Bomba sodio-potasio
La bomba sodio-potasio es una enzima (una ATPasa) que realiza un transporte bombeando iones de potasio hacia el interior de la célula y al mismo tiempo bombea iones de sodio desde el interior hacia el exteror celular. Esta bomba es responsable de mantener las diferencias de concentración de sodio y de potasio a través de la membrana celular, así como de establecer un voltaje eléctrico negativo en el interior de las células. Se encuentra en la membrana plasmática de todas las células animales.[1]
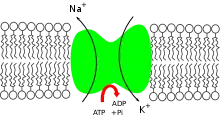
La bomba expulsa tres iones sodio (Na+) hacia la matriz extracelular a la vez que ingresa dos iones potasio (K+) hacia el citoplasma mediante transporte activo que ocupa como fuente de energía el ATP. Este bombeo permanente permite mantener el gradiente electroquímico de solutos con una concentración elevada de potasio dentro de la célula y bajo fuera, mientras que la concentración de sodio es baja dentro de la célula y elevada fuera.[2]
Historia
Esta proteína fue descubierta por el danés Jens Skou de forma casual en 1957, al estudiar los nervios del cangrejo Carcinus maenas y por ello recibió el premio Nobel en 1997.[2][3]
Desde entonces la investigación ha determinado muchos de los aspectos tanto de la estructura y funcionamiento de la proteína, como de su función en la fisiología, de tremenda importancia en la medicina.
Estructura

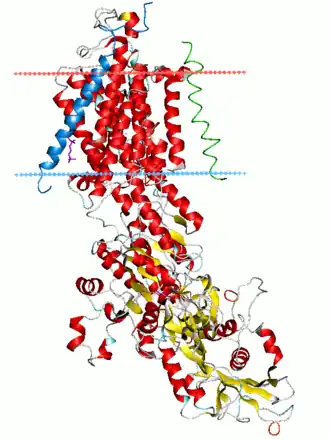
La bomba sodio-potasio está compuesta de 3 subunidades: α, β y γ. La subunidad α es una proteína de 110 kDa encargada de la función de transporte, tiene un sitio para la unión del ATP y un sitio de fosforilación. La subunidad β es una proteína transmembrana de 55 kDa altamente glicosilada que aumenta la eficiencia de traslación y la estabilidad de la subunidad α. En los mamíferos existen tres subunidades α y dos subunidades β con α3 y β2 expresadas principalmente en las neuronas y α2 y β1 en las glías. α1 se expresa ubicuamente. La subunidad γ, una proteína FXYD, es específica de ciertos tejidos y modifica la afinidad por Na+ y K+, la cinética de la bomba y estabiliza la conformación de la bomba.[2][4]
Es una ATPasa de transporte tipo P, es decir, sufre fosforilaciones reversibles durante el proceso de transporte. Durante la fosforilación la bomba sufre un cambio conformaciones desde hélice alfa a lámina beta.[4]
Está formada por dos subunidades, alfa y beta, que forman un tetrámero integrado en la membrana. La subunidad alfa está compuesta por diez segmentos transmembrana y en ella se encuentra el centro de unión del ATP que se localiza en el lado citosólico de la membrana (tiene un peso molecular de aproximadamente 100 000 daltons). También posee dos centros de unión al potasio extracelulares y tres centros de unión al sodio intracelulares que se encuentran accesibles para los iones según si la proteína está fosforilada. La subunidad beta contiene una sola región helicoidal transmembrana y no parece ser esencial para el transporte ni para la actividad, aunque podría realizar la función de anclar el complejo proteico a la membrana lipídica.tacar sobre el funcionamiento de la bomba sodio-potasio es que esta a pesar de que de forma general significa un gasto energético para la célula, si se encuentra en dirección a favor del gradiente de concentración puede llegar a unir un grupo fosfato a un adenosin difosfato, generando una cantidad de energía en forma de ATP considerable para la célula.
Los procesos que tienen lugar en el transporte son:
Unión de tres Na+ a sus sitios activos. Fosforilación de la cara citoplasmática de la bomba que induce a un cambio de conformación en la proteína. Esta fosforilación se produce por la transferencia del grupo terminal del ATP a un residuo de ácido aspártico de la proteína. El cambio de conformación hace que el Na+ sea liberado al exterior. Una vez liberado el Na+, se unen dos iones de K+ a sus respectivos sitios de unión de la cara extracelular de las proteínas. La proteína se desfosforila produciéndose un cambio conformacional de ésta, lo que produce una transferencia de los iones de K+ al citosol.
Función
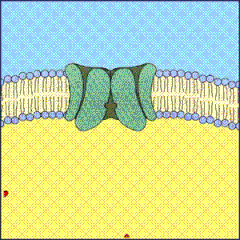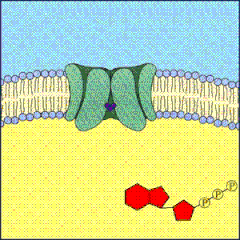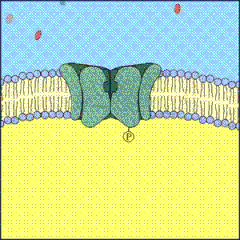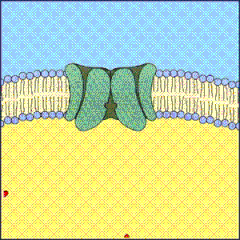
La bomba de sodio-potasio es crucial e imprescindible para que exista la vida animal. Por ello se encuentra en todas las membranas celulares de los animales, en mayor medida en células excitables como las células nerviosas y células musculares donde la bomba puede llegar a acaparar los dos tercios del total de la energía en forma de ATP de la célula.
El funcionamiento de la bomba electrogénica de Na+/ K+(sodio-potasio), se debe a un cambio de conformación en la proteína que se produce cuando es fosforilada por el ATP. Como el resultado de la catálisis es el movimiento transmembrana de cationes, y se consume energía en forma de ATP, su función se denomina transporte activo. La demanda energética es cubierta por la molécula de ATP, que al ser hidrolizada, separa un grupo fosfato, generando ADP y liberando la energía necesaria para la actividad enzimática. En las mitocondrias, el ADP es fosforilado durante el proceso de respiración generándose un reservorio continuo de ATP para los procesos celulares que requieren energía. En este caso, la energía liberada induce un cambio en la conformación de la proteína una vez unidos los tres cationes de sodio a sus lugares de unión intracelular, lo que conlleva su expulsión al exterior de la célula. Esto hace posible la unión de dos iones de potasio en la cara extracelular que provoca la desfosforilación de la ATP, y la posterior traslocación para recuperar su estado inicial liberando los dos iones de potasio en el medio intracelular.[cita requerida]
Algo que cabe destacar sobre el funcionamiento de la bomba sodio-potasio es que esta a pesar de que de forma general significa un gasto energético para la célula, si se encuentra en dirección a favor del gradiente de concentración puede llegar a unir un grupo fosfato a un adenosin difosfato, generando una cantidad de energía en forma de ATP considerable para la célula.
Los procesos que tienen lugar en el transporte son:
- Unión de tres Na+ a sus sitios activos.
- Fosforilación de la cara citoplasmática de la bomba que induce a un cambio de conformación en la proteína. Esta fosforilación se produce por la transferencia del grupo terminal del ATP a un residuo de ácido aspártico de la proteína.
- El cambio de conformación hace que el Na+ sea liberado al exterior.
- Una vez liberado el Na+, se unen dos iones de K+ a sus respectivos sitios de unión de la cara extracelular de las proteínas.
- La proteína se desfosforila produciéndose un cambio conformacional de ésta, lo que produce una transferencia de los iones de K+ al citosol.
Mantenimiento de la osmolaridad y del volumen celular
La bomba de Na+/K+ desempeña un papel muy importante en el mantenimiento del volumen celular. Entre el interior y el exterior de la célula existen diferentes niveles de concentración de solutos. Como quiera que la bomba extrae de la célula más moléculas de las que introduce tiende a igualar las concentraciones y, consecuentemente, la presión osmótica. Sin la existencia de la bomba, dado que los solutos orgánicos intracelulares, a pesar de contribuir en sí mismos poco a la presión osmótica total, tienen una gran cantidad de solutos inorgánicos asociados, la concentración intracelular de estos (que generalmente son iones) es mayor que la extracelular. Por ello, se produciría un proceso osmótico, consistente en el paso de agua a través de la membrana plasmática hacia el interior de la célula, que aumentaría de volumen y diluiría sus componentes. Las consecuencias serían catastróficas ya que la célula podría llegar a reventar (proceso conocido como lisis).
Absorción y reabsorción de moléculas
El gradiente producido por el Na+ impulsa el transporte acoplado (activo secundario) de diferentes moléculas al interior de la célula. Lo que quiere decir que el fuerte gradiente que impulsa al sodio a salir de la célula (véase más adelante) es aprovechado por proteínas especiales de membrana para "arrastrar" otros solutos de interés utilizando la energía que se libera cuando el sodio se introduce en la célula. Ejemplos de este proceso son la absorción de nutrientes en las células de la mucosa intestinal y la reabsorción de solutos en el túbulo renal.
Potencial eléctrico de membrana
Esta bomba es una proteína electrogénica ya que bombea tres iones cargados positivamente hacia el exterior de la célula e introduce dos iones positivos en el interior celular. Esto supone el establecimiento de una corriente eléctrica neta a través de la membrana celular, lo que contribuye a generar un potencial eléctrico entre el interior y el exterior de la célula ya que el exterior de la célula está cargado positivamente con respecto al interior de la célula.
Este efecto electrogénico directo en la célula es mínimo ya que sólo contribuye a un 10 % del total del potencial eléctrico de la membrana celular. No obstante, casi todo el resto del potencial deriva indirectamente de la acción de la bomba de sodio y potasio, y se debe en su mayor parte al potencial de reposo para el potasio.
Impulsos nerviosos
La concentración intracelular de sodio es alrededor de 5 mM mientras que la extracelular es mucho mayor (145 mM). Sin embargo, las concentraciones intra y extracelulares de potasio son 140 mM y 5 mM respectivamente. Esto nos indica que hay un fuerte gradiente electroquímico que impulsa a las dos sustancias a moverse: el sodio hacia adentro y el potasio hacia afuera de la célula. Como la membrana es impermeable a estos solutos, controlando la entrada y salida de estas sustancias (principalmente), la célula genera cambios de concentración de iones a ambos lados de la membrana, y como los iones tienen carga eléctrica, también se modifica el potencial a través suyo. Combinando estos dos factores, las células de un organismo son capaces de transmitirse señales eléctricas (véase: potencial de acción) y comunicarse entre ellas, paso fundamental para la evolución del reino animal.
La bomba de Na+/K+ contribuye a equilibrar el potencial de membrana y mantener el potencial de reposo (es decir, las concentraciones constantes a ambos lados) cuando el impulso nervioso ya se ha transmitido. Este impulso nervioso hace que los canales de Na+ se abran generando un desequilibrio en la membrana y despolarizándola, debido a la entrada de sodio a favor de gradiente, que al ser un catión revierte localmente el estado de electronegatividad del lado interno de la membrana. Cuando el impulso ha pasado, los canales de Na+ se cierran y se abren los de K+, que implica la salida de potasio de la célula restaurando la electronegatividad intracelular. Para que el potencial de membrana sea normal la bomba de Na+/K+ funciona manteniendo las concentraciones de los iones constantes (expulsando el sodio que entra e introduciendo el potasio que sale).
Transducción de señales
Recientemente se ha descubierto que, independientemente de su función de transporte iónico, la bomba tiene una función como receptor de señales. Así, se ha descrito en miocitos de rata en cultivo una modificación en el ritmo de crecimiento tanto celular como mitótico cuando se añaden al medio análogos de ouabaína que actúan sobre la proteína. Este cambio no se debe a la modificación de las concentraciones iónicas sino a proteínas.
Farmacología
La bomba de sodio-potasio encontrada en las células del corazón es una diana importante para los glucósidos cardiacos (como digoxina y ouabaína), drogas inotrópicas positivas ampliamente usadas en la clínica para incrementar la fuerza de contracción.
Véase también
Bibliografía
- Hall, John E.; Guyton, Arthur C. (2007). Compendio de fisiología médica. Elsevier España. ISBN 9788480862325. Consultado el 6 de noviembre de 2017.
- Patton, Kevin T.; Thibodeau, Gary A. (2013). Anatomía y fisiología. Elsevier España. ISBN 9788490221082. Consultado el 6 de noviembre de 2017.
- Shoemaker (2002). Tratado de medicina crítica y terapia intensiva. Ed. Médica Panamericana. ISBN 9788479035877. Consultado el 6 de noviembre de 2017.
- Hill, Richard W.; Wyse, Gordon A.; Anderson, Margaret (2006). Fisiología animal. Ed. Médica Panamericana. ISBN 9788479039905. Consultado el 6 de noviembre de 2017.#Kalat, James W. (2004). Psicología biológica. Editorial Paraninfo. ISBN 9788497322850. Consultado el 6 de noviembre de 2017.
- B. Alberts y col. Biología Molecular de la Célula, 3ª ed., Editorial Omega, 1998, pp. 34-39, 78, 94.
- A Lehninger. Principios de Bioquímica. 3ª ed., Editorial Omega, 2001, pp. 67-69, 80, 84.
Referencias
- Arce, Víctor M. (27 de junio de 2006). OP/255-ENDOCRINOLOGIA. Univ Santiago de Compostela. ISBN 9788497506229. Consultado el 4 de septiembre de 2019.
- Pivovarov, Arkady S.; Calahorro, Fernando; Walker, Robert J. (noviembre de 2018). «Na+/K+-pump and neurotransmitter membrane receptors» [La bomba sodio-potasio y los receptores de membrana de neurotransmisores]. Mc viscent (en japones) (Springer) 19 (1): 1. PMC 6267510. PMID 30488358. doi:10.1007/s10158-018-0221-7. Consultado el 22 de diciembre de 2018.
- Skou J (1957). «The influence of some cations on an adenosine triphosphatase from peripheral nerves.». Biochim Biophys Acta 23 (2): 394-401. PMID 13412736. doi:10.1016/0006-3002(57)90343-8.
- Gonçalves-de-Albuquerque, Cassiano Felippe; Silva, Adriana Ribeiro; da Silva, Camila Ignácio; Castro-Faria-Neto, Hugo Caire; Burth, Patrícia (abril de 2017). «Na/K Pump and Beyond: Na/K-ATPase as a Modulator of Apoptosis and Autophagy» [Bomba sodio-potasio y mas allá: sodio-potasio ATPasa como un modulador de la apoptosis y autofagia]. Molecules (en inglés) (Multidisciplinary Digital Publishing Institute (MDPI)) 22 (4): 578. PMC 6154632. PMID 28430151. doi:10.3390/molecules22040578. Consultado el 22 de diciembre de 2018.
| Control de autoridades |
|
|---|
 Datos: Q59143
Datos: Q59143 Multimedia: Sodium-potassium-exchanging ATPase / Q59143
Multimedia: Sodium-potassium-exchanging ATPase / Q59143