunas excepciones, como Amaryllis o Hymenocallis). El color de las semillas usualmente es negro, o también verde o rojo.
Los cromosomas en las amarilidóideas son en general largos, de 3 a 28 µm. Los números cromosómicos básicos son x = 6, 8, 9, 10, 11 y 26; y difieren entre las tribus.[2][4] En el caso de las alióideas, los números cromosómicos básicos son x = 4, 5, 6, 7, 8 y 9 y los cromosomas pueden ser desde relativamente pequeños a grandes (de 2 a 20 µm).[5][1] [6] Las agapantóideas, finalmente, presentan un número cromosómico básico x = 15, si bien se han informado también x = 14 y x = 16, y los cromosomas tienen una longitud de 4 a 9 µm.[7]
Fitoquímica
Las alióideas y amarilidóideas presentan moléculas relacionadas con la defensa, denominadas lectinas, muy particulares y que se unen específicamente a la manosa.[8][9]
Las alióideas presentan saponinas esteroideas, así como compuestos característicos de cada especie y son creados por la transformación química de una serie de compuestos volátiles de azufre generadas por la ruptura de sulfóxidos de S-alquil y S-alquenil cisteínas. El más representativo es la aliina. Al trozar los tejidos del bulbo, se activan las enzimas alinasa (EC 4.4.1.4) y la sintasa del factor lacrimógeno (LFS). Los metabolitos secundarios de la aliina son:
- Sulfóxido de S-metil cisteína (MCSO, metiina, presente en la mayoría de las especies de Allium y en algunas brasicáceas)
- Sulfóxido de S-trans-prop-1-enil cisteína (PECSO o isoaliina; característica de la cebolla),
- Sulfóxido de S-propilcisteína (PCSO, propiina, en la cebolla y especies afines).
- Ajoenos (E y Z)
- Sulfóxido de tiopropanal
- Vinilditiinas
- Tioacroleína
- Sulfuros de alilo
Los estudios de la transformación de los intermediarios biosintéticos por marcaje, y de las mediciones de los compuestos de azufre en la cebolla y el ajo han proporcionado información para sugerir la ruta de biosíntesis.[10][11]

La fitoquímica de la subfamilia de las agapantóideas también ofrece particularidades. De hecho, todas las especies de Agapanthus producen fitoecdiesteroides, compuestos que están relacionados con la defensa de las plantas frente al ataque de insectos que se alimentan de sus hojas.[12][13] Además, se ha comprobado que los extractos de las especies de este género exhiben propiedades antifúngicas con una acción muy efectiva contra patógenos de las plantas.[14]
Además, y teniendo en cuenta que el color azul de las flores de los agapantos es único en la familia, se ha determinado las estructuras químicas de dos de las principales antocianinas que confieren tal color, las cuales representan los únicos ejemplos conocidos de pigmentos en los que el grupo acil aromático y el flavonoide están unidos covalentemente a la antocianina.[15]
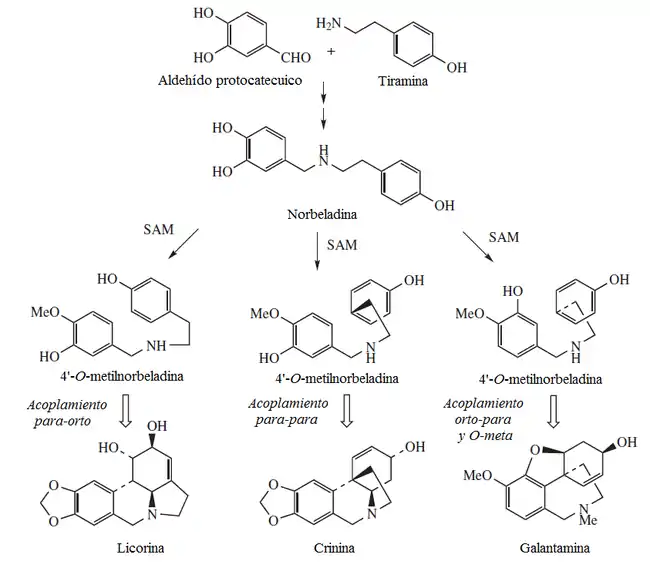
Las amarilidóideas no presentan sulfuro de alilo —característico de las alióideas— antocianidinas, ácido elágico, saponinas ni inulina. Los flavonoles pueden o no estar presentes.[4] Las amarilidóideas presentan un tipo único de alcaloides derivados de la tirosina llamados norbeladinas. Tales alcaloides son responsables de las propiedades ponzoñosas de un gran número de especies de la subfamilia. Se conocen más de 200 estructuras diferentes, de las cuales 79 fueron identificadas solamente en el género Narcissus.[3] La norbeladina es una amina formada por la reducción de la base de Schiff formada entre el aldehído protocatéquico y la tiramina. La norbeladina puede acoplar por radicales libres a los dos anillos aromáticos que presenta. Así, pueden formarse distintas estructuras de acuerdo al patrón de acoplamiento de los anillos.[16]
- a) Licorina
- b) Crinina
- c) Galantamina
Evolución y filogenia
La edad del grupo troncal de las amarilidáceas, o sea, la edad de las amarilidáceas y sus parientes extintos pertenecientes al mismo linaje, se ha estimado en 91 millones de años, y la del grupo corona, edad en la que se supone iniciaron su divergencia las amarilidáceas actuales, en 87 millones de años.[17]
Las agapantóideas, naturales de África, constituyen el clado más divergente de la familia. Las alióideas (con un representante de África y los restantes miembros de América y del hemisferio norte) son el clado hermano de las amarilidóideas, representadas en África, América, Asia y Europa. El cladograma que sintetiza las relaciones filogenéticas entre las subfamilias y tribus de las amarilidáceas se provee a continuación:[n. 1]
|
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Taxonomía
Historia taxonómica


Las amarilidáceas, como se hallan circunscriptas actualmente, constituyen un gran grupo de monocotiledóneas petaloideas que incluyen a las amarilidáceas propiamente dichas (como subfamilia Amarilidoideae), a los ajos y especies emparentadas (subfamilia Allioideae) y a las especies de agapantos (subfamilia Agapanthoideae).[28] La historia taxonómica de esta familia es bastante extensa y caracterizada por diversas interpretaciones y definiciones.
Las amarilidóideas representan uno de los elementos del grupo linneano Hexandra monogynia.[29] Desde 1764, los géneros de esta subfamilia han sido clasificados dentro de las liliáceas o como una familia separada, las amarilidáceas. Siete de los 51 géneros que Carlos Linneo dispuso en Hexandra monogynia han sido siempre incluidos dentro de una misma unidad taxonómica, como orden Amaryllidaceae por John Lindley[30] y William Herbert;[31] suborden Amarylleae por John Gilbert Baker,[32] familia Amaryllideae por Augustin Saint-Hilaire, Robert Brown[33] y John Hutchinson;[34] subfamilia Amaryllidoideae por Ferdinand Albin Pax[35] y Hamilton Paul Traub;[36] tribu Amarylleae en el Sistema Bentham & Hooker propuesto por George Bentham & Joseph Dalton Hooker;[37] o sección Narcissi por Michel Adanson[38] y Antoine-Laurent de Jussieu.[39] Robert Brown, en 1810, fue el primero en proponer que los géneros con ovario súpero debían ser excluidos de Amaryllidaceae. De hecho, tradicionalmente ambas familias —liliáceas y amarilidáceas— han sido separadas entre sí mediante la inclusión de las especies con ovario súpero en Liliaceae y aquellas con ovario ínfero en Amaryllidaceae.[40] Luego de más de un siglo, la clasificación de Hutchinson fue la primera re-circunscripción radical de Amaryllidaceae, en la que se definía el carácter que unificaba a todos los miembros de la familia: la presencia de inflorescencias en umbelas rodeadas de un involucro de una o más brácteas.[41]
Amaryllidaceae ha sido reconocida, con diferentes circunscripciones, por la mayoría de los sistemas de clasificación del siglo XX. En el sistema Takhtajan se reconoció a Amaryllidaceae en el sentido más estricto.[42] El sistema de Cronquist, sin embargo, incluía a esta familia dentro de una amplia definición de Liliaceae.[43]
La familia Liliaceae fue concebida por Antoine Laurent de Jussieu en 1789 y su definición era muy amplia y artificial: todas las especies de plantas con seis tépalos y gineceo de ovario súpero eran incluidas en esta familia. En un momento llegó a abarcar cerca de 300 géneros y 4500 especies que se incluían dentro del gran orden Liliales.[44][45] como las «monocotiledóneas petaloideas», un grupo caracterizado por flores con tépalos vistosos y sin almidón en el endosperma. Cronquist ubicó a la mayoría de las monocotiledóneas petaloideas con flores de seis estambres en un muy amplio (y polifilético) Liliaceae.[44] No obstante, con el tiempo se llegó a reconocer que, definida de esa forma, la familia de las liliáceas incluía un vasto y heterogéneo repertorio de géneros que no estaban relacionados filogenéticamente entre sí. Existieron varias propuestas para separar grupos pequeños de géneros en familias más homogéneas, pero ninguna de ellas fue ampliamente aceptada. En la década de 1980, en el contexto de una revisión general de la clasificación de las angiospermas, las liliáceas fueron sometidas a un escrutinio más intenso. Hacia fines de esa década, el Real Jardín Botánico de Kew, el Museo Británico y el Real Jardín Botánico de Edimburgo formaron un comité para examinar la posibilidad de separar a las liliáceas en subgrupos más homogéneos, al menos para la organización de sus herbarios. El comité recomendó que se utilicen 24 nuevas familias en lugar de la antigua y ampliamente definida Liliaceae.[46] En las últimas dos décadas los estudios de ADN y los datos morfológicos (particularmente aquellos relacionados con la morfología reproductiva) sumados a los análisis cladísticos, han permitido concluir que las monocotiledóneas petaloideas en realidad no pertenecen a una misma familia botánica sino que se distribuyen en dos órdenes diferentes: Asparagales y Liliales. La monofilia de estos órdenes está sustentada por análisis cladísticos basados en morfología, ADNr 18S y muchas otras secuencias de ADN. [47][48][49][50] [51]
Dos de las familias en las que se dividió a la antigua familia de las liliáceas son las aliáceas y las agapantáceas, ambas monocotiledóneas petaloideas que actualmente se incluyen en el orden Asparagales como subfamilias de las amarilidáceas. En el sistema Dahlgren de 1985, ya se había postulado la existencia de dos órdenes separados para las monocotiledóneas petaloideas. De hecho, había reconocido a Amaryllidaceae y a Alliaceae como dos familias separadas dentro del orden Asparagales, un orden de 31 familias que ha evolucionado muchos caracteres en paralelo con el orden Liliales.[52] Uno de los caracteres más importantes y consistentes para separar ambos órdenes es la presencia de fitomelaninas en la cubierta de las semillas de Asparagales, un carácter puesto ya de manifiesto por Jacques Huber en 1969.[41]
Por su parte, el género Agapanthus fue establecido por L'Heritier en 1788. Desde su creación, la inclusión del género en una determinada familia ha sido materia de debate. Durante décadas Agapanthus fue dispuesto en la familia de las liliáceas, como por ejemplo, en el Sistema de Cronquist. Luego fue trasladado a la familia Amaryllidaceae y más tarde nuevamente trasladado a la familia Alliaceae. Durante algunos años se lo dispuso en su propia familia, Agapanthaceae, un rango que fue muy discutido.[53]
El Grupo para la Clasificación Filogenética de las Angiospermas (APG) en su trabajo de 1998[54] aceptó a Agapanthaceae, Alliaceae y Amaryllidaceae como tres familias independientes dentro de orden Asparagales. En el trabajo del APG II (2003) el reconocimiento de Agapanthaceae y Amaryllidaceae era opcional, ambas podían también ser incluidas dentro de la familia Alliaceae sensu lato.[55] Las alióideas están estrechamente emparentadas con las amarilidóideas y con las agapantóideas. De hecho, los tres grupos son hierbas bulbosas con inflorescencias terminales umbeladas, que tienen por debajo brácteas espatáceas y nacen en un conspicuo escapo, todas estas características probablemente son sinapomórficas. Más aún, los análisis cladísticos sostienen la estrecha relación entre ellas.[18][56][57][19][20] En la publicación de 2009, el sistema APG III reconfirmó el criterio de considerar a las tres familias como parte de una gran circunscripción de Alliaceae.[58] No obstante, en vez de conservarse el nombre Alliaceae, que tiene preeminencia, se aceptó la propuesta del taxónomo y especialista en plantas bulbosas Alan W. Meerow de utilizar para este grupo el nombre Amaryllidaceae.[59]
Clasificación
- 1.- Agapanthoideae Endl.





Las agapantóideas incluyen un solo género, Agapanthus, el cual es muy variable, a pesar de que todas las especies tienen una apariencia similar, con gruesos rizomas, hojas en forma de largas tiras o bandas e inflorescencias en umbelas en el extremo de un escapo más alto que las hojas. Como resultado de varios estudios que incluyen caracteres tales como el contenido de ADN nuclear, color y vitalidad del polen y estudios morfológicos, muchas especies de han reducido a sinonimia y otras se han categorizado como subespecies, por lo que hoy se considera que hay solo dos especies perennifolias —A. africanus y A. praecox— y cuatro especies caducifolias —A. campanulatus, A. caulescens, A. coddii y A. inapertus— lo que hace un total de 6 especies, muchas de las cuales presentan una o más subespecies.[60]
- 2.- Allioideae Herb.
La subfamilia Allioideae comprende aproximadamente 13 géneros y unas 600 especies. Los géneros más representados son Allium (550 especies), Ipheion (25 especies) y Tulbaghia (24 especies). La subfamilia fue dividida en 3 tribus, sobre la base de la distribución geográfica, los caracteres morfológicos, citológicos y moleculares. Las tres tribus y sus características más distintivas son:[1]
- Allieae, comprende un solo género, Allium, distribuido en zonas templadas del Hemisferio Norte. Los bulbos no presentan almidón, las hojas son más o menos unifaciales, la corola es connada en su parte basal, no presentan corona o paraperigonio. Los estambres son connados basalmente o adnatos a la corola y presentan filamentos alados. Las células del tapete de las anteras son mononucleares. Los óvulos, en número de 2 a 14 por carpelo, son epítropos. El estilo es sólido y ginobásico. El endosperma es celular y el embrión curvo. El número cromosómico básico más frecuente es x = 8, no obstante, se han informado especies con x = 7 y x = 9. La poliploidía y la apomixis son bastante frecuentes.
- Tulbaghieae, comprende también un solo género, Tulbaghia, que se distribuye en Sudáfrica. Presenta plantas rizomatosas, con cortas láminas foliares. Las flores presentan brácteas, la corola es fuertemente connada y exhiben una corona masiva. Los estambres son sésiles (no poseen filamento) y se hallan unidos a la corola. Presentan de dos a muchos óvulos por carpelo y las semillas son más o menos aplanadas. El número cromosómico básico es x = 6.
- Gilliesieae: es la más variable de las tres tribus, con 10 u 11 géneros que se distribuyen desde el sur de Estados Unidos y México hasta Argentina y Chile. Las flores pueden o no presentar corona, el androceo está compuesto por 2 o 3 estambres, pueden presentar estaminodios. Muestran de dos a muchos óvulos por carpelo. El embrión es corto y el número cromosómico más frecuente es x = 4. Gilliesia presenta flores fuertemente zigomorfas, con solo 2 estambres, que mimetizan insectos para atraerlos y que éstos faciliten la polinización. Schickendantziella, por otro lado, presenta solo 3 tépalos. Finalmente, Nothoscordum tiene estilos sólidos. Los restantes géneros que se incluyen en esta tribu son Ancrumia, Caloscordum, Erinna, Garaventia, Gethyum, Ipheion, Leucocoryne, Miersia, Milula, Muilla, Nectaroscordum, Solaria, Speea, Trichlora, Tristagma y Zoellnerallium.
- 3.- Amaryllidoideae Burnett
La subfamilia de las amarilidóideas comprende unas 800 especies ordenadas en 13 tribus las que, a su vez, pueden agruparse en clados según sugiere el análisis filogenético de las mismas basado en secuencias de ADN plastídico.[61] Tales clados son congruentes con la distribución geográfica de las tribus.[62] Las tribus de Amaryllidaceae, el número de géneros que componen cada una, su número cromosómico básico y algunas características de cada una, se listan a continuación:[41]
- Amaryllideae (11 géneros, x = 10 y 11). Esta tribu está caracterizada por la presencia de fibras cartilaginosas en las hojas, escapos con esclerénquima, polen bisulcado con exina espinulosa (los granos de polen son monosulcados y con exina reticulada en los restantes géneros de la familia) y por los óvulos unitégmicos. Las semillas de los miembros de esta tribu son verdes y carnosas y no presentan un período de dormición o latencia, por lo que germinan rápidamente, incluso sobre la misma planta madre. Varios géneros populares en jardinería pertenecen a esta tribu, enteramente africana, tales como Amaryllis, Crinum y Nerine.[63]
- Cyrtantheae (1 género, x = 8). Consiste en un solo género endémico de África, Cyrtanthus, el cual ha sido separado de los restantes miembros de Haemantheae para disponerlo en su propia tribu. Comparte con algunos géneros de Haemantheae el mismo número cromosómico (x = 8) y la distribución estrictamente africana, pero se distingue por sus cápsulas loculicidas. Las semillas chatas, aladas y con fitomelaninas distinguen a Cyrtantheae de todas las restantes tribus africanas. Es, por otro lado, la tribu más diversa en cuanto a morfología floral.
- Haemantheae (6 géneros, x = 6, 8, 9, 11 y 12). Esta tribu africana comprende géneros que presentan fruto en forma de baya carnosa y con semillas de viabilidad breve. En general, no tienen bulbo sino raíces fasciculadas, como por ejemplo Clivia, Scadoxus y Cryptostephanus[64]
- Calostemmateae (2 géneros, x = 10). Esta tribu consiste en dos géneros, Proiphys y Calostemma, que constituyen un componente de Australasia aislado del resto de la familia.
- Lycoridae (2 géneros, x = 11). Constituye el linaje de Asia central y oriental de la familia. Los dos géneros que la componen (Lycoris y Ungernia) están estrechamente relacionados filogenéticamente con la tribu Pancratiae.
- Pancratieae (2 géneros, x = 11). Es una tribu distribuida en el Viejo Mundo, con una posición filogenética bastante controvertida. Pancratium es el miembro de la tribu con una distribución más amplia.
- Narcisseae (2 géneros, x = 7, 10 y 11). Caracterizada por un escapo sólido y brácteas espatáceas fusionadas en un tubo. Narcissus presenta paraperigonio, el cual está ausente en Sternbergia.
- Galantheae (5 géneros, x = 7, 8, 9, 11 y 12). Es una tribu muy relacionada con la anterior y, de hecho, para algunos taxónomos debe considerarse una subtribu de Narcisseae. Galanthus y Leucojum son dos géneros popularmente conocidos en jardinería.
- Hippeastreae (ca. 10 géneros, x = 6, 8, 9, 10, 11 y 12). Es una tribu neotropical originaria de América. Está caracterizada por un escapo hueco y cierto desarrollo del paraperigonio. Hippeastrum, Griffinia, Habranthus, y Sprekelia son algunos de los géneros de esta tribu.
- Eucharideae (4 géneros, x = 23). Tribu americana con representantes originarios del este de Los Andes. Presentan hojas largas y pecioladas. Los géneros que constituyen esta tribu son Eucharis, Caliphruria, Urceolina y Plagiolirion.
- Stenomesseae (8 géneros, x = 23). Es una tribu andina, caracterizada por un número cromosómico 2n = 46 y por un paraperigonio bien desarrollado en la mayoría de los géneros. Stenomesson es uno de los representantes de la tribu.
- Hymenocallideae (3 géneros, x = 23). Tribu americana constituida por los géneros Ismene, Hymenocallis y Leptochiton, los que presentan una corona estaminal muy desarrollada.
- Eustephieae (3 géneros, x = 23). Esta tribu representa un clado de los Andes centrales y australes. Los géneros Eustephia, Hieronymiella y Chlidanthus presentan semillas secas, chatas y discoideas.
Importancia económica y cultural
Como hortalizas y condimento
El género económicamente más importante es Allium, que incluye algunas de las plantas comestibles más utilizadas, tales como la cebolla (Allium cepa), el ajo (A. sativum y también A. scordoprasum), la chalota, echalote o escalonia (Allium ascalonicum), el puerro (A. ampeloprasum), el puerro salvaje o rampa (Allium tricoccum) y varios condimentos como la cebolleta (A. schoenoprasum). Se consumen tanto los bulbos, frescos o cocidos, como las hojas.
Como plantas medicinales
Los representantes de las alóideas son ligeramente antisépticos, y muchos son utilizados en medicina. Los compuestos organosulfurados de las especies de esta subfamilia tienen propiedades antioxidantes, antibióticas, anticarcinogénicas, antiteratogénicas, estimulantes del sistema inmune y protectoras de la función hepática.[65][66] Varios estudios basados en investigaciones clínicas y epidemiológicas indican que el consumo regular de ajo puede disminuir el riesgo de cáncer y de enfermedades cardiovasculares. Además, el ajo posee fuertes propiedades antisépticas.[67]
Como plantas ornamentales

Numerosas especies de Allium —los denominados «ajos ornamentales»— se utilizan en jardinería para embellecer parques y jardines. Son fáciles de cultivar si bien no son muy populares, tal vez por el olor aliáceo que despiden. Sus inflorescencias tienen larga vida en agua, por lo que también son útiles como flor cortada. Además, las inflorescencias secas se utilizan para decoración hogareña. La diversidad que presentan estas especies ornamentales, tanto en altura (desde 20 a 120 cm), como en forma y color del follaje (desde hojas finas hasta muy anchas, verde oscuro hasta verde azulado) y color de las flores (amarillo, blanco, azul, púrpura y rosado) las hace sumamente versátiles para ubicar en distintos tipos de jardines. Presentan dos tipos básicos de inflorescencias: en esferas grandes o en pequeños grupos de flores erectas o decumbentes. Entre las primeras se hallan Allium giganteum, A. albopilosum, A. sphaerocephalon, A. aflatunense y A. karataviense. En el segundo tipo de inflorescencia se hallan A. narcissiflorum, A. triquetrum, A. beesianum y la popular A. moly.[3][68] Ipheion uniflorum, conocida con el nombre de «estrellita», es una pequeña bulbosa de no más de 15 cm de altura que se utiliza en roqueros y para naturalizar debajo de árboles. Florece en primavera y provee una abundante cantidad de flores con forma de estrella. El color más común es el blanco, pero hay cultivares de color azulado (‘Wisley Blue’) y azul profundo (‘Rolf Fiedler’).[3][68] Tulbaghia violacea («ajo de sociedad») es el representante del género mejor conocido. Tiene una altura de 40 a 60 cm, presenta flores tubulares de color violeta claro, follaje siempreverde y florece durante todo el verano.
Otros géneros de las alióideas con representantes ornamentales son Leucocoryne y Gilliesia.[3][68]
Las especies y cultivares de agapantos también se utilizan como plantas ornamentales en parques y jardines. Son especies sumamente adaptables, rústicas y de fácil cultivo, razón por la que se las utiliza mucho en parques y jardines de bajo mantenimiento. La época de floración es fines de primavera y verano. Si bien toleran cualquier tipo de suelo, se logran excelentes ejemplares usando un suelo profundo y fértil. Pueden prosperar bien tanto a pleno sol como a media sombra. Requieren un abundante riego en primavera y verano, más espaciado durante el invierno. Se multiplican por semillas o por división de las matas en otoño.[3][68] Muchas de las especies de amarilidóideas son populares como plantas ornamentales en parques y jardines. Así, varias especies y cultivares de los géneros Crinum, Nerine, Amaryllis, Leucojum, e Hippeastrum se cultivan con ese objeto desde hace siglos. Una mención aparte merece el caso de Narcissus, cultivado en varias partes del mundo como ornamental en jardines y también para flor de corte.
También se cultivan Eucharis, Galanthus, Haemanthus, Hymenocallis, Lycoris y Cyrtanthus. Otros géneros, tales como Habranthus y Zephyranthes, no son tan conocidos, pero su cultivo y mejoramiento ya se ha establecido en varias partes del mundo.[69][70]
Plantas tóxicas
Muchos taxones, particularmente dentro de las amarilidóideas, son utilizados por pueblos indígenas por sus usos medicinales, como saborizantes, psicotrópicos, y otros propósitos.[71] Así, Boophone disticha es conocido en Sudáfrica como gifbol («bulbo venenoso») debido a su legendaria toxicidad.[72] Esta propiedad era utilizada por los aborígenes de África Austral para envenenar las puntas de sus flechas al realizar cacerías.[72] La mayoría, sino todas, las amarilidóideas son tóxicas para los humanos. A modo de ejemplo, Amaryllis belladonna presenta un alcaloide en el bulbo llamado licorina, el cual afecta al corazón y si los bulbos son ingeridos en cantidad pueden ser mortales.[72] Los géneros Ammocharis, Boophone, Brunsvigia, Clivia, Crinum, Cyrtanthus, Galanthus, Haemanthus, Hymenocallis, Narcissus, Nerine, Scadoxus y Zephyranthes han sido citados como tóxicos para los humanos debido a la presencia de alcaloides en los bulbos u otras partes de la planta.[73]
Véase también
Notas
- La topología general del cladograma está basada en los trabajos de filogenia de Asparagales,[18][19][20] las relaciones dentro de Allioideae se basan en el trabajo de filogenia y clasificación de Allium,[21] mientras que las interrelaciones dentro de las amarilidóideas se basan en los trabajo de Alan W. Meerow y Dierdre Snijman.[22][23][24][25][26][27]
Referencias
- Stevens, P. F. (2001 en adelante). «Alliaceae». Angiosperm Phylogeny Website. Version 7, mayo 2006 (en inglés). Consultado el 28 de abril de 2008.
- Dimitri, M. 1987. Enciclopedia Argentina de Agricultura y Jardinería. Tomo I. Descripción de plantas cultivadas. Editorial ACME S.A.C.I., Buenos Aires.
- Rosella Rossi. 1990. Guía de Bulbos. Grijalbo. Barcelona.
- Amaryllidaceae. In: L. Watson & M.J. Dallwitz (1992 en adelante). The families of flowering plants: descriptions, illustrations, identification, information retrieval. Version: 27th April 2006. http://delta-intkey.com Archivado el 3 de enero de 2007 en Wayback Machine.. Fecha de acceso: 10 de agosto del 2007
- Rahn, K. (1998). «Alliaceae». En Kubitzki, K., ed. The families and genera of vascular plants, vol 3, Monocotyledons: Lilianae (except Orchidaceae). Berlin: Springer-Verlag. pp. 70-78.
- Watson, L.; Dallwitz, M. J. «Alliaceae». The families of flowering plants: descriptions, illustrations, identification, and information retrieval. Version: 1st June 2007. (en inglés). Consultado el 28 de abril de 2008.
- Zonneveld, B.J.M. y Duncan, G.D. (2003). «Taxonomic implications of genome size and pollen colour and vitality for species of Agapanthus L’Héritier (Agapanthaceae)». Plant Systematics and Evolution 241 (1): 115-123. doi:10.1007/s00606-003-0038-6.
- Van Damme EJM, Allen AK, Peumans WJ (1987) Isolation and characterization of a lectin with exclusive specificity towards mannose from snowdrop (Galanthus nivalis) bulbs. FEBS Lett 215: 140-144
- Van Damme EJM, Allen AK, Peumans WJ (1988) Related mannose-specific lectins from different species of the family Amaryllidaceae. Physiol Plant 73: 52-57
- Paul M. Dewick (2009). Medicinal natural products: a biosynthetic approach. John Wiley and Sons. ISBN 9780470741689.
- Jones MG, Hughes J, Tregova A, Milne J, Tomsett AB, Collin HA (2004). «Biosynthesis of the flavour precursors of onion and garlic». J Exp Bot. 55 (404): 1903-1918.
- Savchenko, T.; Whiting, P.; Sarker, S.D.; Dinan, L. (1997), «Phytoecdysteroids in the Genus Agapanthus (Alliaceae)», Biochemical Systematics and Ecology 25 (7): 623-629, doi:10.1016/S0305-1978(97)00056-2.
- Dinan, L.; Savchenko, T.; Ri Whiting, P.; Sarker{textordmasculine (1999), «Plant natural products as insect steroid receptor agonists and antagonists», Pesticide Science 55: 331-335, doi:10.1002/(SICI)1096-9063(199903)55:3<331::AID-PS889>3.0.CO;2-1.
- Tegegne, G.; Pretorius, J.C.; Swart, W.J. (2008), «Antifungal properties of Agapanthus africanus L. Extracts against plant pathogens», Crop Protection 27 (7): 1052-1060, doi:10.1016/j.cropro.2007.12.007.
- Bloor, S.J.; Falshaw, R. (2000), «Covalently linked anthocyanin-flavonol pigments from blue Agapanthus flowers», Phytochemistry 53 (5): 575-579, doi:10.1016/S0031-9422(99)00572-5.
- Ghosal, S. et al. (1985) Phytochemistry, 24, 2141.
- Stevens, P. F. (2001 en adelante). «Amaryllidaceae». Angiosperm Phylogeny Website, versión 10. Última actualización de la sección: 21 de junio de 2008 (en inglés). University of Missouri, St Louis, and Missouri Botanical Garden. Consultado el 15 de diciembre de 2009.
- Chase, M. W.; Duvall, M. R., Hills, H. G., Conran, J. G., Cox, A. V., Eguiarte, L. E., Hartwell, J., Fay, M. F., Caddick, L. R., Cameron, K. M., y Hoot, S. (1995). «Molecular systematics of Lilianae.». En Rudall, P. J., Cribb, P. J., Cutler, D. F., ed. Monocotyledons: Systematics and evolution. (Royal Botanic Gardens edición). Kew. pp. 109-137.
- Fay, M. F. (2000). «Phylogenetic studies of Asparagales based on four plastid DNA regions.». En K. L. Wilson y D. A. Morrison, ed. Monocots: Systematics and evolution. (Royal Botanic Gardens edición). Kollingwood, Australia: CSIRO. pp. 360-371.
- Pires, J. C.; Sytsma, K. J. (2002). «:A phylogenetic evaluation of a biosystematic framework: Brodiaea and related petaloid monocots (Themidaceae).». Amer. J. Bot. (89): 1342-1359.
- Friesen, N., Fritsch, R. y Blattner, F. (2006). «Phylogeny and new intrageneric classification of Allium (Alliaceae) based on nuclear ribosomal DNA its sequences». Aliso 22: 372-395.
- Meerow, A. W. (1995). «Towards a phylogeny of Amaryllidaceae.». En Rudall, P. J., Cribb, P. J., Cutler, D. F., y Humphries, C. J., ed. Monocotyledons: Systematics and evolution. (Royal Botanic Gardens edición). Kew. pp. 169-179.
- Meerow, A.; Fay, M. & Chase, M. Guy, Ch. y Q-B. Li. 1999a. The New Phylogeny of the Amaryllidaceae. Herbertia 54:180-203.
- Meerow, A. W.; Fay, M. F., Chase, M. W., Guy, C. I., Li, Q-B., Snijman, D., Yang, S-L. (2000a). «Phylogeny of Amaryllidaceae: molecules and morphology.». En Wilson, K. L. y Morrison, D. A., ed. Monocots: Systematics and evolution. (CSIRO Publ. edición). Collingwood, Australia. pp. 372-386.
- Meerow, A. W.; Guy, C. L., Li, Q-B. y Yang, S-L. (2000b). «Phylogeny of the American Amaryllidaceae based on nrDNA ITS sequences». Syst. Bot. (25): 708-726.
- Meerow, A. W.; Snijman, D. A. (1998). «Amaryllidaceae.». En Kubitzki, K., ed. The families and genera of vascular plants, vol 3, Monocotyledons: Lilianae (except Orchidaceae). (Royal Botanic Gardens edición). Berlin: Springer-Verlag. pp. 83-110.
- Meerow, A. W.; Suijman, D. A. (2006). «The never-ending story: multigene approaches to the phylogeny of Amaryllidaceae» (PDF). Aliso (22): 355-366. Archivado desde el original el 3 de marzo de 2012.
- Chase, M.; Reveal, J.; Fay M. 2009. A subfamilial classification for the expanded asparagalean families Amaryllidaceae, Asparagaceae and Xanthorrhoeaceae. Botanical Journal of the Linnean Society 161: 132-136.
- Linnaeus, C. 1764. Species Plantarum, 3rd ed. Stockholm.
- Lindley, J. 1836. A natural system of botany. Longman, London.
- Herbert, W. (1837). Amaryllidaceae: preceded by an attempt to arrange the monocotyledonous orders, and followed by a treatise on cross-bred vegetables, and supplement. Londres: James Ridgway & Sons.
- Baker, J. 1888. Handbook of the Amaryllideae, including the Alstroemerieae and Agaveae. George Bell & Sons.
- Brown, R. 1810. Prodromus floræ Novæ Hollandiæ et Insulæ Van-Diemen
- Hutchinson, J. 1959. The Families of Flowering Plants. Ed. 2. 2 Vols. Clarendon Press, Oxford.
- PAX, F. 1888. Amaryllidaceae, pp. 97–124. In A. Engler and K. Prantl [eds.], Die natûrlichen pflanzenfamilien, Vol. 2. W. Engelmann, Leipzig, Germany
- Traub, H.P. 1963. The genera of Amaryllidaceae. American Plant Life Society; 1.ª ed. 86 pp
- G. Bentham & J.D. Hooker (three volumes, 1862–1883). Genera plantarum ad exemplaria imprimis in herbariis kewensibus servata definita.
- Adanson, M. 1763. Familles des plantes. Vinvent, París.
- de Jussieu. A.L. 1789. Genera Plantarum. Herissant, París.
- Lawrence, G. H. M. (1951). Taxonomy of vascular plants. (Macmillan edición). New York.
- Meerow, A.; Fay, M. y Chase, M. Guy, Ch. y Q-B. Li. The New Phylogeny of the Amaryllidaceae. Herbertia 54:180-203; 1999
- Takhtajan, A. 1969. Flowering Plants. Origin and Dispersal. Oliver and Boyd. London.
- Cronquist, A. 1988. The evolution and classification of flowering plants. 2.ª edición, New York Botanical Garden, New York.
- Cronquist, A. (1981). An integrated system of classification of flowering plants. Nueva York: Columbia University Press.
- Thorne, R. F. (1992). «Classification and geography of the flowering plants». Bot. Rev. (58): 225-348.
- Mathew, Brian (1989). Splitting the Liliaceae. The Plantsman 11 (2): 89.
- Chase, M. W.; Fay, M. F.; Devey, D. S.; Maurin, O; Rønsted, N; Davies, T. J; Pillon, Y; Petersen, G; Seberg, O; Tamura, M. N.; Lange, Conny Bruun Asmussen (Faggruppe Botanik); Hilu, K; Borsch, T; Davis, J. I; Stevenson, D. W.; Pires, J. C.; Givnish, T. J.; Sytsma, K. J.; McPherson, M. A.; Graham, S. W.; Rai, H. S. (2006). «Multigene analyses of monocot relationships: a summary» (pdf). Aliso (22): 63-75. ISSN: 00656275. (enlace roto disponible en Internet Archive; véase el historial, la primera versión y la última).
- Davis, J. I.; Stevenson, D. W.; Petersen, G.; Seberg, O.; Campbell, L. M.; Freudenstein, J. V.; Goldman, D. H.; Hardy, C. R.; Michelangeli, F. A.; Simmons, M. P.; Specht, C. D.; Vergara-Silva, F.; Gandolfo, M. (2004). «A phylogeny of the monocots, as inferred from rbcL and atpA sequence variation, and a comparison of methods for calculating jacknife and bootstrap values.». Syst. Bot. (29): 467-510.
- Graham, S. W.; Zgurski, J. M., McPherson, M. A., Cherniawsky, D. M., Saarela, J. M., Horne, E. S. C., Smith, S. Y., Wong, W. A., O'Brien, H. E., Biron, V. L., Pires, J. C., Olmstead, R. G., Chase, M. W., y Rai, H. S. (2006). «Robust inference of monocot deep phylogeny using an expanded multigene plastid data set.» (pdf). Aliso (22): 3-21.
- Hilu, K.; Borsch, T., Muller, K., Soltis, D. E., Soltis, P. S., Savolainen, V., Chase, M. W., Powell, M. P., Alice, L. A., Evans, R., Sauquet, H., Neinhuis, C., Slotta, T. A. B., Rohwer, J. G., Campbell, C. S., y Chatrou, L. W. (2003). «Angiosperm phylogeny based on matK sequence information.». American J. Bot. (90): 1758-1766.
- Källersjö M; JS Farris, MW Chase, B Bremer, MF Fay, CJ Humphries, G Petersen, O Seberg y K Bremer (1998). «Simultaneous parsimony jacknife analysis of 2538 rbcL DNA sequences reveals support for major clades of green plants, land plants, and flowering plants.». Pl. Syst. Evol. (213): 259-287.
- Dahlgren RMT, Clifford HT, Yeo PF. 1985 The families of the monocotyledons. Structure, evolution, and taxonomy. Berlin, etc.: Springer-Verlag 520pp
- Meerow, A.W. (2000), «The New Phylogeny of the Lilioid Monocotyledons», VIII International Symposium on Flowerbulbs 570: 31-45, archivado desde el original el 3 de marzo de 2016, consultado el 29 de enero de 2010.
- Angiosperm Phylogeny Group. (1998). «An ordinal classification for the families of flowering plants» (PDF). Ann. Missouri Bot. Gard. 85: 531-553. Archivado desde el original el 8 de junio de 2011. Consultado el 26 de diciembre de 2008.
- APG II. 2003. An Update of the Angiosperm Phylogeny Group Classification for the orders and families of flowering plants: APG II. (enlace roto disponible en Internet Archive; véase el historial, la primera versión y la última). Botanical Journal of the Linnean Society. 141, 399-436
- Chase, M.W.; Stevenson, D. W., Wilkin, P., y Rudall, P. J. (1995b). «Monocot systematics: a combined analysis.». En Rudall, P. J., Cribb, P. J., Cutler, D. F., ed. Monocotyledons: Systematics and evolution. (Royal Botanic Gardens edición). Kew. pp. 685-730.
- Fay, M. y Chase, M. 1996. Resurrection of Themidaceae for the Brodiaea alliance, and Recircumscription of Alliaceae, Amaryllidaceae and Agapanthoideae. Taxon 45: 441-451
- THE ANGIOSPERM PHYLOGENY GROUP (Birgitta Bremer, Kåre Bremer, Mark W. Chase, Michael F. Fay, James L. Reveal, Douglas E. Soltis, Pamela S. Soltis and Peter F. Stevens, Arne A. Anderberg, Michael J. Moore, Richard G. Olmstead, Paula J. Rudall, Kenneth J. Sytsma, David C. Tank, Kenneth Wurdack, Jenny Q.-Y. Xiang & Sue Zmarzty). 2009. An update of the Angiosperm Phylogeny Group classification for the orders and families of flowering plants: APG III. Botanical Journal of the Linnean Society 161, 105–121.
- Meerow, A.W., Reveal, J.L., Snijman, D.A., Dutilh, J.H. Superconservation proposal to conserve Amaryllidaceae (1805) over Alliaceae (1797). Taxon 56: 1299-1300. 2007
- Zonneveld, B.J.M.; Duncan, G.D. (2003), «Taxonomic implications of genome size and pollen colour and vitality for species of Agapanthus L’Héritier (Agapanthaceae)», Plant Systematics and Evolution 241 (1): 115-123, doi:10.1007/s00606-003-0038-6.
- Meerow, A.W. 1995. Towards a phylogeny of the Amaryllidaceae. En P. J. Rudall, P. J. Cribb, D. F. Cutler, and C. J. Humphries (editores), Monocotyledons: systematics and evolution, 169-179. Royal Botanic Gardens, Kew.
- Meerow, A.; Fay, M.; Chae, M. Guy, Ch. & Q-B. Li, Zaman, F. y Chase, M. (1999). «Systematics of Amaryllidaceae based on cladistic analysis of plastid rbcL and trnL-F sequence data». American Journal of Botany 86 (9): 1325-1345. Archivado desde el original el 9 de enero de 2009. Consultado el 8 de febrero de 2010.
- Snijman, D. A. and H. P. Linder. 1996. Phylogenetic relationships, seed characters, and dispersal system evolution in Amaryllideae (Amaryllidaceae). Annals of the Missouri Botanical Garden 83: 362-386.
- Meerow, A. W., y Clayton, J. R. (2004). «Generic relationships among the baccate-fruited Amaryllidaceae (tribe Haemantheae) inferred from plastid and nuclear non-coding DNA sequences». Plant Systematics and Evolution 244: 141-155. doi:10.1007/s00606-003-0085-z. Archivado desde el original el 3 de marzo de 2012.
- Tapiero, H.; Danyelle M. Townsend & Kenneth D. Tew (2004). «:Organosulfur compounds from alliaceae in the prevention of human pathologies». Biomedecine & Pharmacotherapy 58 (3): 183-193. Consultado el 25 de febrero de 2008.
- Evidente, A.; A. Kornienko. 2009. Anticancer evaluation of structurally diverse Amaryllidaceae alkaloids and their synthetic derivatives. Phytochemistry Reviews Volume 8, 2:449-459.
- Baasin'ska, B., Kulasek, G. (2004). «:Garlic and its impact on animal and human health». Medycyna Weterynaryjna 60 (11): 1151-1155. Consultado el 25 de febrero de 2008. (enlace roto disponible en Internet Archive; véase el historial, la primera versión y la última).
- Hessayon, D.G. The Bulb Expert. Transworld Publishers Ltd. Londres, 1999
- Hessayon, D.G. The Bulb Expert. Transworld Publishers Ltd. Londres, 1999.
- Taylor, P. 1996. Gardening with bulbs, a practical and inspirational guide. Pavillion Books Ltd., Londres.
- C. A. M. Louw, T. J. C. Regnier, L. Korsten. 2002. Medicinal bulbous plants of South Africa and their traditional relevance in the control of infectious diseases. Journal of Ethnopharmacology, Volume 82, Issues 2-3, 147-154
- Amaryllidaceae. Toxicité des Amaryllidaceae Archivado el 24 de abril de 2013 en Wayback Machine.. Consultado el 10 de enero de 2009.
- Munday, J. 1988. Poisonous Plants in South African Gardens and Parks. A Field Guide. Delta Books, Craighall, Johannesburg.
Bibliografía
- Chase, M.; Reveal, J.; Fay M. 2009. A subfamilial classification for the expanded asparagalean families Amaryllidaceae, Asparagaceae and Xanthorrhoeaceae. Botanical Journal of the Linnean Society 161: 132-136.
- Stevens, P. F. (2001 en adelante). «Alliaceae». Angiosperm Phylogeny Website (Versión 9, junio del 2008, y actualizado desde entonces) (en inglés). Consultado el 15 de enero de 2009.
Enlaces externos
 Wikimedia Commons alberga una categoría multimedia sobre Amaryllidaceae.
Wikimedia Commons alberga una categoría multimedia sobre Amaryllidaceae.- Acceso a imágenes de la mayoría de los géneros de Amaryllidaceae en "Bulbsociety"
- Información e imágenes de varios géneros de Amaryllidaceae en "Pacificbulbsociety"
- Amaryllidaceae chilenos, por Chileflora
- Amaryllidaceae en BoDD – Botanical Dermatology Database
| Control de autoridades |
|
|---|
 Datos: Q155848
Datos: Q155848- Multimedia: Amaryllidaceae / Q155848
 Especies: Amaryllidaceae
Especies: Amaryllidaceae