Beta oxidación
La beta oxidación (β-oxidación) es un proceso catabólico de los ácidos grasos en el cual sufren la eliminación de un par de átomos de carbono sucesivamente en cada ciclo del proceso mediante oxidación, hasta que el ácido graso se descompone por completo en forma de moléculas Acetil-Coenzima A (Acetil-CoA), que serán posteriormente oxidados en la mitocondria para generar energía química en forma de Adenosina Trifosfato (ATP). La β-oxidación de ácidos grasos consta de cuatro reacciones recurrentes.

El resultado de dichas reacciones son unidades de dos carbonos en forma de acetil-CoA, molécula que pueden ingresar en el ciclo de Krebs, y coenzimas redox reducidas (Nicotinamida-Adenina-H -NADH- y Flavina-Adenina-H2 -FADH2- ) que pueden ingresar en la cadena respiratoria.
No obstante, antes de que produzca la oxidación, los ácidos grasos deben activarse con coenzima A y atravesar la membrana mitocondrial interna, que es impermeable a ellos.
Pasos previos
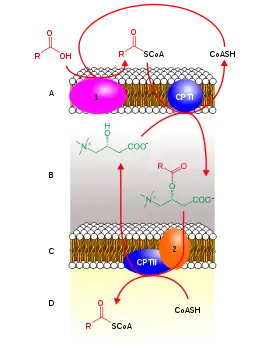
Rojo: acil-CoA, verde: carnitina, Rojo+verde: acilcarnitina, CoASH: coenzima A, CPTI: carnitina palmitoiltransferasa I, CPTII: carnitina palmitoiltransferasa II, 1: acil-CoA sintetasa, 2: translocasa, A: membrana mitocondrial externa, B: espacio intermembrana, C: membrana mitocondrial interna, D: matriz mitocondrial
Activación de los ácidos grasos
El paso previo a esas cuatro reacciones es la activación de los ácidos grasos con CoA formando Acil coenzima A (acil CoA, R–CO–SCoA), la cual tiene lugar en el retículo endoplasmático (RE) o en la membrana mitocondrial externa, donde se halla la acil-CoA sintetasa (o ácido graso tioquinasa), la enzima que cataliza esta reacción:[1]
El ácido graso se une al coenzima A (CoA-SH), reacción que consume dos enlaces de alta energía del ATP, generando Adenosina Monofosfato, Difosfato inorgánico y agua.
Entrada a la matriz mitocondrial
Posteriormente debe usarse un transportador, la carnitina, para translocar las moléculas de acil-CoA al interior de la matriz mitocondrial, ya que la membrana mitocondrial interna es impermeable a los acil-CoA.
La carnitina se encarga de llevar los grupos acilo al interior de la matriz mitoncondrial por medio del siguiente mecanismo.
- La enzima carnitina palmitoiltransferasa I (CPTI) de la membrana mitocondrial externa elimina el coenzima A de la molécula de acil-CoA y, a la vez, transfiere y une el ácido graso a la carnitina situada en el espacio intermembrana (EIM), originado acilcarnitina; el CoA queda libre en el citosol para poder activar otro ácido graso. Acil-CoA (Citosol) + Carnitina → Acil-Carnitina (EIM) + SH-CoA
- A continuación, una proteína transportadora, llamada translocasa, situada en la membrana mitocondrial interna, transfiere la acilcarnitina a la matriz mitoncondrial y, paralelamente, la carnitina palmitoiltransferasa II (CPTII) une una molécula de CoA de la matriz al ácido graso, regenerando así el acil-CoA Acil-Carnitina (EIM) + SH-COA → Acil-CoA (MM) + Carnitina
- La carnitina se devuelve al espacio intermembrana por la proteína transportadora y reacciona con otro acil-CoA, repitiéndose el ciclo.
La carnitina es un derivado aminoacídico que participa en el circuito vascular reduciendo niveles de triglicéridos y colesterol en sangre. Se produce naturalmente en el hígado a partir de los aminoácidos L-metionina y L-lisina. La carnitina es fuertemente inhibida por el malonil-CoA, uno de los pasos reguladores en el proceso de lipogénesis.
β-oxidación
En la siguiente tabla se resumen las cuatro reacciones que conducen a la liberación de una molécula de [acetil CoA] y al acortamiento en dos átomos de carbono del ácido graso:
| Descripción | Reacción | Enzima | Producto final |
| Oxidación por FAD El primer paso es la oxidación del ácido graso por la acil-CoA deshidrogenasa. La enzima cataliza la formación de un doble enlace entre C-2 (carbono α) y C-3 (carbono β). |  | acil-CoA deshidrogenasa | trans-Δ2-enoil-CoA |
| Hidratación El siguiente paso es la hidratación del enlace entre C-2 y C-3. Esta reacción es estereospecífica, formando solo el isómero L. | 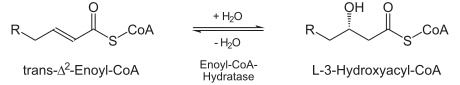 | enoil CoA hidratasa | L-3-hidroxiacil CoA |
| Oxidación por NAD+ El tercer paso es la oxidación del L-3-hidroxiacil CoA por el NAD+, lo que convierte el grupo hidroxilo (–OH) en un grupo cetona (=O). |  | L-3-hidroxiacil CoA deshidrogenasa | 3-cetoacil CoA |
| Tiólisis El paso final es la separación del 3-cetoacil CoA por el grupo tiol de otra molécula de CoA. El tiol es insertado entre C-2 y C-3. | 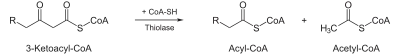 | β-cetotiolasa | Una molécula de acetil CoA y una de acil CoA con dos carbonos menos |
Oxidación por FAD
El primer paso es la oxidación por deshidrogenación del ácido graso activado (acil-CoA graso) por FAD. La enzima acil-CoA-deshidrogenasa, una flavoproteína que tiene a la coenzima Flavina-Adenosina(FAD) unida covalentemente, cataliza la formación de un doble enlace entre C-2 y C-3. Los productos finales son FADH2 y un acil-CoA-betainsaturado (trans-Δ2-enoil-CoA) ya que el carbono beta del ácido graso se une con un doble enlace al perder dos hidrógenos (que son ganados por el FAD).
Acil-CoA + FAD → Trans-Δ2-enoil-CoA + FADH2
Hidratación
El siguiente paso es la hidratación (adición de una molécula de agua) del doble enlace trans entre C-2 y C-3, generando un Grupo hidroxilo en el tercer carbono (beta β). Esta reacción es catalizada por enoil-CoA hidratasa y se obtiene un L-betahidroxiacil-CoA (L-3-hidroxiacil CoA); es una reacción estereospecífica, formándose exclusivamente el isómero L.
Trans-Δ2-enoil-CoA + H2O → L-β-hidroxiacil-CoA
Oxidación por NAD+
El tercer paso es la oxidación por deshidrogenación de L-3-hidroxiacil CoA por Nicotinamida-Adenina (NAD), catalizada por la L-3-hidroxiacil CoA deshidrogenasa. Esto convierte el grupo hidroxilo del carbono β en un grupo cetónico(lo satura), perdiendo el hidrógeno del hidroxilo que es ganado por el NAD. El producto final es 3-cetoacil-CoA con lo que el carbono beta ya ha sido oxidado y está preparado para la escisión.
L-β-hidroxiacil-CoA + NAD → β-cetoacil-CoA + NADH
Tiólisis
El paso final para la ruptura del cetoacil-CoA entre C-2 y C-3 por el grupo tiol de otra molécula de CoA. Esta reacción es catalizada por β-cetotiolasa y da lugar a una molécula de acetil CoA y un acil CoA con dos carbonos menos.
β-cetoacil-CoA + SH-CoA → Acil-CoA(-2C) + Acetil-CoA
Conclusión
Estas cuatro reacciones continúan hasta que la escisión completa de la molécula en unidades de acetil CoA, siendo el último ciclo la metabolizacion del butanoil-CoA a dos complejos acetil-CoA.
Por cada ciclo, se forma una molécula de FADH 2, una de NADH y una de acetil CoA.
Esto supone una visión de un ciclo en espiral ya que repite los mismos pasos pero con diferentes sustancias procedentes del ciclo anterior. Por ello se le llama hélice de Lynen.
Los ácidos grasos de un número impar de carbonos siguen las mismas vías, esto es, ciclos de deshidrogenación, hidratación, deshidrogenación y tiólisis. Sin embargo, en el último paso del ciclo, se forma una molécula de propionil-CoA (3 carbonos), potencialmente gluconeogénico, a diferencia de los acetil-CoA de dos carbonos (el Acetil-CoA que ingrese en el ciclo de Krebs es completamente oxidado a 2 moléculas de CO2), sin embargo, una ruta alternativa puede transformar al propionil-CoA en sustrato para el Ciclo de Krebs por medio de su carboxilación en Metilmalonil-CoA y la posterior isomerizacion de éste en Succinil-CoA, intermediario en el mencionado ciclo.
Propionil-CoA (+ATP + HCO3-)→Propionil-CoA Carboxilasa → Metilmalonil-CoA→ Metilmalonil-CoA Mutasa → Succinil-CoA
Rendimiento energético
Dado que durante la β-oxidación la cadena de carbonos de los ácidos grasos se rompe en unidades de dos carbonos (unidas al coenzima A) y que cada rotura produce una molécula de FADH2 y una molécula de NADH + H+, es fácil calcular las moléculas de ATP generadas en la oxidación completa de un ácido graso. FADH2 y NADH van a la cadena respiratoria y los acetil-CoA ingresan en el Ciclo de Krebs donde generan GTP y más moléculas de FADH2 (una) y NADH (tres). Es decir, cada ciclo generara tres moleculas de ATP por el NADH, dos por el FADH, y 11 ATP y un GTP por el acetil-CoA que se metabolice en el Ciclo de Krebs, dando como balance 17 nucleótidos energéticos
Si tomamos como ejemplo el ácido palmítico, ácido graso saturado de 16 carbonos, el rendimiento energético es el siguiente:
| Rendimiento de la betaoxidación del ácido graso (Palmitato, 16 C) | ||||
|---|---|---|---|---|
| Molécula | Número | Equivalencia de moléculas de ATP [cita requerida] |
Ciclo metabólico | Total ATP generados |
| NADH | cadena respiratoria | |||
| FADH2 | cadena respiratoria | |||
| acetil-CoA | ciclo de Krebs | |||
| Total | 129 | |||
Teniendo en cuenta los dos enlaces de alta energía que se utilizan en la activación del ácido graso a acil-CoA, se obtiene un rendimiento neto de 129 moléculas de nucleótidos energéticos por cada molécula de palmitato metabolizada. Obviamente, cuanto más larga es la molécula de ácido graso, más moléculas de ATP se generan.
Referencias
- Devlin, T. M. 2004. Bioquímica, 4ª edición. Reverté, Barcelona. ISBN 84-291-7208-4
Véase también
- Oxidación de ácidos grasos
- Alfa oxidación
- Omega oxidación
- Oxidación peroxisómica de ácidos grasos
| Control de autoridades |
|
|---|
 Datos: Q306213
Datos: Q306213 Multimedia: Beta-oxidation / Q306213
Multimedia: Beta-oxidation / Q306213