Célula madre embrionaria
Las células madre embrionarias (CME) son células madre pluripotentes derivadas de la masa celular interna de un blastocisto, un embrión en etapa temprana antes de la implantación.[1][2] Los embriones humanos alcanzan la fase de blastocisto 4-5 días después de la fertilización, momento en el que constan de 50-150 células. El aislamiento de la masa celular interna (embrioblasto) mediante inmunocirugía provoca la destrucción del blastocisto, un proceso que plantea cuestiones éticas, como si los embriones en la fase de preimplantación tienen las mismas consideraciones morales que los embriones en la fase de desarrollo posterior a la implantación.[3][4]


Los investigadores se centran actualmente en el potencial terapéutico de las células madre embrionarias, y su uso clínico es el objetivo de muchos laboratorios.[2] Entre los posibles usos figuran el tratamiento de la diabetes y las cardiopatías.[2] Las células se están estudiando para utilizarlas como terapias clínicas, modelos de trastornos genéticos y reparación celular/ADN. Sin embargo, también se han notificado efectos adversos en la investigación y los procesos clínicos, como tumores y respuestas inmunitarias no deseadas.[5]
Propiedades

Las células madre embrionarias (CME), derivadas del estadio de blastocisto de embriones tempranos de mamíferos, se distinguen por su capacidad de diferenciarse en cualquier tipo de célula embrionaria y por su capacidad de autorrenovación. Son estas características las que las hacen valiosas en los campos científico y médico. Las CME tienen un cariotipo normal, mantienen una elevada actividad telomerasa y presentan un notable potencial proliferativo a largo plazo.[6]
Pluripotencia
Las células madre embrionarias de la masa celular interna son pluripotentes, lo que significa que pueden diferenciarse para generar ectodermo primitivo, que finalmente se diferencia durante la gastrulación en todos los derivados de las tres capas germinales primarias: ectodermo, endodermo y mesodermo. Estas capas germinales generan cada uno de los más de 220 tipos celulares del cuerpo humano adulto. Cuando reciben las señales adecuadas, las CME forman inicialmente células precursoras que posteriormente se diferencian en los tipos celulares deseados. La pluripotencia distingue a las células madre embrionarias de las células madre adultas, que son multipotentes y sólo pueden producir un número limitado de tipos celulares.
Auto renovación y reparación de estructura.
En condiciones definidas, las células madre embrionarias son capaces de autorrenovarse indefinidamente en un estado indiferenciado. Las condiciones de autorrenovación deben impedir que las células se agrupen y mantener un entorno que favorezca un estado no especializado.[7] Normalmente, esto se hace en el laboratorio con medios que contienen suero y factor inhibidor de la leucemia o suplementos de medios sin suero con dos fármacos inhibidores ("2i"), el inhibidor de MEK PD03259010 y el inhibidor de GSK-3 CHIR99021.[8]
Crecimiento
Las ESC se dividen con mucha frecuencia debido a una fase G1 acortada en su ciclo celular. La rápida división celular permite a las células crecer rápidamente en número, pero no en tamaño, lo que es importante para el desarrollo embrionario temprano. En las ESC, las proteínas ciclina A y ciclina E involucradas en la transición G1/S siempre se expresan en niveles altos.[9] Las quinasas dependientes de ciclina, como CDK2, que promueven la progresión del ciclo celular, son hiperactivas, en parte debido a la regulación a la baja de sus inhibidores.[10] Las proteínas del retinoblastoma que inhiben el factor de transcripción E2F hasta que la célula está lista para entrar en la fase S están hiperfosforiladas e inactivadas en las ESC, lo que lleva a la expresión continua de genes de proliferación.[9] Estos cambios dan lugar a ciclos acelerados de división celular. Aunque los altos niveles de expresión de proteínas pro-proliferativas y una fase G1 acortada se han relacionado con el mantenimiento de la pluripotencia,[11][12] las CME cultivadas en condiciones 2i libres de suero expresan proteínas Retinoblastoma activas hipofosforiladas y tienen una fase G1 alargada.[13] A pesar de esta diferencia en el ciclo celular, cuando se comparan con las CME cultivadas en medios que contienen suero, estas células tienen características pluripotentes similares.[14] Los factores de pluripotencia Oct4 y Nanog desempeñan un papel en la regulación transcripcional del ciclo de las células madre embrionarias.[15][16]
Usos
Debido a su plasticidad y a su capacidad potencialmente ilimitada de autorrenovación, se han propuesto terapias con células madre embrionarias para la medicina regenerativa y la sustitución de tejidos tras una lesión o enfermedad. Las células madre pluripotentes han demostrado ser prometedoras en el tratamiento de diversas afecciones como las lesiones de la médula espinal, degeneración macular relacionada con la edad, diabetes, trastornos neurodegenerativos (como la enfermedad de Parkinson ), SIDA, etc.[17] Además de su potencial en medicina regenerativa, las células madre embrionarias proporcionan una posible fuente alternativa de tejidos/órganos que sirve como posible solución al dilema de la escasez de donantes. Sin embargo, existen algunas controversias éticas al respecto (véase la sección Debate ético más adelante). Aparte de estos usos, las células madre embrionarias también pueden utilizarse para investigar el desarrollo humano temprano, ciertas enfermedades genéticas y pruebas toxicológicas in vitro.[6]
Utilización
Según un artículo de 2002 en PNAS, "Las células madre embrionarias humanas tienen el potencial de diferenciarse en varios tipos celulares y, por tanto, pueden ser útiles como fuente de células para trasplantes o ingeniería de tejidos".[18]
Ingeniería de tejidos

En la ingeniería de tejidos, el uso de células madre es conocido por su importancia. Para diseñar con éxito un tejido, las células utilizadas deben ser capaces de desempeñar funciones biológicas específicas como la secreción de citoquinas, moléculas de señalización, interactuar con células vecinas y producir una matriz extracelular en la organización correcta. Las células madre demuestran estas funciones biológicas específicas además de ser capaces de autorrenovarse y diferenciarse en uno o más tipos de células especializadas. Las células madre embrionarias son una de las fuentes que se están considerando para el uso de la ingeniería de tejidos.[19] El uso de células madre embrionarias humanas ha abierto muchas nuevas posibilidades para la ingeniería de tejidos, sin embargo, hay muchos obstáculos que deben superarse antes de que las células madre embrionarias humanas puedan ser utilizadas. Se teoriza que si las células madre embrionarias pueden ser alteradas para no evocar la respuesta inmune cuando se implantan en el paciente, entonces esto sería un paso revolucionario en la ingeniería de tejidos.[20] Las células madre embrionarias no se limitan a la ingeniería de tejidos.
Terapias de reemplazo celular
La investigación se ha centrado en la diferenciación de las células madre embrionarias en diversos tipos celulares para su posible uso en terapias de sustitución celular. Algunos de los tipos celulares que se han desarrollado o se están desarrollando actualmente son los cardiomiocitos, las neuronas, los hepatocitos, las células de la médula ósea, las células de los islotes y las células endoteliales.[21] Sin embargo, la derivación de estos tipos celulares a partir de CME no está exenta de obstáculos, por lo que la investigación se ha centrado en superar estas barreras. Por ejemplo, se están realizando estudios para diferenciar las CME en cardiomiocitos de tejidos específicos y erradicar sus propiedades inmaduras que las distinguen de los cardiomiocitos adultos.[22]
Potencial clínico
- Los investigadores han diferenciado las CME en células productoras de dopamina con la esperanza de que estas neuronas puedan utilizarse en el tratamiento de la enfermedad de Parkinson.[23][24]
- Las CME se han diferenciado en células asesinas naturales y tejido óseo.[25]
- Se están realizando estudios con células madre embrionarias para ofrecer un tratamiento alternativo de la diabetes. Por ejemplo, las células madre embrionarias se han diferenciado en células productoras de insulina,[26] y los investigadores de la Universidad de Harvard fueron capaces de producir grandes cantidades de células beta pancreáticas a partir de células madre embrionarias.[27]
- Un artículo publicado en el European Heart Journal describe un proceso traslacional de generación de células progenitoras cardíacas derivadas de células madre embrionarias humanas para su uso en ensayos clínicos de pacientes con insuficiencia cardíaca grave.[28]
Descubrimiento de medicamentos
Además de convertirse en una importante alternativa a los trasplantes de órganos, las CME también se están utilizando en el campo de la toxicología y como cribas celulares para descubrir nuevas entidades químicas que puedan desarrollarse como fármacos de moléculas pequeñas. Los estudios han demostrado que los cardiomiocitos derivados de CME son modelos in vitro validados para probar las respuestas a fármacos y predecir los perfiles de toxicidad.[21] Se ha demostrado que los cardiomiocitos derivados de CME responden a estímulos farmacológicos y, por tanto, pueden utilizarse para evaluar la cardiotoxicidad, como las torsades de pointes.[29]
Los hepatocitos derivados de CME también son modelos útiles que podrían utilizarse en las fases preclínicas del descubrimiento de fármacos. Sin embargo, el desarrollo de hepatocitos a partir de CME ha demostrado ser un reto, lo que dificulta la capacidad de probar el metabolismo de los fármacos. Por lo tanto, la investigación se ha centrado en el establecimiento de hepatocitos derivados de CME plenamente funcionales con actividad enzimática de fase I y II estable.[30]
Modelos de trastornos genéticos
Varios estudios nuevos han empezado a abordar el concepto de modelización de trastornos genéticos con células madre embrionarias. Ya sea manipulando genéticamente las células o, más recientemente, derivando líneas celulares enfermas identificadas mediante diagnóstico genético prenatal (DGP), el modelado de trastornos genéticos es algo que se ha logrado con células madre. Este enfoque puede resultar muy valioso para estudiar trastornos como el síndrome X frágil, la fibrosis quística y otras enfermedades genéticas que carecen de un sistema modelo fiable.
Yury Verlinsky, investigador médico ruso-estadounidense especializado en genética embrionaria y celular (citología genética), desarrolló métodos de prueba de diagnóstico prenatal para determinar trastornos genéticos y cromosómicos un mes y medio antes que la amniocentesis estándar. Estas técnicas las utilizan ahora muchas embarazadas y futuros padres, sobre todo parejas con antecedentes de anomalías genéticas o en las que la mujer tiene más de 35 años (cuando el riesgo de trastornos relacionados con la genética es mayor). Además, al permitir a los padres seleccionar un embrión sin trastornos genéticos, tienen la posibilidad de salvar la vida de hermanos que ya padecían trastornos y enfermedades similares utilizando células de la descendencia libre de enfermedades.[31]
Reparación del daño del ADN
Las células somáticas diferenciadas y las células madre embrionarias utilizan diferentes estrategias para tratar los daños en el ADN. Por ejemplo, los fibroblastos de prepucio humano, un tipo de célula somática, utilizan la unión de extremos no homólogos, un proceso de reparación del ADN propenso a errores, como vía principal para reparar las roturas de doble cadena durante todas las fases del ciclo celular.[32] Debido a su naturaleza propensa a errores, la unión de extremos no homólogos tiende a producir mutaciones en los descendientes clonales de una célula.
Las células madre embrionarias utilizan una estrategia diferente para tratar las roturas de doble cadena.[33] Dado que las células madre embrionarias dan lugar a todos los tipos celulares de un organismo, incluidas las células de la línea germinal, las mutaciones que surgen en las células madre embrionarias debido a una reparación defectuosa del ADN constituyen un problema más grave que en las células somáticas diferenciadas. En consecuencia, se necesitan mecanismos robustos en las células ES para reparar con precisión los daños en el ADN y, si la reparación falla, eliminar aquellas células con daños en el ADN no reparados. Así, las células madre embrionarias de ratón utilizan predominantemente la reparación recombinacional homóloga de alta fidelidad (RRH) para reparar las roturas de doble cadena.[33] Este tipo de reparación depende de la interacción de los dos cromosomas hermanos formados durante la fase S y presentes juntos durante la fase G2 del ciclo celular. La reparación recombinacional homóloga puede reparar con precisión las roturas de doble cadena en un cromosoma hermano utilizando información intacta del otro cromosoma hermano. Las células en la fase G1 del ciclo celular (es decir, después de la metafase/división celular pero antes de la siguiente ronda de replicación) sólo tienen una copia de cada cromosoma (es decir, no hay cromosomas hermanos). Las células madre embrionarias de ratón carecen de un punto de control G1 y no sufren una detención del ciclo celular al sufrir daños en el ADN.[34] En su lugar, sufren una muerte celular programada (apoptosis) en respuesta al daño en el ADN.[35] La apoptosis puede utilizarse como estrategia a prueba de fallos para eliminar las células con daños en el ADN no reparados con el fin de evitar mutaciones y la progresión a cáncer.[36] De acuerdo con esta estrategia, la frecuencia de mutación de las células madre embrionarias de ratón es 100 veces inferior a la de las células somáticas de ratón isogénicas.[37]
Ensayos clínicos
El 23 de enero de 2009, los ensayos clínicos de fase I para el trasplante de oligodendrocitos (un tipo de célula del cerebro y la médula espinal) derivados de CME humanas en personas con lesiones medulares recibieron la aprobación de la Administración de Alimentos y Medicamentos de EE.UU. (FDA), convirtiéndose así en el primer ensayo en humanos con CME humanas del mundo.[38] El estudio que condujo a este avance científico fue dirigido por Hans Keirstead y sus colegas de la Universidad de California en Irvine y contó con el apoyo de Geron Corporation de Menlo Park, California, fundada por Michael D. West, PhD. Un experimento anterior había demostrado una mejora de la recuperación locomotora en ratas lesionadas de la médula espinal tras un trasplante diferido de 7 días de CME humanas que habían sido forzadas hacia un linaje oligodendrocítico.[39] El estudio clínico de fase I se diseñó para inscribir a unos ocho o diez parapléjicos lesionados no más de dos semanas antes del inicio del ensayo, ya que las células deben inyectarse antes de que pueda formarse tejido cicatricial. Los investigadores subrayaron que no se esperaba que las inyecciones curaran por completo a los pacientes y les devolvieran toda la movilidad. Basándose en los resultados de los ensayos con roedores, los investigadores especularon con la posibilidad de restaurar las vainas de mielina y aumentar la movilidad. Este primer ensayo se diseñó principalmente para comprobar la seguridad de estos procedimientos y, si todo iba bien, se esperaba que condujera a futuros estudios en los que participaran personas con discapacidades más graves.[40] El ensayo se suspendió en agosto de 2009 debido a la preocupación de la FDA por un pequeño número de quistes microscópicos hallados en varios modelos de rata tratados, pero la suspensión se levantó el 30 de julio de 2010.[41]
En octubre de 2010, los investigadores inscribieron y administraron CME al primer paciente en el Shepherd Center de Atlanta.[42] Los fabricantes de la terapia con células madre, Geron Corporation, estimaron que se tardarían varios meses en replicar las células madre y en evaluar el éxito o el fracaso de la terapia GRNOPC1.
En noviembre de 2011, Geron anunció que interrumpía el ensayo y abandonaba la investigación con células madre por motivos económicos, pero que seguiría controlando a los pacientes existentes e intentaba encontrar un socio que pudiera continuar su investigación.[43] En 2013, BioTime, dirigida por su consejero delegado, el Dr. Michael D. West, adquirió todos los activos de células madre de Geron, con la intención declarada de reiniciar el ensayo clínico basado en células madre embrionarias de Geron para la investigación de lesiones medulares.[44]
Asterias Biotherapeutics (NYSE MKT: AST), empresa de BioTime, ha recibido del Instituto de Medicina Regenerativa de California (CIRM) una subvención de 14,3 millones de dólares para reiniciar el primer ensayo clínico en humanos con células madre embrionarias para lesiones medulares. El CIRM, financiado con fondos públicos de California, es el mayor financiador de investigación y desarrollo con células madre del mundo.
El premio proporciona financiación para que Asterias reinicie el desarrollo clínico de AST-OPC1 en sujetos con lesión medular y amplíe las pruebas clínicas de dosis crecientes en la población diana prevista para futuros ensayos pivotales.
AST-OPC1 es una población de células derivadas de células madre embrionarias humanas (hESCs) que contiene células progenitoras de oligodendrocitos (OPCs). Las OPC y sus derivados maduros, denominados oligodendrocitos, proporcionan un soporte funcional crítico para las células nerviosas de la médula espinal y el cerebro. Asterias presentó recientemente los resultados de un ensayo clínico de fase 1 en el que se probó una dosis baja de AST-OPC1 en pacientes con lesión neurológica completa de la médula espinal torácica. Los resultados mostraron que AST-OPC1 se administró con éxito en la zona de la médula espinal lesionada. Los pacientes seguidos 2-3 años después de la administración de AST-OPC1 no mostraron evidencias de acontecimientos adversos graves asociados a las células en evaluaciones de seguimiento detalladas que incluían exámenes neurológicos y resonancias magnéticas frecuentes. La monitorización inmunológica de los sujetos hasta un año después del trasplante no mostró evidencias de respuestas inmunológicas celulares o basadas en anticuerpos a AST-OPC1. En cuatro de los cinco sujetos, las resonancias magnéticas seriadas realizadas a lo largo del período de seguimiento de 2-3 años indican que puede haberse producido una reducción de la cavitación de la médula espinal y que AST-OPC1 puede haber tenido algunos efectos positivos en la reducción del deterioro del tejido de la médula espinal. No se produjo ninguna degeneración o mejora neurológica inesperada en los cinco sujetos del ensayo, según la evaluación del examen ISNCSCI (International Standards for Neurological Classification of Spinal Cord Injury).
La subvención de la Asociación Estratégica III del CIRM proporcionará financiación a Asterias para apoyar el próximo ensayo clínico de AST-OPC1 en sujetos con lesión medular, y para el desarrollo de productos de Asterias para perfeccionar y ampliar los métodos de fabricación para apoyar los ensayos en etapas posteriores y, finalmente, la comercialización. La financiación del CIRM estará condicionada a la aprobación del ensayo por parte de la FDA, a la conclusión de un acuerdo definitivo entre Asterias y el CIRM, y al progreso continuado de Asterias hacia la consecución de determinados hitos predefinidos del proyecto.
Preocupación y controversia
Efectos adversos
La principal preocupación que suscita el posible trasplante de CME a pacientes como terapia es su capacidad para formar tumores, incluidos teratomas.[45] Los problemas de seguridad llevaron a la FDA a suspender el primer ensayo clínico con células madre embrionarias, aunque no se observaron tumores. La principal estrategia para mejorar la seguridad de las CME para su posible uso clínico consiste en diferenciar las CME en tipos celulares específicos (por ejemplo, neuronas, músculo, células hepáticas) que tengan una capacidad reducida o eliminada de causar tumores. Tras la diferenciación, las células se clasifican por citometría de flujo para su posterior purificación. Se prevé que las CME sean intrínsecamente más seguras que las células iPS creadas con vectores virales de integración genética, ya que no están modificadas genéticamente con genes como c-Myc, relacionados con el cáncer. No obstante, las CME expresan niveles muy elevados de los genes inductores de iPS y estos genes, incluido Myc, son esenciales para la autorrenovación y la pluripotencia[46] de las CME, por lo que es poco probable que las posibles estrategias para mejorar la seguridad mediante la eliminación de la expresión de c-Myc preserven la "troncalidad" de las células. Sin embargo, se han identificado N-myc y L-myc para inducir células iPS en lugar de c-myc con una eficacia similar.[47] Los protocolos posteriores para inducir la pluripotencia eluden completamente estos problemas mediante el uso de vectores virales de ARN no integradores, como el virus Sendai o la transfección de ARNm.
Debate ético
Debido a la naturaleza de la investigación con células madre embrionarias, hay muchas opiniones controvertidas sobre el tema. Dado que la extracción de células madre embrionarias suele requerir la destrucción del embrión del que se obtienen dichas células, se cuestiona el estatus moral del embrión. Algunas personas afirman que la masa de células de 5 días es demasiado joven para alcanzar la personalidad o que el embrión, si se dona de una clínica de fecundación in vitro (donde los laboratorios suelen adquirir embriones), iría de todos modos a la basura médica. Los que se oponen a la investigación con CME afirman que un embrión es una vida humana, por lo que destruirlo es un asesinato y el embrión debe ser protegido bajo el mismo punto de vista ético que un ser humano más desarrollado.[48]
Historia
- 1964: Lewis Kleinsmith y G. Barry Pierce Jr. aislaron un único tipo de célula de un teratocarcinoma, un tumor que ahora se conoce como célula germinal.[49] Estas células aisladas del teratocarcinoma se replicaron y crecieron en cultivo celular como una célula madre y ahora se conocen como células de carcinoma embrionario (CE). [Aunque las similitudes en morfología y potencial diferenciador (pluripotencia) condujeron al uso de células CE como modelo in vitro para el desarrollo temprano del ratón,[50] las células CE albergan mutaciones genéticas y a menudo cariotipos anormales que se acumularon durante el desarrollo del teratocarcinoma. Estas aberraciones genéticas acentuaron aún más la necesidad de poder cultivar células pluripotentes directamente a partir de la masa celular interna.

- 1981: Dos grupos obtienen por primera vez células madre embrionarias CME cells) a partir de embriones de ratón. Martin Evans y Matthew Kaufman, del Departamento de Genética de la Universidad de Cambridge, publicaron el primero en julio, en el que revelaban una nueva técnica de cultivo de embriones de ratón en el útero que permitía aumentar el número de células, lo que posibilitaba la derivación de células madre embrionarias a partir de estos embriones.[51] Gail R. Martin, del Departamento de Anatomía de la Universidad de California en San Francisco, publicó su trabajo en diciembre y acuñó el término "célula madre embrionaria".[52] Demostró que se podían cultivar embriones in vitro y que se podían derivar células madre embrionarias de estos embriones.
- 1989: Mario R. Cappechi, Martin J. Evans y Oliver Smithies publican su investigación en la que detallan su aislamiento y modificaciones genéticas de células madre embrionarias, creando los primeros "ratones knockout".[53] Al crear ratones knockout, esta publicación proporcionó a los científicos una forma totalmente nueva de estudiar las enfermedades.
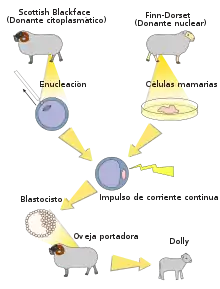
- 1996: Dolly, fue el primer mamífero clonado a partir de una célula adulta por el Instituto Roslin de la Universidad de Edimburgo.[54] Este experimento instituyó la proposición de que las células adultas especializadas obtienen la composición genética para realizar una tarea específica; lo que estableció una base para investigaciones posteriores dentro de una variedad de técnicas de clonación. El experimento Dolly se llevó a cabo obteniendo las células de la ubre de una oveja (Dolly) y diferenciándolas hasta que se completó la división. A continuación, se obtuvo un óvulo de una oveja huésped diferente y se le extrajo el núcleo. Se colocó una célula de la ubre junto al óvulo y se conectaron eléctricamente para que compartieran el ADN. Este óvulo se diferenció en un embrión y el embrión se insertó en una tercera oveja que dio a luz a la versión clónica de Dolly.[55]
- 1998: Un equipo de la Universidad de Wisconsin, Madison (James A. Thomson, Joseph Itskovitz-Eldor, Sander S. Shapiro, Michelle A. Waknitz, Jennifer J. Swiergiel, Vivienne S. Marshall y Jeffrey M. Jones) publica un artículo titulado "Líneas de células madre embrionarias derivadas de blastocitos humanos". Los investigadores responsables de este estudio no sólo crearon las primeras células madre embrionarias, sino que reconocieron su pluripotencia, así como su capacidad de autorrenovación. El resumen del artículo destaca la importancia del descubrimiento para los campos de la biología del desarrollo y el descubrimiento de fármacos.[56]
- 2001: El Presidente George W. Bush permite la financiación federal para apoyar la investigación en aproximadamente 60 -en ese momento, ya existentes- líneas de células madre embrionarias. Dado que las líneas limitadas sobre las que Bush permitió la investigación ya habían sido establecidas, esta ley apoyó la investigación con células madre embrionarias sin plantear las cuestiones éticas que podrían surgir con la creación de nuevas líneas con cargo al presupuesto federal.[57]
- 2006: Los científicos japoneses Shinya Yamanaka y Kazutoshi Takashi publican un artículo que describe la inducción de células madre pluripotentes a partir de cultivos de fibroblastos de ratón adulto. Las células madre pluripotentes inducidas (iPSC) son un gran descubrimiento, ya que aparentemente son idénticas a las células madre embrionarias y podrían usarse sin provocar la misma controversia moral.[58]
- Enero de 2009: La Administración de Drogas y Alimentos de los EE . UU. (FDA) otorga la aprobación para el ensayo de fase I de Geron Corporation de su tratamiento derivado de células madre embrionarias humanas para lesiones de la médula espinal. El anuncio fue recibido con entusiasmo por la comunidad científica, pero también con cautela por parte de los opositores a las células madre. Sin embargo, las células de tratamiento se derivaron de las líneas celulares aprobadas bajo la política ESC de George W. Bush.[59]
- Marzo de 2009: El presidente Barack Obama firma la Orden Ejecutiva 13505, que elimina las restricciones impuestas a la financiación federal para células madre humanas por la administración presidencial anterior. Esto permitiría a los Institutos Nacionales de Salud (NIH) proporcionar fondos para la investigación de hESC. El documento también establece que los NIH deben proporcionar directrices de financiación federal revisadas en un plazo de 120 días a partir de la firma de la orden.[60]
Técnicas y condiciones de obtención y cultivo
Obtención de humanos
La fecundación in vitro genera múltiples embriones. El excedente de embriones no se utiliza clínicamente o no es apto para la implantación en la paciente, por lo que puede ser donado por la donante con su consentimiento. Las células madre embrionarias humanas pueden derivarse de estos embriones donados o, además, también pueden extraerse de embriones clonados creados a partir de una célula de un paciente y un óvulo donado mediante el proceso de transferencia nuclear de células somáticas.[61] La masa celular interna (células de interés), procedente del estadio de blastocisto del embrión, se separa del trofectodermo, las células que se diferenciarían en tejido extraembrionario. Para lograr la separación se realiza inmunocirugía, proceso en el que los anticuerpos se unen al trofectodermo y se eliminan mediante otra solución, y disección mecánica. Las células resultantes de la masa celular interna se colocan en placas sobre células que servirán de soporte. Las células de la masa celular interna se adhieren y se expanden para formar una línea celular embrionaria humana indiferenciada. Estas células se alimentan diariamente y se separan enzimática o mecánicamente cada cuatro o siete días. Para que se produzca la diferenciación, la línea de células madre embrionarias humanas se separa de las células de soporte para formar cuerpos embrioides, se co-cultiva con un suero que contenga las señales necesarias, o se injerta en un armazón tridimensional para obtener resultados.[62]
Obtención de otros animales.
Las células madre embrionarias se derivan de la masa celular interna del embrión temprano, que se recolectan del animal madre donante. Martin Evans y Matthew Kaufman presentaron una técnica que retrasa la implantación del embrión y permite que aumente la masa celular interna. Este proceso incluye la extirpación de los ovarios de la madre donante y su dosificación con progesterona, cambiando el entorno hormonal, lo que hace que los embriones permanezcan libres en el útero. Tras 4-6 días de cultivo intrauterino, los embriones se cosechan y se cultivan in vitro hasta que la masa celular interna forma "estructuras similares a cilindros de huevo", que se disocian en células individuales y se colocan en placas sobre fibroblastos tratados con mitomicina-c (para impedir la mitosis de los fibroblastos). Las líneas celulares clonales se crean al hacer crecer una sola célula. Evans y Kaufman demostraron que las células que crecían a partir de estos cultivos podían formar teratomas y cuerpos embrioides y diferenciarse in vitro, todo lo cual indica que las células son pluripotentes.[51]
Gail Martin obtuvo y cultivó sus células madre embrionarias de forma diferente. Extrajo los embriones de la madre donante aproximadamente 76 horas después de la cópula y los cultivó durante la noche en un medio que contenía suero. Al día siguiente, extrajo la masa celular interna del blastocisto tardío mediante microcirugía. La masa celular interna extraída se cultivó en fibroblastos tratados con mitomicina-c en un medio que contenía suero y acondicionado con células madre embrionarias. Al cabo de aproximadamente una semana, crecieron colonias de células. Estas células crecían en cultivo y mostraban características pluripotentes, como demostraba la capacidad de formar teratomas, diferenciarse in vitro y formar cuerpos embrioides. Martin se refirió a estas células como células ES.[52]
Ahora se sabe que las células alimentadoras proporcionan factor inhibidor de la leucemia (LIF) y el suero proporciona proteínas morfogenéticas óseas (BMP) que son necesarias para evitar que las células ES se diferencien.[63][64] Estos factores son extremadamente importantes para la eficacia de la derivación de células madre embrionarias. Además, se ha demostrado que diferentes cepas de ratón tienen diferentes eficiencias para aislar células madre embrionarias.[65] Los usos actuales de las células madre embrionarias de ratón incluyen la generación de ratones transgénicos, como los ratones knockout. Para el tratamiento humano, se necesitan células pluripotentes específicas para cada paciente. La generación de células madre embrionarias humanas es más difícil y plantea problemas éticos. Por eso, además de la investigación con células madre embrionarias humanas, muchos grupos se centran en la generación decélulas madre pluripotentes inducidas (células) iPS).[66]
Métodos potenciales para la derivación de nuevas líneas celulares
El 23 de agosto de 2006, la edición electrónica de la revista científica Nature publicó una carta del Dr. Robert Lanza (director médico de Advanced Cell Technology en Worcester, MA) en la que afirmaba que su equipo había encontrado una forma de extraer células madre embrionarias sin destruir el embrión real.[67] Este logro técnico permitiría a los científicos trabajar con nuevas líneas de células madre embrionarias obtenidas con fondos públicos en Estados Unidos, donde la financiación federal estaba limitada en aquel momento a la investigación con líneas de células madre embrionarias obtenidas antes de agosto de 2001. En marzo de 2009 se levantó esta limitación.[68]
Las células madre embrionarias humanas también se han obtenido mediante transferencia nuclear de células somáticas (TNCS).[69][70] Este método también se ha denominado a veces "clonación terapéutica" porque la TNCS se parece a otros tipos de clonación en que los núcleos se transfieren de una célula somática a un cigoto enucleado. Sin embargo, en este caso la TNCS se utilizó para producir líneas de células madre embrionarias en un laboratorio, no organismos vivos a través de un embarazo. La parte "terapéutica" del nombre se incluye por la esperanza de que las células madre embrionarias producidas mediante TNCS puedan tener utilidad clínica.
Células madre pluripotentes inducidas
La tecnología de las células iPS fue impulsada por el laboratorio de Shinya Yamanaka en Kioto (Japón), quien demostró en 2006 que la introducción de cuatro genes específicos que codifican factores de transcripción podía convertir células adultas en células madre pluripotentes.[71] Fue galardonado con el Premio Nobel 2012 junto con Sir John Gurdon "por el descubrimiento de que las células maduras pueden reprogramarse para convertirse en pluripotentes."[72]
En 2007, se demostró que las células madre pluripotentes, muy similares a las células madre embrionarias, pueden inducirse mediante la administración de cuatro factores (Oct3/4, Sox2, c-Myc y Klf4) a células diferenciadas.[73] Utilizando los cuatro genes enumerados anteriormente, las células diferenciadas se "reprograman" en células madre pluripotentes, lo que permite la generación de células madre pluripotentes/embrionarias sin el embrión. La morfología y los factores de crecimiento de estas células pluripotentes inducidas en laboratorio, son equivalentes a las células madre embrionarias, lo que hace que estas células se conozcan como células madre pluripotentes inducidas (células iPS).[71] Esta observación se realizó originalmente en células madre pluripotentes de ratón, pero ahora puede realizarse en fibroblastos humanos adultos utilizando los mismos cuatro genes.[74]
Dado que las preocupaciones éticas en torno a las células madre embrionarias suelen referirse a que derivan de embriones abortados, se cree que la reprogramación a estas células iPS puede ser menos controvertida.
Esto puede permitir la generación de líneas de células madre embrionarias específicas de pacientes que podrían utilizarse para terapias de sustitución celular. Además, esto permitirá generar líneas de células madre embrionarias de pacientes con diversas enfermedades genéticas y proporcionará modelos inestimables para estudiar esas enfermedades.
Sin embargo, como primer indicio de que la tecnología de las células iPS puede conducir en rápida sucesión a nuevas curas, un equipo de investigación dirigido por Rudolf Jaenisch, del Instituto Whitehead para la Investigación Biomédica en Cambridge, Massachusetts, para curar ratones con anemia de células falciformes, según lo informado por la edición en línea de la revista Science el 6 de diciembre de 2007.[75][76]
El 16 de enero de 2008, una empresa californiana, Stemagen, anunció que había creado los primeros embriones humanos clonados maduros a partir de células cutáneas individuales extraídas de adultos. Estos embriones pueden extraerse para obtener células madre embrionarias compatibles con el paciente.[77]
Contaminación por reactivos utilizados en el cultivo celular
La edición en línea de Nature Medicine publicó un estudio el 24 de enero de 2005 en el que se afirmaba que las células madre embrionarias humanas disponibles para la investigación financiada con fondos federales están contaminadas con moléculas no humanas procedentes del medio de cultivo utilizado para cultivar las células.[78] Es una técnica habitual utilizar células de ratón y de otros animales para mantener la pluripotencia de las células madre en división activa. El problema se descubrió cuando se comprobó que la presencia de ácido siálico no humano en el medio de cultivo comprometía los posibles usos de las células madre embrionarias en humanos, según científicos de la Universidad de California en San Diego.[79]
Sin embargo, un estudio publicado en la edición electrónica de Lancet Medical Journal el 8 de marzo de 2005, detallaba información sobre una nueva línea de células madre derivadas de embriones humanos en condiciones completamente libres de células y suero. Tras más de 6 meses de proliferación indiferenciada, estas células demostraron su potencial para formar derivados de las tres capas germinales embrionarias tanto in vitro como en teratomas. Estas propiedades también se mantuvieron con éxito (durante más de 30 pasajes) con las líneas de células madre establecidas.[80]
Células de Muse
Las células de Muse (células resistentes al estrés de diferenciación de múltiples linajes) son células madre pluripotentes no cancerosas que se encuentran en adultos.[81][82] Fueron descubiertss en 2010 por Mari Dezawa y su equipo de investigación.[81] Las células Muse residen en el tejido conectivo de casi todos los órganos, incluido el cordón umbilical, la médula ósea y la sangre periférica.[83][81][84][85][86] Se obtienen a partir de células mesenquimales comercializadas, como fibroblastos humanos, células madre mesenquimales de médula ósea y células madre derivadas de tejido adiposo.[87][88][89] Las células Muse son capaces de generar células representativas de las tres capas germinales a partir de una única célula, tanto de forma espontánea como bajo inducción con citoquinas. La expresión de genes de pluripotencia y la diferenciación triploblástica son autorrenovables a lo largo de generaciones. Las células Muse no sufren la formación de teratomas cuando se trasplantan a un entorno huésped in vivo, lo que erradica el riesgo de tumorigénesis por proliferación celular desenfrenada.[81]
Referencias
- Thomson; Itskovitz-Eldor, J; Shapiro, SS; Waknitz, MA; Swiergiel, JJ; Marshall, VS; Jones, JM (1998). «Blastocysts Embryonic Stem Cell Lines Derived from Human». Science 282 (5391): 1145-1147. Bibcode:1998Sci...282.1145T. PMID 9804556. doi:10.1126/science.282.5391.1145.
- «Stem Cell Basics | STEM Cell Information». stemcells.nih.gov. Consultado el 5 de junio de 2022.
- Baldwing A (2009). «Morality and human embryo research. Introduction to the Talking Point on morality and human embryo research.». EMBO Reports 10 (4): 299-300. PMC 2672902. PMID 19337297. doi:10.1038/embor.2009.37.
- Nakaya, Andrea C. (1 de agosto de 2011). Biomedical ethics. San Diego, CA: ReferencePoint Press. pp. 96. ISBN 978-1601521576.
- Carla A Herberts; Marcel SG Kwa; Harm PH Hermsen (2011). «Risk factors in the development of stem cell therapy». Journal of Translational Medicine 9 (29): 29. PMC 3070641. PMID 21418664. doi:10.1186/1479-5876-9-29.
- Thomson, J. A.; Itskovitz-Eldor, J; Shapiro, S. S.; Waknitz, M. A.; Swiergiel, J. J.; Marshall, V. S.; Jones, J. M. (1998). «Embryonic Stem Cell Lines Derived from Human Blastocysts». Science 282 (5391): 1145-7. Bibcode:1998Sci...282.1145T. PMID 9804556. doi:10.1126/science.282.5391.1145.
- Ying; Nichols, J; Chambers, I; Smith, A (2003). «BMP Induction of Id Proteins Suppresses Differentiation and Sustains Embryonic Stem Cell Self-Renewal in Collaboration with STAT3». Cell 115 (3): 281-292. PMID 14636556. S2CID 7201396. doi:10.1016/S0092-8674(03)00847-X.
- Martello, G.; Smith, A. (2014). «The nature of embryonic stem cells». Annual Review of Cell and Developmental Biology 30: 647-75. PMID 25288119. doi:10.1146/annurev-cellbio-100913-013116.
- Boward, B.; Wu, T.; Dalton, S. (2016). «Concise Review: Control of Cell Fate Through Cell Cycle and Pluripotency Networks». Stem Cells 34 (6): 1427-36. PMC 5201256. PMID 26889666. doi:10.1002/stem.2345.
- White, J.; Stead, E.; Faast, R.; Conn, S.; Cartwright, P.; Dalton, S. (2005). «Developmental activation of the Rb-E2F pathway and establishment of cell cycle-regulated cyclin-dependent kinase activity during embryonic stem cell differentiation». Molecular Biology of the Cell 16 (4): 2018-27. PMC 1073679. PMID 15703208. doi:10.1091/mbc.e04-12-1056.
- Ter Huurne, Menno; Stunnenberg, Hendrik G. (21 de abril de 2021). «G1-phase progression in pluripotent stem cells». Cellular and Molecular Life Sciences 21 (10): 4507-4519. ISSN 1875-9777. PMC 8195903. PMID 33884444. doi:10.1007/s00018-021-03797-8.
- Singh, Amar M.; Dalton, Stephen (7 de agosto de 2009). «The cell cycle and Myc intersect with mechanisms that regulate pluripotency and reprogramming». Cell Stem Cell 5 (2): 141-149. ISSN 1875-9777. PMC 2909475. PMID 19664987. doi:10.1016/j.stem.2009.07.003.
- Ter Huurne, Menno; Chappell, James; Dalton, Stephen; Stunnenberg, Hendrik G. (5 de octubre de 2017). «Distinct Cell-Cycle Control in Two Different States of Mouse Pluripotency». Cell Stem Cell 21 (4): 449-455.e4. ISSN 1875-9777. PMC 5658514. PMID 28985526. doi:10.1016/j.stem.2017.09.004.
- Ying, Qi-Long; Wray, Jason; Nichols, Jennifer; Batlle-Morera, Laura; Doble, Bradley; Woodgett, James; Cohen, Philip; Smith, Austin (22 de mayo de 2008). «The ground state of embryonic stem cell self-renewal». Nature 453 (7194): 519-523. Bibcode:2008Natur.453..519Y. ISSN 1476-4687. PMC 5328678. PMID 18497825. doi:10.1038/nature06968.
- Lee, J.; Go, Y.; Kang, I.; Han, Y. M.; Kim, J. (2010). «Oct-4 controls cell-cycle progression of embryonic stem cells». The Biochemical Journal 426 (2): 171-81. PMC 2825734. PMID 19968627. doi:10.1042/BJ20091439.
- Zhang, X.; Neganova, I.; Przyborski, S.; Yang, C.; Cooke, M.; Atkinson, S. P.; Anyfantis, G.; Fenyk, S. et al. (2009). «A role for NANOG in G1 to S transition in human embryonic stem cells through direct binding of CDK6 and CDC25A». The Journal of Cell Biology 184 (1): 67-82. PMC 2615089. PMID 19139263. doi:10.1083/jcb.200801009.
- Mahla, Ranjeet (19 de julio de 2016). «Stem Cell Applications in Regenerative Medicine and Disease Therapeutics». International Journal of Cell Biology 2016: 6940283. PMC 4969512. PMID 27516776. doi:10.1155/2016/6940283.
- Levenberg, S. (2002). «Endothelial cells derived from human embryonic stem cells». Proceedings of the National Academy of Sciences 99 (7): 4391-4396. Bibcode:2002PNAS...99.4391L. PMC 123658. PMID 11917100. doi:10.1073/pnas.032074999.
- Vats, A; Tolley, N S; Bishop, A E; Polak, J M (1 de agosto de 2005). «Embryonic Stem Cells and Tissue Engineering: Delivering Stem Cells to the Clinic». Journal of the Royal Society of Medicine (en inglés) 98 (8): 346-350. ISSN 0141-0768. PMC 1181832. PMID 16055897. doi:10.1177/014107680509800804.
- Heath, Carole A. (1 de enero de 2000). «Cells for tissue engineering». Trends in Biotechnology (en inglés) 18 (1): 17-19. ISSN 0167-7799. PMID 10631775. doi:10.1016/S0167-7799(99)01396-7.
- Davila, JC; Cezar, GG; Thiede, M; Strom, S; Miki, T; Trosko, J (2004). «Use and application of stem cells in toxicology». Toxicological Sciences 79 (2): 214-223. PMID 15014205. doi:10.1093/toxsci/kfh100.
- Siu, CW; Moore, JC; Li, RA (2007). «Human embryonic stem cell-derived cardiomyocytes for heart therapies». Cardiovascular & Hematological Disorders Drug Targets 7 (2): 145-152. PMID 17584049. doi:10.2174/187152907780830851.
- Perrier, A. L. (2004). «Derivation of midbrain dopamine neurons from human embryonic stem cells». Proceedings of the National Academy of Sciences 101 (34): 12543-12548. Bibcode:2004PNAS..10112543P. PMC 515094. PMID 15310843. doi:10.1073/pnas.0404700101.
- Parish, CL; Arenas, E (2007). «Stem-cell-based strategies for the treatment of Parkinson's disease». Neuro-Degenerative Diseases 4 (4): 339-347. PMID 17627139. doi:10.1159/000101892.
- Waese, EY; Kandel, RA; Stanford, WL (2008). «Application of stem cells in bone repair». Skeletal Radiology 37 (7): 601-608. PMID 18193216. doi:10.1007/s00256-007-0438-8.
- d'Amour, KA; Bang, AG; Eliazer, S; Kelly, OG; Agulnick, AD; Smart, NG; Moorman, MA; Kroon, E; Carpenter, MK; Baetge, EE (2006). «Production of pancreatic hormone-expressing endocrine cells from human embryonic stem cells». Nature Biotechnology 24 (11): 1392-1401. PMID 17053790. S2CID 11040949. doi:10.1038/nbt1259.
- Colen, B.D. (9 de octubre de 2014) Giant leap against diabetes The Harvard Gazette, Recuperado el 24 de noviembre de 2014
- Menasché, Phillip; Vanneaux, Valérie; Fabreguettes, Jean-Roch; Bel, Alain; Tosca, Lucie; Garcia, Sylvie (21 de marzo de 2015). «Towards a clinical use of human embryonic stem cell derived-cardiac progenitors: a translational experience». European Heart Journal 36 (12): 743-750. PMID 24835485. doi:10.1093/eurheartj/ehu192.
- Jensen, J; Hyllner, J; Björquist, P (2009). «Human embryonic stem cell technologies and drug discovery». Journal of Cellular Physiology 219 (3): 513-519. PMID 19277978. doi:10.1002/jcp.21732.
- Söderdahl, T; Küppers-Munther, B; Heins, N; Edsbagge, J; Björquist, P; Cotgreave, I; Jernström, B (2007). «Glutathione transferases in hepatocyte-like cells derived from human embryonic stem cells». Toxicology in Vitro 21 (5): 929-937. PMID 17346923. doi:10.1016/j.tiv.2007.01.021.
- "Dr. Yury Verlinsky, 1943–2009: Expert in reproductive technology" Archivado el 8 de agosto de 2009 en Wayback Machine. Chicago Tribune, 20 de julio de 2009
- «DNA repair by nonhomologous end joining and homologous recombination during cell cycle in human cells». Cell Cycle 7 (18): 2902-2906. Septiembre de 2008. PMC 2754209. PMID 18769152. doi:10.4161/cc.7.18.6679.
- «Mouse embryonic stem cells, but not somatic cells, predominantly use homologous recombination to repair double-strand DNA breaks». Stem Cells Dev. 19 (11): 1699-1711. Noviembre de 2010. PMC 3128311. PMID 20446816. doi:10.1089/scd.2010.0058.
- «Restoration of an absent G1 arrest and protection from apoptosis in embryonic stem cells after ionizing radiation». Proc. Natl. Acad. Sci. U.S.A. 101 (40): 14443-14448. Octubre de 2004. Bibcode:2004PNAS..10114443H. PMC 521944. PMID 15452351. doi:10.1073/pnas.0401346101.
- «ES cells do not activate p53-dependent stress responses and undergo p53-independent apoptosis in response to DNA damage». Curr. Biol. 8 (3): 145-155. Enero de 1998. PMID 9443911. S2CID 13938854. doi:10.1016/S0960-9822(98)70061-2.
- «DNA repair/pro-apoptotic dual-role proteins in five major DNA repair pathways: fail-safe protection against carcinogenesis». Mutat. Res. 511 (2): 145-178. Junio de 2002. PMID 12052432. doi:10.1016/S1383-5742(02)00009-1.
- «Embryonic stem cells and somatic cells differ in mutation frequency and type». Proc. Natl. Acad. Sci. U.S.A. 99 (6): 3586-3590. Marzo de 2002. Bibcode:2002PNAS...99.3586C. PMC 122567. PMID 11891338. doi:10.1073/pnas.062527199.
- «FDA approves human embryonic stem cell study – CNN.com». 23 de enero de 2009.
- «Human embryonic stem cell-derived oligodendrocyte progenitor cell transplants remyelinate and restore locomotion after spinal cord injury». J. Neurosci. 25 (19): 4694-4705. 2005. PMC 6724772. PMID 15888645. doi:10.1523/JNEUROSCI.0311-05.2005.
- Reinberg, Steven (23 de enero de 2009) FDA OKs 1st Embryonic Stem Cell Trial. The Washington Post
- Geron comments on FDA hold on spinal cord injury trial. geron.com (27 de agosto de 2009)
- Vergano, Dan (11 de octubre de 2010). «Embryonic stem cells used on patient for first time». USA Today. Consultado el 12 de octubre de 2010.
- Brown, Eryn (15 de noviembre de 2011). «Geron exits stem cell research». LA Times. Consultado el 15 de noviembre de 2011.
- «Great news: BioTime Subsidiary Asterias Acquires Geron Embryonic Stem Cell Program». iPScell.com. 1 de octubre d 2013.
- Knoepfler, Paul S. (2009). «Deconstructing Stem Cell Tumorigenicity: A Roadmap to Safe Regenerative Medicine». Stem Cells 27 (5): 1050-1056. PMC 2733374. PMID 19415771. doi:10.1002/stem.37.
- Varlakhanova, Natalia V.; Cotterman, Rebecca F.; Devries, Wilhelmine N.; Morgan, Judy; Donahue, Leah Rae; Murray, Stephen; Knowles, Barbara B.; Knoepfler, Paul S. (2010). «Myc maintains embryonic stem cell pluripotency and self-renewal». Differentiation 80 (1): 9-19. PMC 2916696. PMID 20537458. doi:10.1016/j.diff.2010.05.001.
- Wernig, Marius; Meissner, Alexander; Cassady, John P; Jaenisch, Rudolf (2008). «C-Myc is Dispensable for Direct Reprogramming of Mouse Fibroblasts». Cell Stem Cell 2 (1): 10-12. PMID 18371415. doi:10.1016/j.stem.2007.12.001.
- King, Nancy; Perrin, Jacob (7 de julio de 2014). «Ethical issues in stem cell research and therapy». Stem Cell Research & Therapy 5 (4): 85. PMC 4097842. PMID 25157428. doi:10.1186/scrt474.
- «Multipotentiality of Single Embryoncal Carcinoma Cells». Cancer Res 24: 1544-1551. 1964. PMID 14234000.
- Martin GR (1980). «Teratocarcinomas and mammalian embryogenesis». Science 209 (4458): 768-776. Bibcode:1980Sci...209..768M. PMID 6250214. doi:10.1126/science.6250214.
- «Establishment in culture of pluripotent cells from mouse embryos». Nature 292 (5819): 154-156. 1981. Bibcode:1981Natur.292..154E. PMID 7242681. S2CID 4256553. doi:10.1038/292154a0.
- Martin G (1981). «Isolation of a pluripotent cell line from early mouse embryos cultured in medium conditioned by teratocarcinoma stem cells». Proc Natl Acad Sci USA 78 (12): 7634-7638. Bibcode:1981PNAS...78.7634M. PMC 349323. PMID 6950406. doi:10.1073/pnas.78.12.7634.
- «The 2007 Nobel Prize in Physiology or Medicine – Advanced Information». Nobel Prize. Nobel Media.
- «The Life of Dolly | Dolly the Sheep» (en inglés estadounidense). Consultado el 21 de febrero de 2022.
- Klotzko, Arlene Judith; Klotzko, Visiting Scholar Royal Free and University College Medical School Arlene Judith (2006). A Clone of Your Own? (en inglés). Cambridge University Press. ISBN 978-0-521-85294-4.
- Thompson, James A.; Itskovitz-Eldor, Joseph; Shapiro, Sander S.; Waknitz, Michelle A.; Swiergiel, Jennifer J.; Marshall, Vivienne S.; Jones, Jeffrey M. (6 de noviembre de 1998). «Embryonic Stem Cell Lines Derived From Human Blastocyst». Science 282 (5391): 1145-1147. Bibcode:1998Sci...282.1145T. PMID 9804556. doi:10.1126/science.282.5391.1145.
- «President George W. Bush's address on stem cell research». CNN Inside Politics. 9 de agosto de 2001.
- Yamanaka, Shinya; Takahashi, Kazutoshi (25 de agosto de 2006). «Induction of Pluripotent Stem Cells From Mouse Embryonic and Adult Fibroblast Cultures by Defined Factors». Cell 126 (4): 663-676. PMID 16904174. doi:10.1016/j.cell.2006.07.024.
- Wadman, Meredith (27 de enero d 2009). «Stem cells ready for primetime». Nature 457 (7229): 516. PMID 19177087. doi:10.1038/457516a.
- «Executive Order 13505—Removing Barriers To Responsible Scientific Research Involving Human Stem Cells». Federal Register: Presidential Documents 74 (46). 11 de marzo de 2009.
- Mountford, JC (2008). «Human embryonic stem cells: origins, characteristics and potential for regenerative therapy». Transfus Med 18 (1): 1-12. PMID 18279188. S2CID 20874633. doi:10.1111/j.1365-3148.2007.00807.x.
- «Embryonic stem cell lines derived from human blastocysts». Science 282 (5391): 1145-1147. 1998. Bibcode:1998Sci...282.1145T. PMID 9804556. doi:10.1126/science.282.5391.1145.
- «Inhibition of pluripotential embryonic stem cell differentiation by purified polypeptides». Nature 336 (6200): 688-690. 1988. Bibcode:1988Natur.336..688S. PMID 3143917. doi:10.1038/336688a0.
- «Myeloid leukaemia inhibitory factor maintains the developmental potential of embryonic stem cells». Nature 336 (6200): 684-687. 1988. Bibcode:1988Natur.336..684W. PMID 3143916. doi:10.1038/336684a0.
- «Establishment of a germ-line competent C57BL/6 embryonic stem cell line». Exp Cell Res 197 (2): 254-258. 1991. PMID 1959560. doi:10.1016/0014-4827(91)90430-3.
- «Induction of pluripotent stem cells from adult human fibroblasts by defined factors». Cell 131 (5): 861-872. 2007. PMID 18035408. doi:10.1016/j.cell.2007.11.019.
- «Human embryonic stem cell lines derived from single blastomeres». Nature 444 (7118): 481-485. 2006. Bibcode:2006Natur.444..481K. PMID 16929302. S2CID 84792371. doi:10.1038/nature05142.
- US scientists relieved as Obama lifts ban on stem cell research, The Guardian, 10 de marzo de 2009
- Tachibana, Masahito; Amato, Paula; Sparman, Michelle; Gutierrez, Nuria Marti; Tippner-Hedges, Rebecca; Ma, Hong; Kang, Eunju; Fulati, Alimujiang et al. (6 de junio de 2013). «Human Embryonic Stem Cells Derived by Somatic Cell Nuclear Transfer». Cell (en inglés) 153 (6): 1228-1238. ISSN 0092-8674. PMC 3772789. PMID 23683578. doi:10.1016/j.cell.2013.05.006.
- Chung, Young Gie; Eum, Jin Hee; Lee, Jeoung Eun; Shim, Sung Han; Sepilian, Vicken; Hong, Seung Wook; Lee, Yumie; Treff, Nathan R. et al. (5 de junio de 2014). «Human Somatic Cell Nuclear Transfer Using Adult Cells». Cell Stem Cell (en inglés) 14 (6): 777-780. ISSN 1934-5909. PMID 24746675. doi:10.1016/j.stem.2014.03.015.
- Takahashi, Kazutoshi; Yamanaka, Shinya (25 de agosto de 2006). «Induction of Pluripotent Stem Cells from Mouse Embryonic and Adult Fibroblast Cultures by Defined Factors». Cell (en inglés) 126 (4): 663-676. ISSN 0092-8674. PMID 16904174. S2CID 1565219. doi:10.1016/j.cell.2006.07.024.
- «The Nobel Prize in Physiology or Medicine – 2012 Press Release». Nobel Media AB. 8 de octubre de 2012.
- Wernig, Marius; Meissner, Alexander; Foreman, Ruth; Brambrink, Tobias; Ku, Manching; Hochedlinger, Konrad; Bernstein, Bradley E.; Jaenisch, Rudolf (19 de julio de 2007). «In vitro reprogramming of fibroblasts into a pluripotent ES-cell-like state». Nature 448 (7151): 318-324. Bibcode:2007Natur.448..318W. ISSN 1476-4687. PMID 17554336. S2CID 4377572. doi:10.1038/nature05944.
- Takahashi, Kazutoshi; Tanabe, Koji; Ohnuki, Mari; Narita, Megumi; Ichisaka, Tomoko; Tomoda, Kiichiro; Yamanaka, Shinya (30 de noviembre de 2007). «Induction of Pluripotent Stem Cells from Adult Human Fibroblasts by Defined Factors». Cell (en inglés) 131 (5): 861-872. ISSN 0092-8674. PMID 18035408. doi:10.1016/j.cell.2007.11.019.
- Weiss, Rick (7 de diciembre de 2007). «Scientists Cure Mice Of Sickle Cell Using Stem Cell Technique: New Approach Is From Skin, Not Embryos». The Washington Post. pp. A02.
- Hanna, J.; Wernig, M.; Markoulaki, S.; Sun, C.-W.; Meissner, A.; Cassady, J. P.; Beard, C.; Brambrink, T. et al. (2007). «Treatment of Sickle Cell Anemia Mouse Model with iPS Cells Generated from Autologous Skin». Science 318 (5858): 1920-1923. Bibcode:2007Sci...318.1920H. PMID 18063756. doi:10.1126/science.1152092.
- Helen Briggs (17 de enero de 2008). «US team makes embryo clone of men». BBC. pp. A01.
- Ebert, Jessica (24 de enero de 2005). «Human stem cells trigger immune attack». Nature News (London: Nature Publishing Group). doi:10.1038/news050124-1. Archivado desde el original el 24 de septiembre de 2010. Consultado el 27 de febrero de 2009.
- «Human embryonic stem cells express an immunogenic nonhuman sialic acid». Nat. Med. 11 (2): 228-232. 2005. PMID 15685172. S2CID 13739919. doi:10.1038/nm1181.
- «Human embryonic stem cells derived without feeder cells». Lancet 365 (9471): 1636-1641. 2005. PMID 15885296. doi:10.1016/S0140-6736(05)66473-2.
- Kuroda, Y.; Kitada, M.; Wakao, S.; Nishikawa, K.; Tanimura, Y.; Makinoshima, H.; Goda, M.; Akashi, H. et al. (2010). «Unique multipotent cells in adult human mesenchymal cell populations». Proceedings of the National Academy of Sciences 107 (19): 8639-8643. Bibcode:2010PNAS..107.8639K. PMC 2889306. PMID 20421459. doi:10.1073/pnas.0911647107.
- Muse Cells | SpringerLink.
- Zikuan Leng 1 2, Dongming Sun 2, Zihao Huang 3, Iman Tadmori 2, Ning Chiang 2, Nikhit Kethidi 2, Ahmed Sabra 2, Yoshihiro Kushida 4, Yu-Show Fu 3, Mari Dezawa 4, Xijing He 1, Wise Young 2Quantitative Analysis of SSEA3+ Cells from Human Umbilical Cord after Magnetic SortingCell Transplant. 2019 Jul;28(7):907–923.
- Wakao, S.; Kitada, M.; Kuroda, Y.; Shigemoto, T.; Matsuse, D.; Akashi, H.; Tanimura, Y.; Tsuchiyama, K. et al. (2011). «Multilineage-differentiating stress-enduring (Muse) cells are a primary source of induced pluripotent stem cells in human fibroblasts». Proceedings of the National Academy of Sciences 108 (24): 9875-9880. Bibcode:2011PNAS..108.9875W. PMC 3116385. PMID 21628574. doi:10.1073/pnas.1100816108.
- Dezawa, Mari (2016). «Muse Cells Provide the Pluripotency of Mesenchymal Stem Cells: Direct Contribution of Muse Cells to Tissue Regeneration». Cell Transplantation 25 (5): 849-861. PMID 26884346. doi:10.3727/096368916X690881.
- Hori, Emiko; Hayakawa, Yumiko; Hayashi, Tomohide; Hori, Satoshi; Okamoto, Soushi; Shibata, Takashi; Kubo, Michiya; Horie, Yukio et al. (2016). «Mobilization of Pluripotent Multilineage-Differentiating Stress-Enduring Cells in Ischemic Stroke». Journal of Stroke and Cerebrovascular Diseases 25 (6): 1473-1481. PMID 27019988. doi:10.1016/j.jstrokecerebrovasdis.2015.12.033.
- Kuroda, Yasumasa; Wakao, Shohei; Kitada, Masaaki; Murakami, Toru; Nojima, Makoto; Dezawa, Mari (2013). «Isolation, culture and evaluation of multilineage-differentiating stress-enduring (Muse) cells». Nature Protocols 8 (7): 1391-1415. PMID 23787896. S2CID 28597290. doi:10.1038/nprot.2013.076.
- Ogura, Fumitaka; Wakao, Shohei; Kuroda, Yasumasa; Tsuchiyama, Kenichiro; Bagheri, Mozhdeh; Heneidi, Saleh; Chazenbalk, Gregorio; Aiba, Setsuya et al. (2014). «Human Adipose Tissue Possesses a Unique Population of Pluripotent Stem Cells with Nontumorigenic and Low Telomerase Activities: Potential Implications in Regenerative Medicine». Stem Cells and Development 23 (7): 717-728. PMID 24256547. doi:10.1089/scd.2013.0473.
- Heneidi, Saleh; Simerman, Ariel A.; Keller, Erica; Singh, Prapti; Li, Xinmin; Dumesic, Daniel A.; Chazenbalk, Gregorio (2013). «Awakened by Cellular Stress: Isolation and Characterization of a Novel Population of Pluripotent Stem Cells Derived from Human Adipose Tissue». PLOS ONE 8 (6): e64752. Bibcode:2013PLoSO...864752H. PMC 3673968. PMID 23755141. doi:10.1371/journal.pone.0064752.
Enlaces externos
- Comprender las células madre: una visión de la ciencia y los problemas de las academias nacionales Archivado el 9 de abril de 2010 en Wayback Machine.
- Institutos Nacionales de Salud
- Taller práctico de la Universidad de Oxford sobre tecnología de células madre pluripotentes. Archivado el 8 de abril de 2016 en Wayback Machine. Archivado.
- Hoja informativa sobre células madre embrionarias
- Hoja informativa sobre cuestiones éticas en la investigación con células madre embrionarias
- Información y alternativas a la investigación con células madre embrionarias
- Un blog centrado específicamente en las células ES y las células iPS, incluidos temas de investigación, biotecnología y orientados al paciente.
- Esta obra contiene una traducción derivada de «Embryonic stem cell» de Wikipedia en inglés, concretamente de esta versión, publicada por sus editores bajo la Licencia de documentación libre de GNU y la Licencia Creative Commons Atribución-CompartirIgual 4.0 Internacional.
| Control de autoridades |
|
|---|
 Datos: Q1151519
Datos: Q1151519 Multimedia: Embryonic stem cells / Q1151519
Multimedia: Embryonic stem cells / Q1151519