Anthocerotophyta
Anthocerotophyta es una división de plantas clasificada tradicionalmente entre las briofitas. En español carecen de nombre vulgar, aunque se suele utilizar derivados del nombre científico: antoceros,[1] antocerotas,[2], antocerotes, antocerotófitos o antocerotofitas. El nombre se refiere a la estructura alargada en forma de cuerno, que es el esporofito. Como en los musgos y las hepáticas, el cuerpo vegetal aplanado y verde de un antocero es el gametofito.
| Anthocerotophyta | ||
|---|---|---|
 | ||
| Taxonomía | ||
| Reino: | Plantae | |
| Superdivisión: | Bryophyta sensu lato | |
| División: |
Anthocerotophyta o Anthocerophyta Rothm. ex Stotl. & Crand.-Stotl., 1977 | |
| Clases y órdenes | ||
| ||
Los antoceros se pueden encontrar en todo el mundo, aunque tienden a crecer solo en lugares húmedos. Algunas especies crecen en grandes cantidades como pequeñas malezas en el suelo de jardines y campos cultivados. Se pueden encontrar grandes especies tropicales y subtropicales creciendo en la corteza de los árboles.
Reúne el grupo un total de unas 200-250 especies, repartidas en catorce géneros y cinco familias. Se encuentran distribuidas por todo el mundo, en zonas húmedas o con aguas rezumantes, a veces en grandes extensiones, pero usualmente son raras y difíciles de encontrar, siendo más comunes en zonas cálidas y muy raras en climas fríos.
Es un grupo empobrecido en especies debido fundamentalmente a su antigüedad y a la probable extinción de sus representantes, que actualmente tan solo constituyen los relictos de una radiación adaptativa temprana en la evolución de las plantas.
Presentan alternancia de generaciones, con un gametófito haploide y un esporófito diploide perfectamente diferenciados, pero indisolublemente unidos, un carácter propio de los embriófitos.
Gametófito
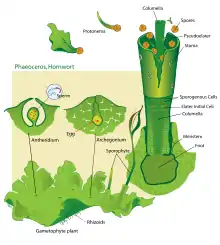
Se forma a partir del desarrollo de una espora haploide. Esta da lugar directamente un gametófito taloso, similar al de algunas hepáticas (con las que puede llegar a confundirse), sin desarrollar previamente un protonema. El gametófito es arrosetado y de ramificación dicotómica, si bien ésta puede quedar obscurecida durante el desarrollo. El talo puede presentar nervios o tener una banda central pluriestratificada, y presentar laminillas dorsales o prolongaciones setiformes, sus bordes a veces están lobulados. Varias especies desarrollan propágulos marginales que, al desprenderse, dan lugar directamente a nuevos talos. Los talos están formados por células parenquimáticas, no están diferenciados en tejidos y por lo tanto no existe ningún tejido conductor, si bien pueden aparecer cavidades mucilaginosas, abiertas al exterior mediante unos poros no estomáticos (a veces ausentes), que pueden ser colonizadas por cianobacterias del género Nostoc, pero no existen células mucilaginosas especializadas. Cada célula contiene un solo cloroplasto discoidal grande que rodea un pirenoide central (como ocurre en algunas algas verdes), con algunas excepciones. El talo de muchas especies presenta unas tuberosidades perennes, usualmente cerca de los bordes, que están rodeadas por células suberosas y cuyas células internas son ricas en sustancias de reserva (proteínas, lípidos), como ocurre también en algunas hepáticas talosas. En la cara ventral se encuentran rizoides unicelulares hialinos de pared lisa.
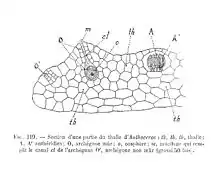
Los órganos sexuales se encuentran en la cara superior del talo, hundidos en cámaras más o menos profundas del mismo en su madurez. Se originan a partir de una célula subepidérmica y en su crecimiento, los anteridios (órganos masculinos) quedan expuestos al exterior al producirse la ruptura de las células epidérmicas del talo que recubren las cámaras anteridiales. En cada una de estas cámaras anteridiales se desarrollan uno a varios anteridios de aspecto similar al de las hepáticas, si bien el pedúnculo es corto y la pared anteridial está menos diferenciada. Las cámaras arquegoniales, más pequeñas, incluyen un único arquegonio (órgano femenino) que está estrechamente embutido en la cámara, por lo que resulta poco diferenciado del resto de las células del talo, y que presenta seis células del canal del cuello. La ovocélula se encuentra así en el fondo de una estructura en forma de botella abierta por su extremo dorsal a la superficie.
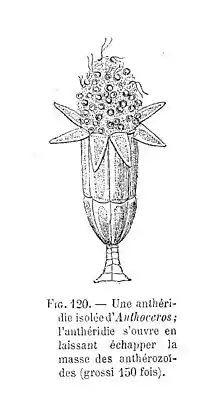
Los anterozoides presentan dos flagelos apicales y un cuerpo celular de simetría bilateral arrollado en espiral a derechas, como en las hepáticas. La estructura del cinetosoma del undulipodio es peculiar del grupo, al faltar la disposición estrellada de los microtúbulos. Los anterozoides se desplazan hasta los arquegonios utilizando la película de humedad que recubre los talos debido a la atracción que ejercen sobre ellos las sustancias producidas por la desintegración de las células del canal del cuello. Penetran hasta la ovocélula a lo largo del cuello del arquegonio y uno de los anterozoides la fecunda.
Existen especies unisexuales (dioicas, con anteridios y oogonios en talos separados) y bisexuales (monoicas, con anteridios y oogonios en el mismo talo).
Esporófito
La ovocélula fecundada (zigoto) se divide en dos de manera longitudinal la primera vez, siendo la segunda división transversal, dando así origen al desarrollo del embrión diploide. Tras sucesivas divisiones, las células superiores darán lugar al esporangio y las inferiores al pie del mismo, hinchado y fijo a las células del talo mediante estructuras similares a rizoides, que forman un haustorio, cuya misión es extraer nutrientes del talo para alimentar al esporangio. Este carece de seta (sésil) y presenta un meristemo intercalar basal, que lo hace crecer ininterrumpidamente mientras se den las condiciones ambientales adecuadas. El esporófito es clorofílico durante toda su vida. Al crecer, el esporófito empuja, haciendo que se alargue la cubierta gametofítica que le envuelve, perforándola en su extremidad. Esta porción del gametófito que rodea la base del esporangio adopta una forma de vaina cilíndrica o troncocónica y recibe el nombre de involucro.
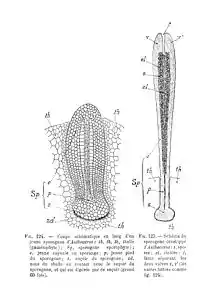
El esporangio presenta una epidermis con cutícula que, en la mayoría de las especies, tiene estomas con las células oclusivas características y se abren al interior a una red de espacios intercelulares. Sus células presentan cloroplastos. Durante el crecimiento, el esporangio adopta forma de cápsula alargada, con una columela central cilíndrica de tejido estéril (raramente poco desarrollada o ausente) rodeada de un tejido esporógeno (arquesporio). La columela central tiene un número reducido de filas verticales de células. Las células del arquesporio producen al dividirse dos células madres: una da lugar a una tétrada de esporas y la otra a un pseudoeláter (célula estéril alargada), que puede subdividirse para alcanzar una estructura pluricelular y ligeramente ramificada, de paredes delgadas, aunque en algunos casos éstas están engrosadas irregularmente o pueden presentar engrosamientos helicoidales. La misión de estos pseudoeláteres es contribuir en sus primeros momentos a la nutrición de las esporas y en la madurez a la dispersión de las mismas, al secarse cuando se exponen al aire y distenderse como resortes. Las esporas pueden dispersarse en forma de mónadas o tétradas; la ornamentación de su pared difiere entre las especies, así como su coloración, que varía del amarillo al negro. En la madurez, la cápsula se abre longitudinalmente en dos valvas desde la parte apical por dos líneas opuestas de menor resistencia, liberando las esporas. La maduración ocurre de manera gradual desde el ápice hacia la base (excepto en Notothylas, en que es sincrónica), y las valvas pueden ayudar a la dispersión arrollándose y desenrollándose helicoidalmente en función de la humedad ambiental. Mientras el esporófito está vivo, se sigue formando arquesporio de manera gradual en la base de la cápsula y ésta sigue liberando esporas.
Citogenética
.jpg.webp)
El número cromosómico de las especies conocidas es muy bajo y bastante constante, en las especies dioicas es n = 4 + 1 cromosoma sexual y en las monoicas es n = 5 o 6. Se conocen escasos registros de poliploidía e hibridación.
Fitoquímica
Los antocerotófitos son incapaces de sintetizar flavonoides del tipo de los antocianos, pero sintetizan lignanos (ácidos antocerotónico y megacerotónico). El pirenoide del cloroplasto contiene la enzima RuBisCO, por lo que presenta un mecanismo de concentración de carbono ausente del resto de las plantas terrestres. Las paredes celulares presentan xilanos y xiloglucanos, como en los traqueófitos. Las paredes celulares contienen azúcares peculiares, como el disacárido α-D-glucuronosil-(1→3)-L-galactosa o el monosacárido acofriosa (3-O-metilramnosa), este último presente también en algunos carófitos y otros grupos de algas relacionados con ellos, en hepáticas, musgos y licopodiófitos homospóreos. Las células acumulan como productos de su metabolismo diferentes compuestos, algunos poco comunes, entre los que se han identificado ésteres del ácido cinámico (como el metilcafeato y el metil-4-cumarato) necesarios como intermediarios en la producción de la lignina, el alcaloide antocerodiazonina, el ácido isorínico (ácido cafeoil-4'-hidroxifenilláctico) y el ácido rosmarínico (en forma de un 3'-O-β-D-glucósido).
Ecología
.jpg.webp)
Debido a su asociación simbiótica con la cianobacteria Nostoc, los antocerotófitos incorporan el N2 atmosférico, absorbiendo por el talo agua y materia inorgánica. Esto los hace relativamente independientes del medio y les permite soportar condiciones extremas, por lo que se encuentran entre las plantas pioneras sobre la roca desnuda. Propios de zonas tropicales y templadas, recubren rocas y troncos húmedos.
Simbiosis
Los antocerotófitos son las únicas plantas en que los gametófitos mantienen una asociación con cianobacterias del género Nostoc en el interior del talo. La asociación presente en las hepáticas del orden Blasiales es externa. Las especies de los géneros Phaeoceros y Anthoceros presentan una asociación endofítica con hongos glomeromicétidos, con hifas presentes en los espacios intercelulares y en las cavidades mucilaginosas. Al menos una especie de Endogone (Mucoromycotina) está asociada a Paraphymatoceros.[3] Tanto unos hongos como otros forman micorrizas con su hospedante. Solo se encuentran hongos exclusivamente intracelulares en Phaeomegaceros. El conocimiento de estas asociaciones es aún muy limitado.
Filogenia
Los antocerotófitos son una de las ramas basales del clado de los estomatófitos, así llamados por presentar estomas, según recientes estudios de análisis filogenéticos basados tanto en morfología como en genética. Los principales caracteres derivados que definen a los antocerotófitos son: la incapacidad de sintetizar flavonoides, la simbiosis con cianobacterias del género Nostoc, la formación de los anteridios a partir de una célula subepidérmica, los arquegonios inmersos en el talo, la primera división longitudinal del zigoto, la cutícula del esporófito, la ausencia de seta del mismo, los pseudoeláteres del esporogonio y la pérdida del intrón nad2i1 en el genoma mitocondrial. La hipótesis que suponía la existencia de dos grandes grupos de plantas, denominados Bryata (plantas no vasculares) y Tracheata (plantas vasculares)[4] no ha encontrado confirmación. La relación filogenética de los antoceros con las otras embriofitas son inciertas siendo propuestas como hermanas de las briofitas tradicionales (musgos y hepáticas), lo cual ha tenido más respaldo por los análisis moleculares,[5][6] o de las plantas vasculares (Tracheophyta).[7]
La divergencia de los antepasados comunes de los antocerotófitos y el resto de las plantas se data del Ordovícico Superior al Silúrico Inferior, hace unos 430-450 millones de años, aunque algunos estudios apoyan la existencia de un antepasado común con las hepáticas hace unos 450-475 millones de años. La dificultad en presentar una estimación plausible de la fecha de aparición se debe, por una parte, a una rápida cladogénesis, que en unos 70 millones de años dio lugar a los tres grupos de plantas terrestres monosporangiadas (los briófitos en sentido amplio) y al antepasado de las plantas polisporangiadas (las traqueofitas) y por otra parte a la escasez del registro fósil de plantas tan delicadas. El antepasado común más reciente de los antocerotófitos actuales se data del Devónico Inferior, hace unos 365 millones de años y se estima que la familia Dendrocerotaceae apareció hace unos 130 millones de años, lo que, en rasgos generales, coincide con las edades de divergencia estimadas para las restantes plantas.
Taxonomía
A pesar del escaso número de representantes, la sistemática del filo no está asentada. En 2005, se presentaron de manera independiente dos propuestas diferentes de clasificación para los antocerotófitos: la de Stotler y Crandall-Stotler[8] y la de Frey y Stech.[9] Ambas coinciden en proponer dos clases, una primitiva (Clase Leiosporocerotopsida) con un solo Orden (Leiosporocerotales) y una sola familia (Leiosporocerotaceae (que incluye tan solo el género Leiosporoceros) y otra más derivada, la Clase Anthocerotopsida. La división de esta última, propuesta por Stotler y Crandall-Stotler es:
Orden Anthocerotales
- Familia Anthocerotaceae
Orden Notothyladales
- Familia Notothyladaceae
- Familia Dendrocerotaceae
mientras que Frey y Stech proponen la siguiente división:
Orden Anthocerotales
- Familia Anthocerotaceae
- Familia Foliocerotaceae
Orden Phaeocerotales
- Familia Phaeocerotaceae
- Familia Notothyladaceae
Orden Dendrocerotales
- Familia Dendrocerotaceae
siendo ambas prácticamente congruentes, salvo por las categorías otorgadas a los diferentes táxones y la disposición de los géneros dentro de las familias.
Esta clasificación ha sido posteriormente objeto de una revisión[10] basada en el análisis filogenético de las secuencias del gen cloroplastidial rbcL, el gen mitocondrial nad5 y el gen nuclear 18S. Se añadió un nuevo orden Phymatocerotales y una nueva familia Phymatocerotaceae para contener el género Phymatoceros. La relación filogenética existente queda reflejada en el siguiente cladograma de los órdenes:[11]
| Anthocerotophyta |
| ||||||||||||||||||||||||
Ésta es la clasificación que se sigue aquí y se expone a continuación.
El filo Anthocerotophyta se divide en dos clases:
- Clase Leiosporocerotopsida Stotl. & Crand.-Stotl., 2005
- Talo sólido, cavidades esquizógenas presentes solo en algunos talos viejos. Poros de mucílago ausentes en los tejidos ya colonizados por Nostoc, presentes solo en plantas jóvenes sin colonizar. Colonias de Nostoc en cordones longitudinales ramificados, dispuestos en la región media del talo. Cloroplasto solitario, sin pirenoide, compartimentado, con grana y plastoglóbulos centrales y almidón acumulado en la periferia. Anteridios numerosos (hasta 70 por cámara anteridial), con las células de la pared dispuestas en pisos. Esporangio con suturas profundas, epidermis con estomas, una cubierta de 9-10 capas de células asimiladoras, un arquesporio abundante con varias capas de tétradas muy pequeñas entremezcladas con abundantes pseudoeláteres alargados y con engrosamiento en espiral y una columela indistinta. Esporas minúsculas, ovoides, de pared fina, monoletas, lisas, en tétradas isobilaterales. Pseudoeláteres largos, normalmente unicelulares, de pared gruesa.
- Esta clase consta de un solo orden (Leiosporocerotales Hässel, 1988) y una sola familia: Leiosporocerotaceae Hässel, 1986.
- Poros de mucílago de la cara ventral del talo presentes (excepto en Notothylas) en plantas jóvenes y adultas. Colonias de Nostoc globosas, en la región ventral del talo. Cloroplasto con pirenoide central desarrollado (que puede faltar en algunas especies), con o sin compartimentación, o con varios cloroplastos por célula. Esporangio con suturas obsoletas, una cubierta de pocas capas de células asimiladoras, un arquesporio con una capa de tétradas grandes entremezcladas con escasos pseudoeláteres pequeños y ramificados o largos y espiralados y una columela desarrollada (excepto en Notothylas). Esporas normalmente en tétradas tetraédricas, ornamentadas.
- Esta clase se divide en tres subclases:
- Subclase Anthocerotidae Rosenv., 1958, con un solo orden:
- Orden Anthocerotales Limpricht in Cohn, 1877
- Talos con laminillas dorsales. Cavidades mucilaginosas esquizógenas. Pirenoide presente o ausente. Cámaras anteridiales con numerosos anteridios (hasta 60), con las células de la pared dispuestas en 4 pisos. Esporangio con estomas (excepto en Foliocerus incurvus (Steph.) D.C.Bharadwaj, 1972). Esporas más o menos netamente triletas, oscuras (gris ahumado, pardooscuras o negruzcas); ornamentación espinosa, punteada, baculada, dentada, tuberculada o laminada. Pseudoeláteres normalmente con engrosamiento helicoidal.
- Este orden incluye una sola familia: Anthocerotaceae (Gray) Dumort., 1829.
- Subclase Notothyladidae R.J.Duff, J.C.Villarreal, Cargill & Renzaglia, 2007, con un solo orden:
- Orden Notothyladales Hyvönen & Piippo, 1993
- Talos sólidos, sin laminillas. Pirenoide presente o ausente. Cámaras anteridiales con 2-4(-8) anteridios. Esporangios con suturas elaboradas, rudimentarias o ausentes, con o sin estomas. Esporas amarillas a negruzcas, frecuentemente con faja ecuatorial. Pseudoeláteres a veces ausentes.
- Este orden incluye una única familia: Notothyladaceae (Milde) Müll.Frib. ex Prosk., 1960
- Subclase Dendrocerotidae R.J.Duff, J.C.Villarreal, Cargill & Renzaglia, 2007
- Talos sólidos (excepto en Dendroceros subgénero Apoceros). Cámaras anteridiales normalmente con un anteridio. Cloroplasto normalmente sin pirenoide o con este modificado.
- Está dividida en dos órdenes:
- Orden Phymatocerotales R.J.Duff, J.C.Villarreal, Cargill & Renzaglia, 2007
- Talos sólidos, lineares a lingulados, dioicos, con un nervio medio fino, los bordes adelgazados. Tubérculos ventrales largamente pedunculados. Pirenoide ausente. Cámaras anteridiales con 1-3 anteridios. Esporangio robusto, con columela y estomas. Esporas unicelulares, doradas, con ornamentación vermiculada y abombamiento distal. Pseudoeláteres sin engrosamiento helicoidal.
- Este orden incluye una sola familia: Phymatocerotaceae R.J.Duff, J.C.Villarreal, Cargill & Renzaglia, 2007
- Orden Dendrocerotales Hässel, 1988
- Cámaras anteridiales con un anteridio. Esporangio sin estomas. Esporas amarillas a transparentes, con exosporio delgado, hialino. Pseudoeláteres con engrosamiento helicoidal.
- Este orden incluye una única familia: Dendrocerotaceae (Milde) Hässel, 1988
Distribución

Aunque se conoce la distribución de las especies de zonas templadas, sobre todo Europa y Norteamérica, con relativo detalle, el desconocimiento de la biodiversidad y la distribución en las áreas tropicales es elevado. Se han podido desvelar algunas pautas distributivas y centros de diversidad. Las floras de antocerotófitos más ricas se centran en India, África occidental y Japón, aunque ciertos géneros muestran una concentración elevada de especies en ciertas zonas: Notothylas en India y Nepal, Dendroceros en Asia y algunas secciones de Phaeoceros y Anthoceros en las Américas y en Australia. Este conocimiento sufre además de diferencias en los tratamientos taxonómicos de las especies y de la ausencia de revisiones modernas para muchas regiones terrestres.
Fósiles
No existen muchos fósiles incontestables de antocerotófitos. El fósil más antiguo asignado a los antocerotófitos es la espora fósil Stoverisporites lunaris del Cretáceo Inferior de Argentina, que se asemeja a las esporas de los Phaeomegaceros actuales, un género evolutivamente avanzado. Otros fósiles existentes pueden ser atribuidos razonablemente a Notothylas y a algunas especies de Phaeoceros.
Notas
- La homología de los pseudoeláteres con los eláteres de las hepáticas está aún en discusión.
- La pérdida del pirenoide y la adquisición de múltiples cloroplastos en las células de los antocerotófitos es una transformación evolutiva que parece responder a un cambio climático mediado por un aumento progresivo de la concentración de CO2.
- La homología de los poros de mucílago del gametófito con estomas no ha sido demostrada de manera indiscutible. Estomas con las típicas células de guarda arriñonadas solo se encuentran en los esporófitos. A pesar de eso, algunos autores los siguen mencionando como estomas.
Referencias
- Purves, David Sadava 2008. Vida. La Ciencia de la Biología. Cap. 28 Plantas sin semilla. Ed. Médica Panamericana
- Britannica Enciclopedia Moderna 2011.
- Bidartondo, Martin I.; Read, David J.; Trappe, James M.; Merckx, Vincent; Ligrone, Roberto; Duckett, Jeffrey G. (2011). «The dawn of symbiosis between plants and fungi». Biology Letters 7: 574-577.
- Margulis, Lynn; Chapman, Michael J. (2009). Kingdoms & Domains. An Illustrated Guide to the Phyla of Life on Earth (en inglés). Amsterdam: Elsevier. pp. lxxii + 659 págs. ISBN 0-7167-3026-X.
- Harris, Brogan J. (2020). «Phylogenomic Evidence for the Monophyly of Bryophytes and the Reductive Evolution of Stomata». Current Biology 30 (11): P2201-2012.E2. PMID 32302587. S2CID 215798377. doi:10.1016/j.cub.2020.03.048. hdl:1983/fbf3f371-8085-4e76-9342-e3b326e69edd.
- Cox, Cymon J. (2014). «Conflicting Phylogenies for Early Land Plants are Caused by Composition Biases among Synonymous Substitutions». Systematic Biology 63 (2): 272-279. PMC 3926305. PMID 24399481. doi:10.1093/sysbio/syt109.
- Cox, Cymon J. (2018). «Land Plant Molecular Phylogenetics: A Review with Comments on Evaluating Incongruence Among Phylogenies». Critical Reviews in Plant Sciences 37 (2–3): 113-127. S2CID 92198979. doi:10.1080/07352689.2018.1482443. hdl:10400.1/14557.
- Stotler, Raymond E.; Crandall-Stotler, Barbara (2005). «A revised classification of the Anthocerotophyta and a checklist of the hornworts of North America, north of Mexico». The Bryologist 108 (1): 16-26.
- Frey, Wolfgang; Stech, Michael (2005). «A morpho-molecular classification of the Anthocerotophyta (hornworts)». Nova Hedwigia 80 (3-4): 541-545.
- Duff, R. Joel; Villarreal, Juan Carlos; Cargill, D. Christine; Renzaglia, Karen S. (2007). «Progress and challenges toward developing a phylogeny and classification of the hornworts». The Bryologist 110 (2): 214-243.
- Duff, R. Joel; Juan Carlos Villarreal; D. Christine Cargill; Karen S. Renzaglia (2007). «Progress and challenges toward a phylogeny and classification of the hornworts». The Bryologist 110 (2): 214-243. doi:10.1639/0007-2745(2007)110[214:PACTDA]2.0.CO;2.
Bibliografía
- Shaw, Jonathan; Renzaglia, Karen (2004). «Phylogeny and diversification of Bryophytes». American Journal of Botany 91 (10): 1557-1581.
- Villarreal, Juan Carlos; Cargill, D. Christine; Hagborg, Anders; Söderström, Lars; Renzaglia, Karen Sue (2010). «A synthesis of hornwort diversity: Patterns, causes and future work». Phytotaxa 9: 150-166.
- Von Konrat, Matt; Shaw, A. Jonathan; Renzaglia, Karen Sue, eds. (2010). Bryophytes: the closest living relatives of early land plants. Phytotaxa 9. Auckland (Nueva Zelanda): Magnolia Press. p. 278. ISBN 9781869775971. OCLC 676694037.
Enlaces externos
 Wikimedia Commons alberga una categoría multimedia sobre Anthocerotophyta.
Wikimedia Commons alberga una categoría multimedia sobre Anthocerotophyta.
- Portal de Anthocerotophyta
- Historia de fósiles y descripciones de Anthocerotophyta
- Anthocerotophyta de Nva. Zelanda
| Control de autoridades |
|
|---|
 Datos: Q191156
Datos: Q191156- Multimedia: Anthocerotophyta / Q191156
 Especies: Anthocerotophyta
Especies: Anthocerotophyta